
Афферентные и эфферентные связи базальных ганглиев
К базальным ганглиям относятся следующие анатомические образования: полосатое тело (стриатум), состоящее из хвостатого ядра и скорлупы; бледный шар (паллидум), подразделяющийся на внутренний и внешний отделы; черная субстанция и субталамическое ядро Льюиса.
Функции БГ:
1. Центры сложных безусловных рефлексов и инстинктов
2. Участие в формировании условных рефлексов
3. Координация тонуса мышц и произвольных движений. Контроль амплитуды, силы, направления движений
4. Координация сочетанных двигательных актов
5. Контроль за движением глаз (саккады).
6. Программирование сложных целенаправленных движений
7. Центры торможения агрессивных реакций
8. Высшие психические функции (мотивации, прогнозирование, познавательная деятельность). Сложные формы восприятия внешней информации (например, осмысление текста)
9. Участие в механизмах сна
Афферентные связи базальных ганглиев. Большая часть афферентных сигналов, приходящих к базальным ганглиям поступает в полосатое тело. Эти сигналы исходят почти исключительно из трех источников:
- от всех областей коры больших полушарий;
- от внутрипластинчатых ядер таламуса;
- от черной субстанции ( по дофаминэргическому пути).
Эфферентные волокна от стриатума идут к бледному шару и черной субстанции. От последней начинается не только дофаминэргический путь к полосатому телу, но и пути, идущие к таламусу.
От внутреннего отдела бледного шара берет начало самый важный из всех эфферентных трактов базальных ганглиев, заканчивающийся в таламусе, а так же в крыше среднего мозга. Посредством стволовых образований, с которыми связаны базальные ганглии, центробежные импульсы следуют к сегментарным двигательным аппаратам и мускулатуре по нисходящим проводникам.
- от красных ядер - по руброспинальному тракту;
- от ядра Даркшевича – по заднему продольному пучку к ядрам 3, 4,6 нервов и через его посредство к ядру вестибулярного нерва;
- от ядра вестибулярного нерва – по вестибулоспинальному тракту;
- от четверохолмия - по тектоспинальному тракту;
- от ретикулярной формации - по ретикулоспинальному тракту.
Таким образом, базальные ганглии играют, главным образом, роль промежуточного звена в цепи, связываемой двигательные области коры со всеми остальными ее областями.
В раннем филогенезе, когда кора головного мозга еще не была развита, стриопаллидарная система являлась главным двигательным центром, определяющим поведение животного. Чувствительные импульсы, притекающие из зрительного бугра, перерабатывались здесь в двигательные, направляющиеся к сегментарному аппарату и мускулатуре. За счет стрио-паллидарных аппаратов осуществлялись диффузные движения тела достаточно сложного характера: передвижения, плавание и др.
При дальнейшей эволюции нервной системы ведущая роль в движениях переходит к коре головного мозга с ее двигательным анализатором и пирамидной системой. Наконец, у человека возникают сложнейшие действия, носящие целенаправленный, произвольный характер с тонкой дифференцировкой отдельных движений.
Становление функции базальных ганглиев в онтогенезе. Базальные ганглии развиваются интенсивнее, чем зрительные бугры. Бледное ядро миелинизируется раньше, чем полосатое тело и кора головного мозга. Установлено, что миелинизация в бледном шаре почти полностью заканчивается к 8 месяцам развития плода. В полосатом теле миелинизация начинается у плода, а заканчивается только к 2 месяцам жизни. Хвостатое тело в течение первых 2 лет жизни увеличивается в 2 раза, что связывают с развитием у ребенка автоматических двигательных актов.
Двигательная активность новорожденного в значительной мере связана с бледным ядром, импульсы от которого вызывают некоординированные движения головы, туловища и конечностей.
У новорожденного паллидум уже имеет связи со зрительным бугром, подбугровой областью и черной субстанцией. Связь паллидума со стриатутом развивается позже, часть стриопаллидарных волокон оказывается миелинизированная на первом месяце жизни, а другая часть - лишь к 6 месяцам и позже.
Считают, что такие акты, как плач, в моторном отношении осуществляются за счет одного паллидума. С развитием полосатого тела связано появление мимических движений, а затем умение сидеть и стоять. Так как стриатум оказывают тормозное влияние на паллидум, то создается постепенное разделение движений. Для того чтобы сидеть, ребенок должен уметь вертикально держать голову и спину. Это появляется у него к двум месяцам. Сидеть начинает к 6-8 месяцам.
В первые месяцы жизни у ребенка имеется отрицательная реакция опоры: при попытке поставить его на ножки он поднимает их и подтягивает к животу. Затем эта реакция становится положительной: при прикосновении к опоре ножки разгибаются. В 9 месяцев ребенок может стоять с помощью поддержки, в 10 месяцев он стоит свободно.
С 4-5 месячного возраста довольно быстро развиваются произвольные движения, но они еще длительное время сопровождаются многообразными дополнительными движениями.
Появление произвольных (таких как схватывание) и выразительных движений (улыбка, смех) связывают с развитием стриатной системы и двигательных центров коры больших полушарий. Громко смеяться ребенок начинает с 8 месяцев.
По мере роста и развития всех отделов головного мозга и коры больших полушарий движение ребенка становится менее обобщенными и более координированными. Только к концу дошкольного периода устанавливается определенное равновесие коркового и подкоркового двигательных механизмов.
К базальным ганглиям относятся следующие анатомические образования:
- Центры сложных безусловных рефлексов и инстинктов
- Участие в формировании условных рефлексов
- Координация тонуса мышц и произвольных движений. Контроль амплитуды, силы, направления движений
- Координация сочетанных двигательных актов
- Контроль за движением глаз (саккады).
- Программирование сложных целенаправленных движений
- Центры торможения агрессивных реакций
- Высшие психические функции (мотивации, прогнозирование, познавательная деятельность). Сложные формы восприятия внешней информации (например, осмысление текста)
- Участие в механизмах сна
Афферентные связи базальных ганглиев.
Большая часть афферентных сигналов, приходящих к базальным ганглиям поступает в полосатое тело. Эти сигналы исходят почти исключительно из трех источников:
— от всех областей коры больших полушарий;
— от внутрипластинчатых ядер таламуса;
— от черной субстанции ( по дофаминэргическому пути).
Эфферентные волокна от стриатума идут к бледному шару и черной субстанции. От последней начинается не только дофаминэргический путь к полосатому телу, но и пути, идущие к таламусу.
От внутреннего отдела бледного шара берет начало самый важный из всех эфферентных трактов базальных ганглиев, заканчивающийся в таламусе, а так же в крыше среднего мозга. Посредством стволовых образований, с которыми связаны базальные ганглии, центробежные импульсы следуют к сегментарным двигательным аппаратам и мускулатуре по нисходящим проводникам.
— от красных ядер — по руброспинальному тракту;
— от ядра Даркшевича – по заднему продольному пучку к ядрам 3, 4,6 нервов и через его посредство к ядру вестибулярного нерва;
— от ядра вестибулярного нерва – по вестибулоспинальному тракту;
— от четверохолмия — по тектоспинальному тракту;
— от ретикулярной формации — по ретикулоспинальному тракту.
Таким образом, базальные ганглии играют, главным образом, роль промежуточного звена в цепи, связываемой двигательные области коры со всеми остальными ее областями.
Повреждение базальных ганглиев сопровождается самыми различными нарушениями движений. Из всех этих нарушений наиболее известен синдром Паркинсона.
Походка — осторожная, мелкими шажками, замедленная, напоминает старческую походку. Нарушена инициация движения: двинутся вперед удается не сразу. Но в дальнейшем больной не может сразу остановиться: его все еще продолжает тянуть вперед.
Мимика – крайне бедна, лицо принимает застывшее маскообразное выражение. Улыбка, гримаса плача при эмоциях с запозданием возникают и так же медленно исчезают.
Обычная поза — спина согнута, голова наклонена к груди, руки согнуты в локтевых, в лучезапястных, ноги – в коленных суставах (поза просителя).
Речь — тихая, монотонная, глухая, без достаточных модуляций и звучности.
Акинезия — (гипокинезия) – большие трудности в проявлении и двигательной инициации: затруднение при начале и завершения движения.
Тремор покоя — дрожание, которое наблюдается в покое, выражено в дистальных отделах конечностей, иногда в нижней челюсти и отличается малой амплитудой, частотой и ритмичностью. Тремор исчезает во время целенаправленных движений и возобновляется после их окончания (отличие от мозжечкового тремора, появляющегося при движении и исчезающего в покое).
Синдром Паркинсона связан с разрушением пути (тормозного), идущего от черной субстанции к полосатому телу. В области полосатого тела из волокон этого пути выделяется медиатор дофамин. Проявление паркинсонизма и, в частности, акинезия успешно лечатся введением предшественника дофамина — дофа. Наоборот, разрушение областей бледного шара и таламуса (вентролатерального ядра), при котором прерываются пути к двигательной коре, приводит к подавлению непроизвольных движений, но не снимает акинезии.
При поражении хвостатого ядра развивается атетоз — в дистальных отделах конечностей наблюдаются медленные, червеобразные, извивающиеся движения с некоторыми интервалами, во время которых конечность принимает неестественные положения. Атетоз может быть ограниченным и распространенным.
При поражении скорлупы развивается хорея — отличается от атетоза быстротой подергиваний и наблюдаются в проксимальных отделах конечностей и на лице. Характерна быстрая сменяемость локализации судорог, то подергиваются мимические мышцы, то мускулатура ноги, одновременно глазные мышцы и рука и т. д. В выраженных случаях больной становится похожим на паяца. Часто наблюдается гримасничанье, причмокивание, расстраивается речь. Движения становятся размашистыми, избыточными, походка танцующей.
Базальные ганглии (стриопаллидарная система)– это совокупность трех парных образований, расположенных в конечном мозге на основании больших полушарий. Включает в себя 3 парных образования (рис.17):

Рис.17. Анатомия базальных ядер.
- бледный шар – состоит из наружного и внутреннего сегмента, является наиболее филогенетически древней частью;
- полосатое тело – состоит из хвостатого ядра и скорлупы;
- ограда – расположена между скорлупой и островковой корой, является наиболее молодой частью.
Установлены внутренние связи между базальными ганглиями. За их счет компоненты базальных ганглиев тесно взаимодействуют и образуют единую стриопаллидарную систему. Кроме того, базальные ганглии имеют многочисленные афферентные и эфферентные связи с другими структурами мозга.
Афферентные связи поступают (рис. 17) в базальные ядра от всех областей коры прямо и через таламус, от неспецифических ядер таламуса, от черного вещества.

Рис. 17. Схема основных афферентных и эфферентных связей базальных ядер:
ЧВ – черное вещество; ВЯ – вентральные ядра; ИЛЯ – интраламинарное ядро;
Имеются три эфферентных выхода:
1. от полосатого тела тормозящие пути идут к бледному шару, от бледного шара к двигательным ядрам таламуса и от них к двигательной коре (поля 4 и 6);
2. часть эфферентных волокон из бледного шара и полосатого тела идет к ретикулярной формации и красному ядру ствола мозга и далее в спинной мозг, а также через нижнюю оливу в мозжечок;
3. от полосатого тела тормозящие пути идут также к черному веществу и далее к ядрам таламуса.
В последнее время было выявлено, что связи базальных ганглий образуют параллельные функциональные петли (рис.18): скелетно-моторную и глазодвигательную петли.
Скелетно-моторная петля служит для регуляции таких параметров движения как амплитуда, сила и направление. Она соединяет премоторную, двигательную и соматосенсорную области коры со скорлупой базальных ядер, далее импульсация идет в бледный шар и черное вещество среднего мозга и через двигательное ядро (вентролатеральное ядро) таламуса возвращается в премоторную кору.
Глазодвигательная петля участвует в регуляции движений глаз (например, скачкообразных движений глаз – саккад). Она соединяет области коры, контролирующие направление взгляда (поле 8 лобной коры и поле 7 теменной коры), с хвостатым ядром. Оттуда импульсы поступают в бледный шар и черное вещество из которых она поступает в ассоциативное медиадорсальное и переднее релейное вентральное ядро таламуса и возвращается в лобное глазодвигательное поле 8.
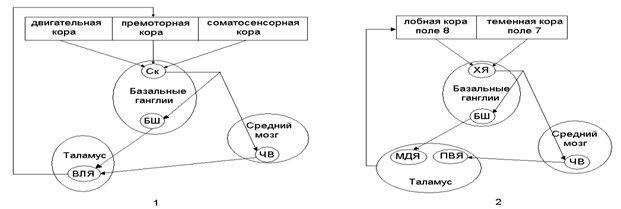
Рис.18. Схема функциональных петель, проходящих через
базальные ганглии (по Р.Шмидту, 1996, с изменениями)
1 – скелетно-моторная петля; 2 – глазодвигательная петля;
Ск – скорлупа; БШ – бледный шар; ЧВ – черное вещество;
ВЛЯ – вентролатеральное ядро; ХЯ - хвостатое ядро; МДЯ – медиодорсальное ядро; ПВЯ – переднее вентральное ядро
Функции базальных ядер
Основными функциями базальных ядер являются:
1. Регуляция двигательной активности. Важную роль базальные ядра играют при переходе от замысла движения (фазы подготовки к движению) к выбранной программе действия (фазе выполнения).
2. Участие в интеграции (объединении) тонических рефлексов и в обеспечении позы.
3. Участие, наряду с мозжечком, в выработке сложных двигательных программ.
4. Участие в контроле таких параметров движения как сила, амплитуда, скорость и направление движения.
Функции полосатого тела.Полосатое тело состоит из более крупных нейронов с длинными отростками, которые выходят за пределы стриапаллидарной системы. Полосатое тело участвует в регуляции мышечного тонуса, уменьшая его, а также в организации и регуляции движений. Стимуляция полосатого тела вызывает:
- простые двигательные реакции;
- торможение активности коры, подкорковых образований, безусловных и условных рефлексов;
- торможение поведенческих реакций (пищедобывательной и ориентировочной).
Полосатое тело оказывает на бледный шар тормозящее влияние (медиатор ГАМК), снижая активность бледного шара, и тем самым, снижая двигательную активность.
Полосатое тело имеет двухсторонние связи с черным веществом среднего мозга. Нейроны полосатого тела оказывают тормозное влияние (медиатор ГАМК) на нейроны черного вещества. Нейроны черного вещества секретируют медиатор дофамин, который посредством аксонного транспорта поступает к базальным ганглиям. При резком снижении концентрации дофамина в хвостатом ядре появляются симптомы болезни Паркинсона: сильное дрожание пальцев, губ, головы и других частей тела, ригидность мышц и т.д.
На двигательную кору полосатое тело также оказывает тормозное влияние, сдерживая ненужную в данный момент степень свободы движения, и тем самым обеспечивая четкость и направленность двигательных реакций.
Поражение полосатого тела сопровождается возникновением так называемых гиперкинезов. Гиперкинезы – это избыточные, насильственные, непроизвольные движения. Движения у больных становятся размашистыми, избыточным, широко амплитудными. Больные не могут усидеть на одном месте, все время вертятся, меняют положение тела и т.д. Кроме того, могут наблюдаться следующие симптомы:
· Тремор – подергивание конечностей;
· Ригидность мышц – повышение мышечного тонуса;
· Баллизм – крупноразмашистый гиперкинез конечностей;
· Атетоз – больше наблюдается в дистальных отделах конечностей – в кистях и пальцах рук; проявляется медленными, извивающимися, червеобразными движениями;
Скорлупа отвечает за организацию пищевого поведения. При раздражении скорлупы появляется слюноотделение и изменение дыхания, при поражении наблюдаются трофические изменения в коже.
Функции бледного шара. Бледный шар оказывает модулирующее влияние на двигательную кору, мозжечок, ретикулярную формацию и красное ядро. Его функцией является провоцирование ориентировочной реакции, движения конечностей, активация пищевого поведения (жевание, глотание).
Ограда – функция мало изучена. Имеет двухсторонние связи с лобной, затылочной, височной корой, обонятельной луковицей, таламусом и другими базальными ядрами. При раздражении ограды вызываются моторные реакции со стороны пищеварительного тракта (жевание, глотание, рвотные движения), ориентировочные реакции. При повреждении – теряется способность говорить.

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.
Большая часть афферентных сигналов, приходящих к базальным ганглиям, поступает в полосатое тело базальных ганглий. Эти сигналы исходят почти исключительно из трех источников: от всех областей коры больших полушарий, от внутрипластинчатых ядер таламуса и черной субстанции базальных ганглий. Эфферентные волокна от стриатума идут к бледному шару базальных ганглий и черной субстанции, от которой начинается дофаминергический путь к полосатому телу и пути, идущие к таламусу. От внутреннего отдела бледного шара берет начало самый важный из всех эфферентных трактов, заканчивающийся в таламусе и в крыше среднего мозга.
Гипоталамус: зоны
В поперечном направлении гипоталамус можно разделить на три зоны - перивентрикулярную, медиальную и латеральную.
Перивентрикулярная зона гипоталамуса представляет собой тонкую полоску, прилежащую к третьему желудочку.
В медиальной зоне гипоталамуса различают несколько дискретных участков, называемых ядрами и представляющих собой скопления тел нейронов: преоптическое ядро, паравентрикулярное ядро, супраоптическое ядро, переднее ядро, инфундибулярное ядро, вентромедиальное ядро, дорсомедиальное ядро и заднее ядро. От медиальной области гипоталамуса начинается ножка гипофиза, соединяющаяся с аденогипофизом и нейрогипофизом. Передняя часть этой ножки называется срединным возвышением гипофиза. Здесь оканчиваются отростки многих нейронов преоптической области гипоталамуса и передней области гипоталамуса, а также инфундибулярного и вентромедиального ядер. Здесь из этих отростков высвобождаются гормоны, поступающие через систему сосудов к передней доле гипофиза. Совокупность ядерных зон, содержащих гормонопродуцирующие нейроны, называется гипофизотропной областью. Отростки нейронов супраоптического и паравентрикулярного ядер идут к задней доле гипофиза, они регулируют образование и высвобождение окситоцина и вазопрессина. Связать конкретные функции гипоталамуса с его остальными ядрами невозможно.
В латеральной зоне гипоталамуса не существует ядер. Нейроны этой зоны располагаются диффузно вокруг медиального пучка переднего мозга, идущего от основания лимбической системы к передним центрам промежуточного мозга.
Гипоталамус: афферентные и эфферентные связи
Латеральный гипоталамус образует двусторонние связи с верхними отделами ствола мозга, центральным серым веществом среднего мозга и с лимбической системой. Сигналы от поверхности тела и внутренних органов поступают в гипоталамус по восходящим путям через таламус или через лимбическую область среднего мозга. Эфферентные связи гипоталамуса с вегетативными и соматическими ядрами ствола мозга и спинного мозга образованы полисинаптическими путями, идущими в составе ретикулярной формации. Медиальный гипоталамус обладает двусторонними связями с латеральным, и, кроме того, непосредственно получает сигналы от некоторых отделов головного мозга. В медиальной области гипоталамуса существуют нейроны, воспринимающие важнейшие параметры крови и спинномозговой жидкости, т.е. следят за состоянием внутренней среды организма. Они могут воспринимать температуру крови, водно-электролитный состав плазмы и содержание гормонов в крови. Через нервные механизмы медиальная область гипоталамуса управляет деятельностью нейрогипофиза, а через гормональные - аденогипофиза. Таким образом, эта область служит промежуточным звеном между нервной системой и эндокринной системой.
Гипоксемия - пониженное содержание кислорода в крови, ведущее к гипоксии.
Гипоксия - кислородное голодание.
Гипоталамус - отдел промежуточного мозга (под таламусом), в котором расположены центры вегетативной нервной системы; тесно связан с гипофизом. Нервные клетки гипоталамуса вырабатывают гормоны вазопрессин и окситоцин, а также рилизинг-факторы, стимулирующие или угнетающие секрецию гормонов гипофизом. Гипоталамус регулирует обмен веществ, деятельность пищеварительной, сердечно-сосудистой, выделительной систем и желез внутренней секреции, механизмы сна, бодрствования, эмоции. Осуществляет связь нервной и эндокринной систем.
Базальные ганглии (также базальные ядра, лат. nuclei basales) — комплекс подкорковых нейронных узлов, расположенных в центральном белом веществе полушарий большого мозга. Базальные ганглии входят в состав переднего мозга, расположенного на границе между лобными долями и над стволом мозга и включают в себя следующие компоненты:
Функция базальных ганглиев.
Базальные ганглии обеспечивают регуляцию двигательных и вегетативных функций, участвуют в осуществлении интегративных процессов высшей нервной деятельности.
Нарушения в базальных ядрах приводит к моторным дисфункциям, таким как, замедленность движения, изменения мышечного тонуса, непроизвольные движения, тремор. Эти нарушения фиксируются при болезни Паркинсона и болезни Хантингтона.
В основании больших полушарий (нижняя стенка боковых желудочков) расположены ядра серого вещества — базальные ганглии. Они составляют примерно 3% от объема полушарий. Всебазальные ганглии функционально объединены в две системы. Первая группа ядер представляет собой стриопаллидарную систему. К ним относятся: хвостатое ядро (nucleus caudatus), скорлупа (putamen) и бледный шар (globus pallidus). Скорлупа и хвостатое ядро имеют слоистую структуру, и поэтому их общее название — полосатое тело (corpus striatum). Бледный шар не имеет слоистости и выглядит светлее стриатума. Скорлупа и бледный шар объединены в
чечевицеобразное ядро (nucleus lentiförmis). Скорлупа образует наружный слой чечевицеобразного
ядра, а бледный шар — внутренние его части. Бледный шар, в свою очередь, состоит из наружного
и внутреннего члеников.
Анатомически хвостатое ядро тесно связано с боковым желудочком. Расположенная впереди и
медиально расширенная его часть — головка хвостатого ядра образует боковую стенку переднего
рога желудочка, тело ядра — нижнюю стенку центральной части желудочка, а тонкий хвост —
верхнюю стенку нижнего рога. Следуя форме бокового желудочка, хвостатое ядро охватывает
дугой чечевицеобразное ядро. Друг от друга хвостатое и чечевицеобразное ядра
отделены прослойкой белого вещества — частью внутренней капсулы (capsula interna). Другая
часть внутренней капсулы отделяет чечевицеобразное ядро от нижележащего таламуса. Таким образом, строение дна бокового желудочка (представляющего собой стриопаллидарную
систему) схематично можно представить себе так: стенку самого желудочка образует слоистое
хвостатое ядро, затем ниже идет прослойка белого вещества. Стриопаллидарная система получает афферентные волокна от неспецифических медиальных
таламических ядер, лобных отделов коры больших полушарий, коры мозжечка и черной
субстанции среднего мозга. Основная масса эфферентных волокон стриатума радиальными
пучками сходится к бледному шару. Таким образом, бледный шар является выходной структурой
стриопаллидарной системы. Эфферентные волокна бледного шара идут к передним ядрам
таламуса, которые связаны с фронтальной и теменной корой больших полушарий. Часть
эфферентных волокон, не переключающихся в ядре бледного шара, идет к черной субстанции и
красному ядру среднего мозга. Стриопаллидум, совместно со своими проводящими
путями, входит в экстрапирамидную систему, оказывающую тоническое влияние на моторную
деятельность. Эта система контроля над движениями называется экстрапирамидной потому, что
переключается на пути к спинному мозгу, минуя пирамиды продолговатого мозга.
Стриопаллидарная система является высшим центром непроизвольных и автоматизированных
движений, снижает мышечный тонус, тормозит движения, осуществляемые двигательной корой.
Латеральнее стриопаллидарной системы базальных ганглиев расположена тонкая пластинка
серого вещества — ограда (claustrum). Она ограничена со всех сторон волокнами белого вещества
— наружной капсулой (capsula externa).
Остальные базальные ядра входят в лимбическую.
Структура и функции таламических ядер. Специфические и неспецифические ядра таламуса. Реакция вовлечения. Взаимодействие между неспецифическими ядрами таламуса и ретикулярной фармацией. Релейная функция таламических ядер. Роль таламуса в механизмах формирования боли.
Промежуточный мозг (diencephalon) интегрирует сенсорные, двигательные и вегетативные реакции, необходимые для целостной деятельности организма. Основными образованиями промежуточного мозга являются таламус, гипоталамус, который состоит из свода и эпифиза, и таламической области, которая включает в себя таламус, эпиталамус и метаталамус.
Таламус
Таламус (thalamus, зрительный бугор) — структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга.
Морфофункциональная организация. В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприорецепторов и интероцепторов и начинаются таламокортикальные пути.
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом (подтверждением тому служит наличие в таламусе около 120 разнофункциональных ядер). Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная — в лобную долю коры; латеральная — в теменную, височную, затылочную доли коры. По проекциям определяется и функция ядер. Такое деление не абсолютно, так как одна часть волокон от ядер таламуса идет в строго ограниченные корковые образования, другая — в разные области коры большого мозга.
Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
К специфическим ядрам относятся переднее вентральное, медиальное, вентролатеральиое, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Последние относятся к подкорковым центрам зрения и слуха соответственно.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III—IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
В медиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Ассоциативные ядра таламуса представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное — с лобной долей коры, латеральное дорсальное — с теменной, подушка — с ассоциативными зонами теменной и височной долями коры большого мозга.
Основными клеточными структурами этих ядер являются мультиполярные, биполярные трехотростчатые нейроны, т. е. нейроны, способные выполнять полисенсорные функции. Ряд нейронов изменяет активность только при одновременном комплексном раздражении. На полисенсорных нейронах происходит конвергенция возбуждений разных модальностей, формируется интегрированный сигнал, который затем передается в ассоциативную кору мозга. Нейроны подушки связаны главным образом с ассоциативными зонами теменной и височной долей коры большого мозга, нейроны латерального ядра — с теменной, нейроны медиального ядра — с лобной долей коры большого мозга.
Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса.
Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, т. е. развитие сонного состояния.
Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер, позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех. Двигательные реакции интегрируются в таламусе с вегетативными процессами, обеспечивающими эти движения.
Конвергенцию сенсорных стимулов в таламус обусловливает возникновение так называемых таламических неукротимых болей, которые возникают при патологических процессах в самом таламусе.
Читайте также:
