
Бездекрементное проведение возбуждения по нервным волокнам
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000)
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Тип
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.
Лекционный материал.
1. Какие энергии (в кДж/Моль) имеют Ван-дер-Ваальсовы взаимодействия? 0,8 — 8,16 кДж/Моль.
2. Какие виды химической связи известны для атомов углерода? В большинстве соединений углерод имеет валентность IV и образует обычно ковалентные связи. Алмаз имеет твёрдую кристаллическую решётку, а графит слоистое строение за счёт Ван-дер-Ваальсовых взаимодействий.
Какие молекулы имеют нулевой электрический дипольный момент? Неполярные молекулы при отсутствии внешних воздействий.
4. Каким законом описывается взаимодействие точечных зарядов? Закон Кулона.
5. Какие химические элементы определяют состав живой клетки? С, Н, N, О, Р, S.
6. Какие степени окисления присущи углероду в химических связях? -4, 0, +2,+4.
7. Какими терминами характеризуют ионную связь? Ненаправленная, ненасыщенная.
8. Какое вещество имеет наибольшую теплоемкость? Вода.
9. Какова величина (в градусах) угла между О-Н связями в молекуле воды? 108°.
10. Как меняется объем воды при замерзании? Объём воды при замерзании возрастает примерно на 11 %.
11. Свойства воды. 1)Аномально высокая теплоёмкость; 2)Высокая удельная теплота плавления; 3) Универсальный растворитель; 4)Самое высокое поверхностное натяжение среди жидкостей; 5) Дистиллированная вода не проводит электрический ток.
12. Как называются вещества, растворяющиеся друг в друге в любых пропорциях? Полностью взаиморастворимыми.
13. Что называют растворителем? Растворитель - компонент, присутствующий в растворе в большем количестве (вещество, способное растворять в себе другое вещество).
14. Какими размерами частиц характеризуются коллоидные смеси? 10 в –9–10 в –7 м или 10 в -7 до 10 в -5 см или 1-500 (1000) нм.
15. Что называют суспензией? Суспензия — это грубо дисперсная система с твёрдой дисперсной фазой и жидкой дисперсионной средой. Частицы дисперсной фазы > 10 мкм. Суспензии, в которых оседание идёт медленно из-за малой разницы плотности дисперсной фазы и дисперсионной среды, называютвзвесями.
16. Чем определяется диффузия ионов в живой клетке? Диффузия(пассивный транспорт) ионов идёт в направлении их меньшей концентрации. Диффузия идёт без затрат химической энергии.
Что такое термодинамическая гибкость полимерных цепей? Гибкость – способность полимерных цепей изменять конфигурацию. Термодинамическая гибкость осуществляется под действием микроброуновского движения. Гибкость обусловлена возможностью вращенияатомов в цепи и отдельных звеньев вокруг одинарных связей; зависит от внешних факторов.
18. Что такое липиды? Липиды - жироподобные вещества, входящие в состав всех живых клеток и играющие важную роль в жизненных процессах, нерастворимые в воде, но хорошо растворимые в неполярных растворителях (эфире, бензине).
19. Что такое флип-флоп диффузия? Флип-флоп - это диффузия молекул липидного бислоя поперек мембраны (переход молекулы с одной стороны липидной мембраны на другую).
20. Что такое температура фазового перехода? Температура фазового перехода – температура, при которой половина молекул фосфолипидов находится в твердой фазе, половина в жидкой.
21. Из чего складывается полный электростатический потенциал липидного бислоя? 1)поверхностный потенциал; 2)трансмембранный потенциал;3)внутренний потенциал (потенциал внутреннего диполя).
22. Что такое мицелла? Мицеллы — частицы в коллоидных системах, состоят из нерастворимого в данной среде ядра очень малого размера, окруженного стабилизирующей оболочкой адсорбированных ионов и молекул растворителя.
23. Какиесилы способствуют образованию липидных агрегатов в водной среде? 1)гидрофобные взаимодействия; 2)силы Ван-дер-Ваальса; 3)водородные связи.
24. Что такое кинк? Кинк– изгиб молекулы (нарушение регулярного расположения липидных молекул в бислое и появление в нем дефектов упаковки.)
25. При каких температурах возникают кинки? Температура окружающей среды > температуры фазового перехода.
26. Сколько уровней структуры белка существует? Четыре.
27. Какие типы связей стабилизируют первичную структуру белка? Ковалентные связи.
29. Классификация белков по функциям (без повторений). 1)Каталитические (ферменты); 2) Структурные(фибриллярные: коллаген, эластин); 3)Защитные (антитела); 4)Регуляторные (гормоны); 5)Транспортные(Hb, K-Na-АТФаза); 6)Сократительные (актин, миозин); 7) Рецепторные (рецепторы в мембране); 8)Резервные (казеин молока); 9)Специализированные (белки-шапероны).
30. Какую функцию выполняют белки-шапероны? Главная функция - обеспечениевосстановления структурыбелков после повреждения.
31. Что такое высаливание? Высаливание — обратимая реакция осаждения белков из раствора с помощью больших концентраций солей.
32. ?Каким образом можно вычислить толщину билипидного слоя мембраны?
33. Что такое активный транспорт ионов через мембрану? Активный транспорт - это перенос иона через мембрану, протекающий против электрохимического градиента с помощью белка-переносчика с затратой свободнойэнергии организма.
34. Концентрационный мембранный градиент какогоиона играет ключевую роль в формировании потенциала покоя клетки? К+.
35. С чем связано происхождение потенциала действия клетки? Причины возникновения потенциала действия: 1)увеличивается электропроводность клеточной мембраны; 2)изменяется проницаемость мембраны для разных типов ионов.
36. Какие физические величины располагаются в уравнении Гольдмана-Ходжкина-Катца после логарифма? Проницаемостии концентрации ионов Na+, K+, Cl-.
37. Какое явление в клетке описывает уравнение Ходжкина-Хаксли? Процесс возбуждения мембраны за счёт ионных токов(процесс формирования нервного импульса).
38. На что влияет явление рефрактерности в клетке? На изменение проницаемости мембран, сопровождающее возникновением потенциала действия. Рефрактерность -кратковременное снижение возбудимости нервной и мышечной тканей непосредственно вслед за потенциалом действия. Время рефрактерности – время, за которое возникает новый потенциал, влияет на относительную проницаемость, связанную с действием калиевого канала.
39. Чем обусловлено проведение импульса в немиелинизированных нервных волокнах? Скорость проведения импульса в немиелинизированных нервных волокнах будет расти с увеличением радиуса (диаметра) нервного волокна,будет зависеть от колебательных свойств волокна (электрическая ёмкость и сопротивление мембраны). Импульс будет распространяться непрерывно, независимые (локальные) ионные потоки препятствуют затуханию импульса.
40. Какая величина равна расстоянию, на котором деполяризующий потенциал в нервном волокне уменьшается в е (
2,73) раз? Постоянная длины.
41. На что влияет постоянная времени (RC) мембраны нервного волокна? На скорость распространения нервного импульса. Медленное распространение связано с тем, что постоянная времени перезарядки мембраны Т = RC велика, так как велики емкость мембраны C и сопротивление R нервного волокна.
Что означает бездекрементное проведение возбуждения по нервному волокну? Это проведение возбуждения без существенного изменения величины нервного импульса.
43. Каким законом можно охарактеризовать упругие (пассивные) свойства мышцы? Закон Гука.
44. Что подразумевает изотоническое сокращение мышцы? Сокращение мышцы при постнагрузке.
45. Каков физический смысл максимальной скорости укорочения мышцы в уравнении Хилла? Максимальная скорость – при нулевой нагрузке. Максимальная сила при нулевой скорости – величина изометрического сокращения мышцы.
46. Какую фундаментальную зависимость в мышце описывает уравнение Хилла? Изменение скорости сокращения мышцы в зависимости от её нагрузки.
47. Каков физический смысл закона Франка для физиологического сокращения мышцы? Приувеличении длинымышцы её активнаясила сокращения возрастает.
48. Какие параметры связывает зависимость в мышце сила-скорость? Изменение скорости укороченияот приложенной силы(величина постнагрузки – максимальная скорость сокращения).
49. Что подразумевает понятие гладкий тетанус в скелетной мышце? Тетанус - сильное и длительное сокращение мышц при достаточно высокой частоте их стимуляции. Гладкий тетанус – тетанус, при котором отсутствует хотя бы частичное расслабление мышцы.
50. Что является элементарной сократительной единицей мышечной клетки? Саркомер.
51. С каким химическим элементом связана АТФ в мышечной клетке? Mg.
При возбуждении нервного волокна можно зарегистрировать ПД как в месте раздражения, так и на расстоянии от него. На всем протяжении нервного волокна ПД имеет одинаковую амплитуду, но проявляется с задержкой, которая пропорциональна расстоянию от места нанесения стимула.
Распространение возбуждения по нерву:
Распространение ЭМВ с затуханием
В месте возникновения ПД потенциал внутренней стороны мембраны положителен, наружной- отрицателен. Т.к. распространение ЭМВ происходит со скоростью света, пассивные сдвиги трансмембранной разности потенциалов происходят быстро, и скорость распространения возбуждения по мембране зависит от величины участка, одновременно охваченного возбуждением
Величина деполяризующего потенциала:
Ux- Величина деполяризующего потенциала
U0-изменение мембранного потенциала в точке возбуждения
Х-расстояние от места возникновения возбуждения
Λ-постоянная длины мембраны
У позвоночных, которые имеют нервы с большим кол-вом проводящих волокон, возможность их утолщения ограничена размерами животного. Большая скорость передачи возбуждения обусловлена наличием миелиновой оболочки, которой снабжены аксоны. Она образуется в р-те наматывания на аксон шванновских клеток. Диффузия ионов невозможна, поэтому генерация ПД возможна только там, где миелиновая оболочка отсутствует(перехваты Ранвье). Мембрана перехвата Ранвье специализировна для генерации возбуждения, т.к. плотность натриевых потенциалзависимых каналов в 100 раз выше, чем в немиелинизорованных нервных волокнах. От перехвата к перехвату возбуждение распространяется за счет декрементного распр. ЭМВ. Ретрансляция происходит на 2-3 соседних перехватах ранвье, поэтому возбуждение как бы перепрыгивает через миелинизированные участки мембраны-сальтотарное проведение возбуждения. Миелинизация обеспечивает увеличение скорости проведения при существенной экономии энергетических ресурсов.
Бездекрементное движение ПД:
Декрементное распространение потенциала
Генерация нового ПД
В безмякотных нервных волокнах наблюдается непрерывное распространение возбуждения, в основе которого лежат локальные токи-токи, возникающие в живых тканях вследствие возбуждения, распространяющиеся на незначительные расстояния от возбуждения.
Сm-удельная емкость мембраны
Rm-удельное сопротивление мембраны
Λi-удельная электропроводность цитоплазмы
Н-фактор надежности(разность значений ПП и КМП)
Нервные и мышечные волокна имеют низкую электропроводность. В возбудимой мембране регенерированный ПД будет распространяться с декрементов,но,угасая сам,возбудит последующие участки волокна.
Бездекрементное движение ПД является р-том взаимодействия 2х процессов:
Декрементное распространение потенциала(от предыдущего ПД)
Генерация нового ПД
18) Функциональный межклеточный контакт, обеспечивающий переход возбуждения с одной клетки на другую, получил название синапса (от греч. Глагола "синапто" – смыкать).
Существует два принципиально различных типа синапсов – электрические и химические.
Электрическая синаптическая передача возможна только при очень тесном соприкосновении взаимодействующих клеток – при расстоянии между ними не более 10-20 нанометров (часто 2-4 нм).
В этом случае развитие ПД на мембране одной клетки приводит за счет возникновения локальных токов к деполяризации мембраны другой клетки, которая может оказаться выше порога генерирования ПД.
Большое значение для осуществления электрической передачи нервного импульса имеет существование в области синапса особых межклеточных контактов – щелевых контактов (нексусов).
При этом в каждой из двух соседних мембран находятся регулярно расположенные коннексоны (канальные белки с большим диаметром канала и, соответственно, высокой проводимостью для ионов, и даже более крупных молекул с молекулярной массой до 1000).

Такие контакты обычны для ЦНС, миокарда и гладкой мускулатуры, где связанные щелевыми контактами клетки образуют функциональный синцитий (возбуждение переходит от одной клетки к другой очень быстро и без заметного снижения амплитуды потенциала действия на границе).
Щелевые контакты регулируемы, они могут закрываться при снижении рН или повышении концентрации Са 2+ (повреждение клеток или глубокие нарушения обмена).
За счет такого механизма пораженные места изолируются от остальной части синцития, и распространение патологии ограничивается (инфаркт миокарда).
Химическая синаптическая передача осуществляется с помощью химических веществ-посредников (медиаторов).
В этом случае расстояние между взаимодействующими клетками в области контакта (ширина синаптической щели) больше. Электрическое поле затухает в пределах синаптической щели и не может деполяризовать постсинаптическую мембрану. Отсюда возникает необходимость химического посредника.
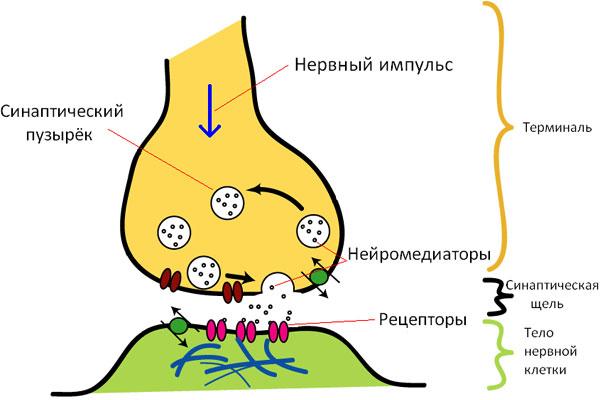
Деполяризация пресинапса приводит к изменению проницаемости пресинаптической мембраны для медиатора, медиатор выбрасывается в синаптическую щель, диффундирует через нее и взаимодействует с белками-рецепторами постсинаптической мембраны.
Изменение конформации белков-рецепторов при образовании комплекса "рецептор-медиатор" приводит к открытию на мембране специфических химиочувствительных ионных каналов, протекающие через которые ионные токи изменяют мембранный потенциал на мембране.
В зависимости от направления изменения трансмембранного потенциала химические синапсы могут быть возбуждающими (деполяризация постсинаптической мембраны) или тормозными (гиперполяризация постсинаптической мембраны).
Потенциал действия нервной клетки
- Потенциал действия нервной клетки
- Ионные механизмы потенциала действия
- Проведение возбуждения по нервным волокнам
- Законы проведения возбуждения
- Типы нервных волокон и их функции
- Все страницы
Проведение ПД по мембране можно сравнить с поджиганием пороховой дорожки: вспыхнувший порох немедленно воспламеняет впереди лежащие частицы, и пламя движется вперёд до конца дорожки.
Продолжительность потенциала действия нервной клетки измеряется единицами миллисекунд (мс).
Потенциалы действия, зарегистрированные двумя электродами, один из которых находится внутри клетки, а другой — в окружающем растворе, представлены на рис. 5-3 и 5-7.
Рис. 5–3. Изменения мембранного потенциала и потенциал действия. Вертикальная стрелка в нижней части рисунка — момент появления раздражающего стимула, на отметке –80 мВ — исходный уровень МП.
Между моментом нанесения раздражения и первым проявлением ПД имеется задержка — латентный период. Латентный период соответствует времени, когда ПД движется по мембране нервной клетки от места раздражения до отводящего электрода. Под действием раздражающего стимула происходит нарастающая деполяризация мембраны — локальный ответ. При достижении критического уровня деполяризации, который в среднем составляет —55 мВ, начинается фаза деполяризации. В эту фазу уровень МП падает до нуля и даже приобретает положительное значение (овершут), а затем возвращается к исходному уровню (фазареполяризации). Фазы деполяризации, овершута и реполяризации образуют спайк (пик) ПД. Длительность спайка составляет 1—2 мс. После спайка наблюдается замедление скорости спада потенциала — (раза следовой деполяризации. После достижения исходного уровня покоя нередко наблюдается фаза следовой гиперполяризации. Эти следовые потенциалы могут длиться десятки и сотни миллисекунд.
В основе изменений мембранного потенциала (МП), происходящих в течение потенциала действия (ПД), лежат ионные механизмы. На рис. 5—7 представлены суммарные ионные токи, протекающие через мембрану нервной клетки в ходе потенциала действия.
Рис. 5-7. Потенциал действия и ионные токи нервной клетки [5|. По оси ординат отложены значения МП (мВ), по оси абсцисс — время (мс)
Локальный потенциал. Раздражение клетки приводит к открытию части Na+-каналов и появлению локального (нераспространяющегося) потенциала.
Фаза деполяризации. При достижении критического уровня деполяризации мембраны (соответствует порогу активации потен-циалозависимых Na+-каналов) начинается лавинообразный процесс открытия большого количества Na+-каналов. В фазу деполяризации происходит массивный вход в клетку ионов Na+ по концентрационному и электрохимическому градиентам.
Овершут. Деполяризация мембраны приводит к реверсии МП (МП становится положительным). В фазу овершута Na+-ток начинает стремительно спадать, что связано с инактивацией потенциало-зависимых Na+-каналов (время открытого состояния –– доли миллисекунды) и исчезновением электрохимического градиента Na+.
Фаза реполяризации. Помимо инактивации Na+-каналов, развитию реполяризации способствует открытие потенциалозависимых K+-каналов. Этот процесс происходит медленнее, чем открытие Na+-каналов, но K+-каналы остаются открытыми более продолжительное время. Выход К+ наружу способствует полному завершению фазы реполяризации.
Следовые потенциалы связаны с длительными изменениями кинетических свойств К+-каналов. Восстановление исходного уровня МП приводит Na+- и K+-каналы в состояние покоя.
В ходе развития ПД происходят изменения возбудимости мембраны нервной клетки.
Абсолютно рефрактерный период. Во время фазы деполяризации и большей части фазы реполяризации ПД клетка находится в абсолютно рефрактерном периоде, в течение которого даже сверхпороговое раздражение не способно вызвать ПД. Этот феномен связан с инактивацией большинства Na+-каналов.
Относительно рефрактерный период. В конце фазы реполяризации, а также во время следовой гиперполяризации клетка способна генерировать ПД только в ответ на сверхпороговые раздражители. Это связано со значительным реполяризующим действием выходящих калиевых токов.
Наличие рефрактерности ограничивает частоту генерации ПД. Физиологическое значение рефрактерности заключается в создании условий для своевременного и полного осуществления восстановительных процессов в нервной клетке. Феномен рефрактерности лежит в основе понятия о функциональной подвижности, или лабильности (Н.Е. Введенский [1] ).
Лабильность — максимально возможная частота генерации ПД для данного типа возбудимой клетки. Лабильность большинства нейронов составляет приблизительно 400 ПД/с, а у интернейронов спинного мозга доходит до 1000 ПД/с.
В нормальных условиях рефрактерность предохраняет нервные клетки от излишне частой генерации ПД. Мутации, гипоксия, механические травмы и другие патологические воздействия приводят к значительным изменениям возбудимости нейронов. Такие нейроны или группы нейронов являются потенциальными источниками возникновения пароксизмальных состояний ЦНС, эпилептических припадков и других неврологических расстройств.
Нервные волокна — аксоны нервных клеток, окружённые оболочкой из олигодендроглиоцитов в ЦНС и шванновских [2] клеток в периферических нервах. Нервные волокна подразделяют на 2 типа — безмиелиновые и миелиновые. Основная функция нервных волокон — проведение ПД. Скорость проведения в миелиновых и безмиелиновых волокнах различна (рис. 5–8) и существенно зависит от диаметра нервных волокон.
Рис. 5–8. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокнах разного диаметра [4]. Скорость проведения пропорциональна диаметру нервного волокна и в миелиновых волокнах выше, чем в безмиелиновых.
Безмиелиновые нервные волокна (рис. 5–9А). В покое мембрана аксона (осевого цилиндра) поляризована — положительно заряжена снаружи и отрицательно внутри. При ПД полярность изменяется, и наружная поверхность мембраны приобретает отрицательный заряд. Из-за разности потенциалов между возбуждённым и невозбуждёнными сегментами возникают локальные токи, деполяризующие соседний участок мембраны. Теперь этот участок становится возбуждённым и деполяризует следующий участок мембраны.
Рис. 5–9. Проведение возбуждения в нервных волокнах [7]. А — безмиелиновое волокно (электротоническое проведение), Б — миелиновое волокно (скачкообразное проведение). Миелин, полностью окружая аксон в межузловых промежутках, выступает в роли электрического изолятора, а межклеточная жидкость в перехватах Ранвье [3] — проводник.
Появление так называемых рефрактерных каналов (рефрактерное состояние мембраны после прохождения ПД) предупреждает распространение возбуждения в обратном направлении.
Скорость проведения возбуждения по безмиелиновому нервному волокну в основном составляет 0,5–2 м/с и зависит от диаметра волокна: чем больше диаметр, тем выше скорость проведения ПД (см. рис. 5–8).
Миелиновое нервное волокно (рис. 5–9Б) состоит из осевого цилиндра (аксона), вокруг которого шванновские клетки образуют миелин за счёт концентрического наслаивания собственной плазматической мембраны. Миелин прерывается через регулярные промежутки (от 0,2 до 2 мм) концентрической щелью шириной около 1 мкм, это узлы, или перехваты Ранвье. Таким образом, межузловые сегменты аксона, расположенные между соседними перехватами Ранвье, содержат миелин — электрический изолятор, не позволяющий проходить через него локальным токам, поэтому ПД возникают только в перехватах Ранвье. Другими словами, ПД перемещается вдоль нервного волокна скачками, от одного перехвата Ранвье к другому перехвату (скачкообразное проведение).
Плотность потенциалозависимых Na+?каналов аксолеммы в перехватах Ранвье — до 2000 на 1 мкм2 (в перикарионе — 50–70, в начальном сегменте аксона — 2000, в межузловых сегментах Na+?каналы практически отсутствуют). В силу высокой плотности Na+?каналов перехваты Ранвье характеризуются высокой возбудимостью, а локальные токи достаточно велики для возбуждения соседнего перехвата.
Локальные токи текут от перехвата к перехвату (через внеклеточную жидкость кнаружи от миелина и через аксоплазму внутри аксона) с минимальными потерями.
Энергозатраты нервного волокна на проведение ПД относительно невелики, поскольку возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% общей поверхности мембраны аксона. Поэтому даже после длительных ритмических пачек ПД трансмембранный градиент концентраций ионов практически не изменяется.
В физиологических условиях ПД движутся в одном направлении от места раздражения (ортодромное проведение). ПД, проходящий по нервному волокну, возбуждает следующий, но не предыдущий участок мембраны. Это связано с рефрактерностью предыдущего участка после возбуждения. Проведение в противоположном направлении (антидромное проведение) возможно при травматическом поражении нервных волокон и в редких случаях (аксон–рефлекс).
Нарушение миелинизации нервных волокон приводит к нарушениям проводимости (демиелинизирующие заболевания). При разрушении миелиновой оболочки происходит резкое снижение скорости и надёжности проведения возбуждения по нервам. Наиболее распространённым среди демиелинизирующих заболеваний является множественный склероз, проявляющийся различными параличами и потерей чувствительности.
Бездекрементное проведение возбуждения. Амплитуда ПД в различных участках нерва одинакова, то есть проведение возбуждения по нервному волокну осуществляется без затухания (бездекрементно). Таким образом, кодирование информации осуществляется не за счёт изменения амплитуды ПД, а путём изменения их частоты и распределения во времени.
Изолированное проведение возбуждения. Нервные стволы обычно образованы большим количеством нервных волокон, однако ПД, идущие по каждому из них, не передаются на соседние. Эта особенность нервных волокон обусловлена:
- наличием оболочек, окружающих отдельные нервные волокна и их пучки (в результате образуется барьер, предупреждающий переход возбуждения с волокна на волокно);
- сопротивлением межклеточной жидкости (жидкость, находящаяся между волокнами, имеет гораздо меньшее сопротивление току, чем мембрана аксонов; поэтому ток шунтируется по межволоконным пространствам и не доходит до соседних волокон).
Физиологическая и анатомическая целостность. Необходимым условием проведения возбуждения является не только его анатомическая целостность, но и нормальное функционирование мембраны нервного волокна (физиологическая целостность). В клинике широко применяют различные ЛС, нарушающие физиологическую целостность нервных волокон. Так, эффекты местных анестетиков (новокаин, лидокаин, и др.) основаны на блокаде потенциалозависимых Na+?каналов. Нарушение физиологической целостности чувствительных нервных волокон вызывает анестезию (потерю чувствительности).
При регистрации электрической активности нервного ствола Джозеф Эрлангер и Герберт Гассер в 1937 г. обнаружили составной характер тока действия нервного ствола. На основании полученных данных (диаметр, скорость проведения, функция) разработана классификация (табл. 5—1), в соответствии с которой нервные волокна подразделены на группы А, В и С с дальнейшими градациями ( ? , ? , и т.д.).
Орлов Р.С., Ноздрачёв А.Д. Нормальная физиология : Учебник. - М.: ГЭОТАР-Медиа, 2009. 688 с. - Глава 5. Физиология нейронов. - Потенциал действия. С. 73-78.
[2] Шванн Теодор (Schwann T.), немецкий гистолог и физиолог (1810–1882); вместе с М. Шульце создал клеточную теорию (1839 г.); в 1836 г. открыл пепсин, в 1838 г. опубликовал первую работу по строению миелиновой оболочки.
[3] Ранвье Луи (Ranvier L.), французский патолог (1835–1922). Занимался изучением нервной ткани с применением азотнокислого серебра и хлорного золота. Его именем названы безмиелиновые участки (узловые перехваты) миелинового нервного волокна.
Читайте также:
