
Чем образована нервная система позвоночного животного образована
Тело разделено на множество систем. Какие элементы составляют нервную систему, и каковы их функции? Дышать, передвигаться, заниматься самыми разными делами без нее было бы невозможно. Это - система, которая получает и обрабатывает всю информацию, полученную со всех частей тела. Она состоит из головного мозга, спинного мозга и нейронов. Давайте охарактеризуем особенности строения нервной системы позвоночных. Пожалуй, самой важной.
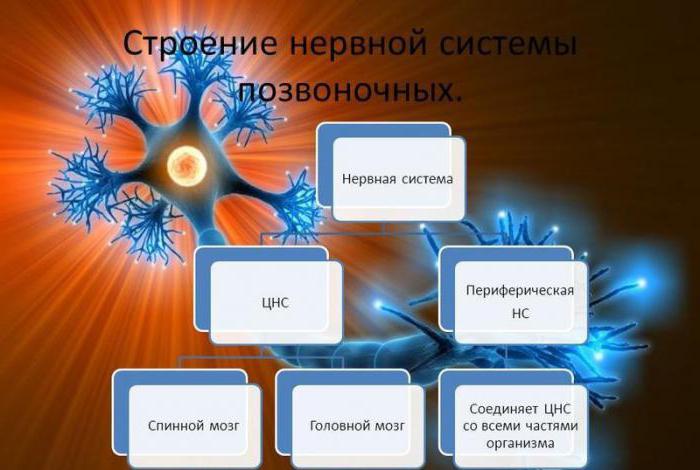
Части и функции центральной нервной системы
Итак, подробнее. Давайте охарактеризуем особенности строения нервной системы позвоночных. Ее основными единицами являются нейроны. Из них сделаны все клетки нервной системы. Нейроны способны передавать электрические импульсы по всему телу в виде сигналов, которые вызывают реакцию. Существует три типа нейронов:
- моторные передают информацию между органами, железами и мышцами.
- сенсорные посылают информацию в головной и спинной мозг из внутренних органов или внешних раздражителей, с которыми встречаются части тела.
- интернейроны выступают в качестве посланников, которые связывают сигналы между моторными и сенсорными нейронами.
Мозг - центр управления тела
Головной и спинной мозг являются двумя основными компонентами центральной нервной системы. Первый отвечает за принятие и обработку сенсорной информации: мышление, понимание, речь и моторные функции. Передний состоит из таламуса, гипоталамуса, головного мозга и коры головного мозга. Задний функционирует в поддержании баланса и равновесия, координации движения и обработки некоторой сенсорной информации, и имеет части, известные как мост, мозжечок и продолговатый мозг. Шток мозга простирается вплоть до спинного мозга и отвечает за слуховые и зрительные реакции, а также за двигательную функцию.
Строение нервной системы позвоночных включает также спинной мозг, который соединяется с головным, чтобы посылать информацию по телу в виде электрических сигналов. Он похож на толстую веревку. И состоит из нервов, которые передают сообщения между мозгом и остальной частью тела. Он простирается от продолговатого мозга до нижней части спины и размещается в пространстве, созданном позвонками или костями позвоночника. У всех позвоночных животных есть спинальные корды, от простых беззубых рыб до крупных птиц и млекопитающих.

Функции спинного мозга
Следующий момент. Охарактеризуем особенности строения нервной системы позвоночных на примере спинного мозга. Интересно то, что он работает наподобие оператора телефонного коммутатора. Помогает мозгу общаться с различными частями тела и наоборот. Его три основные роли:
- Передача сообщений из мозга в разные части тела (обычно в мышцы) для выполнения определенного действия.
- Перенос сообщений от сенсорных рецепторов, обнаруженных по всему телу, в мозг.
- Координация рефлексов (быстрые ответы на внешние раздражители), которые не проходят через мозг и управляются только спинным мозгом.
Общие характеристики
У взрослого человека спинной мозг имеет длину около 44 см и ширину - как у большого пальца. Он такой же тонкий, как соломка для для питья на конце. Толстый пучок нервов защищен тремя слоями мембраны. Между сердечником нервных пучков и мозговыми оболочками имеется также спинномозговая жидкость для дополнительной амортизации. Спинной мозг заканчивается каскадом нервов, напоминающих хвост лошади. Нервный шнур состоит из пяти основных областей, состоящих из 33 сегментов, каждый из которых содержит нервы, связанные с различными частями тела:
- Шейный край соединяется с головой, шеей, верхней частью тела, руками.
- Грудная область соединяется с руками, пальцами, грудью и мышцами живота.
- Поясничная область связана с бедрами, коленями, лодыжками и мышцами ног.
- Крестцовая область соединяется с ногами, пальцами ног, мочевым пузырем и анальными мышцами.
- Копчиковая область соединяется с кожей вокруг копчика.
Нервная система позвоночных животных: схема
И напоследок. Позвоночные разнообразны и широко распространены на Земле, но это не всегда было так. Трудно представить, но из-за определенных характеристик человек связан не только с другими млекопитающими, такими как собаки, зебры и кролики, но и с такими существами, как рыбы, змеи и саламандры. На самом деле, все животные, имеющие позвоночник, являются позвоночными . Это означает, что все они попадают в ту же категорию, что и люди.
Эти организмы представляют собой разнообразную группу животных, представляющих множество различных форм, размеров, цветов, мест обитания и экосистем. Понятно, что все они отличаются друг от друга. Однако существуют некоторые сходства, которые позволяют увидеть связь даже между теми организмами, внешность которых совершенно не похожа. Например, есть голова, две пары конечностей и так далее.
Охарактеризуем особенности строения нервной системы позвоночных, по сравнению с беспозвоночными. У первых совершенно другой тип построения. У беспозвоночных он более узкий и примитивный. ЦНС позвоночных имеет следующую особенность: с самых ранних этапов развития эмбриона для нее не является характерным наличие узлового строения.
Она имеет вид сплошной нервной трубки, дифференцирующейся на различные отделы мозга. Кроме центральной нервной системы есть также переферическая, где располагаются нервные узлы, функцией которых является соединение ЦНС со всеми частями организма.
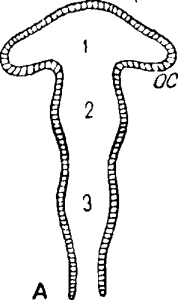
Закладка центральной нервной системы. У позвоночных животных центральная нервная система возникает в виде эктодермической нервной бороздки, расположенной на спинной стороне зародыша и ограниченной небольшими валиковидными возвышениями—нервными складками. Нервная бороздка, вследствие смыкания нервных складок, превращается в трубку. В заднем отделе образующейся нервной трубки некоторое время сохраняется отверстие, соединяющее последнюю с эмбриональными кишечником (canalis neurentericus), а спереди в описываемой нервной трубке можно отметить небольшое отверстие, ведущее наружу (н е в р о п о р).
Рассматривая при довольно больших увеличениях структуру нервной трубки, можно увидеть легкие перехваты, разделяющие трубку на ряд последовательных сегментов, или невромеров. Эта сегментация нервной системы, или н евромерия, является одним из показателей метамерии, свойственной зародышам позвоночных. В переднем, рано расширяющемся отделе нервной трубки невромеры также наблюдаютоя, но расположение их вскоре нарушается появлением новых подразделений, характерных для развивающегося головного мозга (рис.). Продолговато вздуткый мозговой пузырь разделяется перетяжками на три отдела: передний мозговой пузырь составляет первичный передний мозг (prosencephalon; рис, А1) средний пузырь образует средний мозг (mesencephalon; рис., А2), задний дает начало первичному заднему мозгу (rhombencephalon, рис, А3). Далее начинается новое усложнение, ведущее к образованию пяти пузырей (рис., В). От первичного переднего мозга спереди отделяется простой или парный выступ, дающий начало собственно переднему мозгу (telencephalon; рис., В, р). У огромного большинства позвоночных из этого переднего отдела развиваются полушария большого мозга, внутри заключающие полости двух рядом лежащих боковых желудочков.
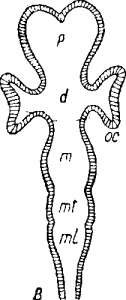
Рис. Развитие отде лов головного мозга. Схемы.
А—стадия трех пузырей (с глазными пузырями ос); В— стадия пяти отделов (с глазными бокалами ос), d—dien-cephalon; т—mesencephalon; ml — myelencephalon; mi — metencephalon; p — telencephalon.
От заднего отдела переднего пузыря отделяется новый отдел— промежуточный мозг (diencephalon; рис., d); его полость называется третьим желудочком. Средний пузырь не разделяется, а целиком образует упомянутый средний мозг (рис., m), в центре которого проходит узкий канал (так называемый сильвиев водопровод), соединяющий III желудочек с IV, заключенным в продолговатом мозгу (рис., ml). Здесь необходимо указать, что пер вичный задний (третий пузырь), подобно переднему, делится на два отдела : первый составляет упомянутый продолговатый мозг (myelencephalon), а второй образует задний мозг (metencephalon; рис., В1 mt), крыша которого выступает в виде поперечной, дорэально расположенной складки, давая начало мозжечку (cerebellum; рис., А, сbl).
Головной мозг позвоночных никогда не сохраняет структуры прямой трубки. Неравномерный рост отдельных частей мозга в связи с различным ходом утолщения его стенок ведет к характерному искривлению мозгового контура и к появлению так называемого мозгового перегиба. Проследим несколько внимательнее постепенную эволюцию в развитии каждого из упомянутых пяти отделов головного мозга (рис. 2).
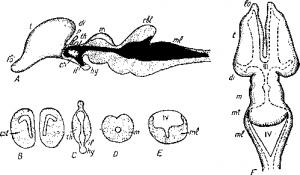
Как уже говорилось, передний мозг, достигающий различного развития у разных позвоночных, слагается из двух полушарий, из которых каждое имеет свой желудочек. В свою очередь каждый из этих желудочков сообщается с III желудочком промежуточного мозга посредством узкого монроева отверстия. На нижне-передних стенках полушарий ясно видной два выступа, образующие простирающиеся вперед обонятельные доли (рис. 2, F, l o), от которых берут начало обонятельные нервы. Дно каждого из полушарий образует заметное ганглиозное утолщение серого вещества, каждое из них называется полосатым телом (рис. 2, B, c st). Крыша полушарий слагается из мантии, состоящей из серого вещества и имеющей различную толщину. У низших позвоночных мантия весьма тонка и повторяет очертания лежащих под ней полосатых тел. У более высокоорганизованных форм мантия утолщается и наиболее сложна у млекопитающих. Мантия несет у них бугры и борозды, крайне осложняющие ее внешнюю структуру. Между полушариями переднего мозга устанавливается связь посредством соединений, носящих название комиссур. Так, в стенке (lamina terminalis), ограничивающей спереди III желудочек, развивается передняя комиссура. Впервые в мозгу амфибий и рептилий появляются и другие перемычки, достигающие особого развития у млекопитающих. У последних мы отмечаем мощную верхнюю комиссуру, или мозолистое тело (corpus callosum) и нижнюю—с вод (fornix).
Рис. 2. Схема головного мозга наземного позвоночного.
А —срединный разрез; B , С, D, Е —поперечные разрезы через передний, промежуточный, средний и продолговатый мозг; F —мозг сверху, ch— хиазма; сbl —мозжечок; est— полосатые тела; di —промежуточный мозг;. hy —гипофиз; if —воронка; 1о —-обонятельные доли; m — средний мозг; ml —продолговатый мозг; р— теменной орган; t —передний мозг; th —зрительные бугры; III—третий желудочек; IV—четвертый желудочек.
Переходим к обзору промежуточного мозга. Этот небольшой мозговой отдел виден сверху только у животных с примитивной структурой полушарий (например, у круглоротых, у рыб и др.). Обычно промежуточный мозг сжат или прикрыт смежными отделами. В промежуточном мозгу весьма утолщаются нижние боковые стенки, образующие зрительные бугры (thalami optici; рис.2, А, С, th); дно остается относительно тонким, а крыша не содержит нервного вещества и слагается из эпителиального слоя. Обычно он срастается с нижней, богатой сосудами оболочкой мозга. Тогда в крыше промежуточного мозга ясно обозначается сосудистое сплетение (plexus chorioideus). Верхняя стенка промежуточного мозга образует два лежащих друг за другом выступа: передний теменной, или париетальный, орган (рис 2, А, р) и задний— эпифиз. У круглоротых (миног) оба эти образования имеют характер глазоподобных органов, снабженных каждый непарным нервом. У ящериц такой жарактер сохраняет лишь теменной орган, который превращаетск в так называемый теменной глаз. У высших позвоночных удерживается лишь один эпифиз, который приобретает характер железы внутренней секреции.
Нижняя стенка промежуточного мозга образует направленный назад непарный полый выступ—воронку (рис. 2, if), к нижней стенке которого приле гаёт другая железа внутренней секреции—гипофиз, или подмозгова желез а (рис. 2, hy). Гипофиз возникает в виде отделяющегося выступа ротовой полости, прилегающего к воронке и входящего в состав последней.
Таким образом, гипофиз не может считаться производной частью мозга.
В гипофизе надо различать две самостоятельные части: 1) передний отдел, состоящий из эпителиальной ткани, и 2) задний, или инфундибулярный, отдел, слагающийся из нервной ткани и непосредственно переходящий в ткань мозга. Между обоими отделами залегает промежуточн ая часть.
Что касается гормонов, выделяемых гипофизом, то они распределяются таким образом.
1. Передний отдел выделяет:
a) гормон, влияющий на рост, развитие и функционирование половых желез у обоих полов (пролан);
b) гормон роста, обусловливающий общий рост тела, отдельных его тканей и органов;
c) гормон обмена веществ и усиления метаморфоза (тиреотропный гормон).
Известно, что при опухоли переднего отдела гипофиза и при разрастании этого отдела начинается чрезмерный рост организма, особенно резко сказывающийся в крайнем удлинении конечностей; иногда при этом утолщается слой подкожной клетчатки, увеличиваются в размерах кисти рук и ног. Этот болезненный гигантизм носит название акромегалии. В некоторых случаях к гигантизму присоединяется еще и половое бессилие. Напротив, недоразвитие переднего отдела гипофиза влечет за собой карликовое развитие организма.
Промежуточная часть продуцирует гормоны, влияющие на пигментные клетки организма (меланофорный гормон), на сон и психические функции (бромсодержащий гормон Цондека).
2. Задний отдел выделяет:
а) гормон, вызывающий сокращение матки (орастин); b) гормон, суживающий сосуды и повышающий кровяное давление (тонефин).
В медицинском отношении особенно важен гормон, действующий на матку. Врачебная практика уже давно имеет дело с опасным и мучительным явлением— ослаблением деятельности матки во время родов. Введение питуитрина—препарата из задней доли гипофиза—вызывает усиленное сокращение матки и дает возможность нормального окончания родов.
Средний мозг у низших позвоночных (например, у круглоротых, у рыб) относительно велик. Его крыша вздута в парные зрительные доли. Полость среднего мозга низших позвоночных довольно заметна и продолжается в два боковых желудочка, расположенных в зрительных долях. У высших позвоночных вследствие утолщения стенок крайне сокращается внутренняя полость описываемого мозгового отдела и превращается в узкий канал—уже упомянутый нами сильвиев водопровод. У млекопитающих крыша среднего мозга несет четыре бугра (четверохолмие).
В продолговатом мозгу чрезвычайно утолщается его дно, где образуются так называемое пирамиды (рис. 2, Е, ml), которые состоят из белого вещества, служащего продолжением наружных стенок спинного мозга. Верхнюю стенку продолговатого мозга составляет заднее сосудистое сплетение, состоящее из слоя эпителия (эпендимы) и тончайшей сети кровеносных сосудов. Если удалить пленчатую крышу продолговатого мозга, то ясно обнаруживается IV желудочек с полостью в виде ромбоидальной ямки (рис.2, IV). В передней части продолговатого мозга, вдоль края крыши IV желудочка, заметны тяжи продольных волокон, переходящих на мозжечок,— это так называемые веревочные тела.
Мозжечок отличается различными размерами и структурой. У круглоротых и у амфибий он имеет характер небольшой поперечной складки, спереди ограничивающей крышу IV желудочка, а у многих рыб (поперечноротых) достигает мощного развития. У птиц и млекопитающих на поверхности средней части мозжечка, носящей название червячка (vermis), можно различить многочисленные ряды поперечных складок; по бокам мозжечка у них выдаются парные выступы. На продольном разрезе мозжечка можно отчетливо видеть центральный слой белого вещества, имеющий очертание разветвленного дерева (древо жизни, arbor vitae). Наружный слой серого вещества заполняет промежутки между разветвлениями центральной белой части.
Статья на тему Нервная система позвоночных
Позвоночных считают высшим подтипом типа Хордовые. В то время как оболочников и бесчерепных — низшими хордовыми. Позвоночных животных насчитывается более 40 тысяч видов. Они разнообразны по строению, размерам, жизнедеятельности, средам обитания. При этом имеют ряд общих черт, особенно в период эмбрионального развития, что говорит об общности их эволюционного происхождения.
Почти все позвоночные животные имеют высокоразвитую нервную систему и ведут активный образ жизни (ищут пищу и партнеров для размножения, убегают от опасности).
Первые обнаруженные остатки позвоночных относятся к силуру.
К позвоночным принадлежат: круглоротые, хрящевые и костные рыбы, земноводные, пресмыкающиеся, птицы и млекопитающие (звери). Круглоротые являются бесчелюстными. Остальные классы подтипа относятся к разделу Челюстноротые.
Опорно-двигательная система позвоночных
Ароморфозы: формирование осевого скелета в форме позвоночного столба; появление черепа для защиты головного мозга; развитие челюстей для схватывания добычи и, у более высокоорганизованных, измельчения пищи; возникновение парных конечностей, позволяющих быстро перемещаться в пространстве.
Скелет позвоночных хрящевой или костный (у большинства). Его главные функции — это обеспечение движения животного и защита его внутренних органов. Кроме этого кости скелета служат местом прикрепления мускулатуры тела, в отдельных костях происходит кроветворение, осуществляется хранение ряда веществ.
Позвоночник образуется на основе хорды. У ряда видов позвоночных (миног) хорда сохраняется во взрослом состоянии, но около нее развиваются хрящи, защищающие спинной мозг. У осетровых вокруг хорды образуются верхние и нижние дуги позвонков.
У большинства позвоночных позвоночник состоит из отдельных относительно подвижных друг относительно друга позвонков. Каждый позвонок имеет тело, верхнюю и нижнюю дуги. Через канал верхней дуги проходит спинной мозг. Дуги позвонков служат защитой спинному мозгу. К позвонкам крепятся ребра, защищающие органы грудной полости.
Скелет позвоночных делят на:
Осевой скелет — позвоночник и мозговой череп.
Висцеральный скелет — жаберные дуги и кости произошедшие от жаберных дуг (челюсти и некоторые другие).
Скелет конечностей и их поясов (за исключением миног и миксин).
Конечности бывают двух основных типов — плавник и пятипалая конечность. В плавнике хрящи или кости конечности перемещаются относительно своего пояса как единый рычаг. Пятипалая конечность наземных животных представляет собой ряд рычагов, независимо перемещающихся относительно друг друга и пояса конечности.
Мускулатура тела образована поперечно-полосатыми мышцами. У высших позвоночных (пресмыкающиеся, птицы, млекопитающие) мышцы разделены на отдельные пучки. У низших позвоночных мышцы носят сегментированный характер.
Есть гладкая мускулатура внутренних органов. Ее называют висцеральной.
Нервная система и органы чувств позвоночных
Ароморфозы: образование головного мозга, подразделение его на пять отделов, выполняющих разные функции (передний, промежуточный, средний, продолговатый мозг и мозжечок).
Нервная трубка у позвоночных дифференцируется на спинной и головной мозг, которые вместе образуют центральную нервную систему. Кроме нее выделяют периферическую, симпатическую, парасимпатическую и вегетативную нервную системы.
Развитый головной мозг обеспечивает сложное поведение, в том числе коллективное. Высшая нервная деятельность является основой приспособительного поведения.
Невроцель (полосто внутри нервной трубки) в головном мозге превращается в желудочки мозга. От головного мозга отходят 10-12 пар нервов (обонятельные, зрительные, глазодвигательные, блоковые, тройничные, отводящие, лицевые, слуховые, языкоглоточные, блуждающие, добавочные, подъязычные). От спинного мозга нервы отходят попарно.
Органы чувств обеспечивают связь организма с внешней средой. У позвоночных они многообразны и имеют сложное строение. Глаза с хрусталиком, форма которого у наземных позвоночных может меняться. У рыб для достижения четкости изображения хрусталик может перемещаться.
Органы слуха связаны с органами равновесия. У различных групп позвоночных имеют разное строение. Обонятельная полость открывается наружу ноздрями. В коже имеются рецепторы осязания, температурные, давления и др.
Кровеносная и сердечно-сосудистая система позвоночных
Ароморфозы: появление сердца, обеспечивающего быстрый кровоток; полное разделение артериального и венозного кровотока у птиц и млекопитающих, в следствие этого появление теплокровности, позволившая животным меньше зависеть от неблагоприятных условий абиотической среды.
Для позвоночных, как и всех хордовых, характерна замкнутая кровеносная система.
Количество камер сердца (от 2 до 4) зависит от уровня организации класса. Низшие позвоночные имеют один круг кровообращения. В таком случае через сердце проходит венозная кровь, которая далее идет к жабрам, где насыщается кислородом, далее артериальная кровь разносится по всему телу. Легочный (второй) круг кровообращения впервые появляется у амфибий (земноводных).
Кровь позвоночных состоит из плазмы, в которой находятся красные и белые кровяные клетки.
Кожный покров позвоночных
Ароморфоз: появление двухслойной кожи.
Поверхностный слой кожи — многослойный эпидермис. В нем развиваются различные железы (потовые, сальные, слизистые и др.) и ряд твердых образований (когти, волосы, перья, чешуя). Внутренний слой кожи — дерма, представляющей собой прочную соединительную ткань. Здесь также образуются такие твердые образования как костная чешуя, кожные (накладные) кости.
Пищеварительная система позвоночных
В пищеварительной системе позвоночных выделяют пять отделов: ротовая полость, глотка, пищевод, желудок, тонкая кишка, толстая кишка. В процессе эволюции кишечник постепенно удлинялся.
Пищеварительные железы: слюнные, печень, поджелудочная железа.
Дыхательная система позвоночных
Жабры у круглоротых, рыб и личинок земноводных. Легкие — у всех остальных позвоночных. У низших позвоночных большую роль играет кожное дыхание.
Жабры представляют собой пластинчатые выросты стенок жаберных щелей. В таких пластинах находится сеть мелких кровеносных сосудов.
В процессе эмбрионального развития легкие образуются как пара выростов глотки. У земноводных и пресмыкающихся легкие мешкообразные. У птиц имеют губчатое строение. У млекопитающих разветвления бронхов оканчиваются альвеолами (мелкими пузырьками).
Выделительная система позвоночных
Органами выделения позвоночных служит пара почек. Почки имеют разное строение у разных групп позвоночных. Бывают головные, туловищные, тазовые почки. В процессе эмбриогенеза происходит смена головных на туловищные или туловищных на тазовые.
Половая система и эмбриональное развитие позвоночных
Почти все виды позвоночных раздельнополы. Имеются парные половые железы (семенники или яичники). За исключением круглоротых, у остальных есть специальные протоки, выводящие половые продукты.
Челюстноротых разделяют на две группы: анамнии и амниоты. К анамниям относятся рыбы и земноводные, так как их личиночная стадия живет в воде, а развитие эмбриона проходит без образования специальных зародышевых оболочек. Для анамний обычно наружнее оплодотворение.
К амниотам относятся пресмыкающиеся, птицы и звери. Их эмбрион имеет зародышевые оболочки (амнион и аллантоис). Характерно внутреннее оплодотворение.
Нервная система позвоночных закладывается в виде сплошной нервной трубки, которая в процессе онто- и филогенеза дифференцируется на различные отделы и является также источником периферических симпатических и парасимпатических нервных узлов. У наиболее древних хордовых (бесчерепных) головной мозг отсутствует и нервная трубка представлена в малодифференцированном состоянии.
Согласно представлениям Л. А. Орбели, С. Херрика, А. И. Ка-рамяна, этот критический этап развития центральной нервной си-
стемы обозначается как спинальный. Нервная трубка современного бесчерепного (ланцетника), как и спинной мозг более высоко организованных позвоночных, имеет метамерное строение и состоит из 62—64 сегментов, в центре которых проходит спинно-мозговой канал. От каждого сегмента отходят брюшные (двигательные) и спинные (чувствительные) корешки, которые не образуют смешанных нервов, а идут в виде отдельных стволов. В головных и хвостовых отделах нервной трубки локализованы гигантские клетки Родэ, толстые аксоны которых образуют проводниковый аппарат. С клетками Родэ связаны светочувствительные глазки Гесса, возбуждение которых вызывает отрицательный фототаксис .
В головной части нервной трубки ланцетника находятся крупные ганглиозные клетки Овсянникова, имеющие синаптические контакты с биполярными чувствительными клетками обонятельной ямки. В последнее время в головной части нервной трубки идентифицированы нейросекреторные клетки, напоминающие гипофи-зарную систему высших позвоночных. Однако анализ восприятия и простых форм обучения ланцетника показывает, что на данном этапе развития ЦНС функционирует по принципу эквипотенци-альности, и утверждение о специфике головного отдела нервной трубки не имеет достаточных оснований.
В ходе дальнейшей эволюции наблюдается перемещение некоторых функций и систем интеграции из спинного мозга в головной — процесс энцефализации, который был рассмотрен на примере беспозвоночных животных. В период филогенетического развития от уровня бесчерепных до уровня круглоротых формируется головной мозг как надстройка над системами дистантной рецепции.
Исследование ЦНС современных круглоротых показывает, что их головной мозг в зачаточном состоянии содержит все основные структурные элементы. Развитие вестибулолатеральной системы, связанной с полукружными каналами и рецепторами боковой линии, возникновение ядер блуждающего нерва и дыхательного центра создают основу для формирования заднего мозга. Задний мозг миноги включает продолговатый мозг и мозжечок в виде небольших выпячиваний нервной трубки.
Развитие дистантной зрительной рецепции дает толчок к закладке среднего мозга. На дорсальной поверхности нервной трубки развивается зрительный рефлекторный центр — крыша среднего мозга, куда приходят волокна зрительного нерва. И наконец, развитие обонятельных рецепторов способствует формированию переднего или конечного мозга, к которому примыкает слаборазвитый промежуточный мозг.
Указанная выше направленность процесса энцефализации согласуется с ходом онтогенетического развития мозга у круглоротых. В процессе эмбриогенеза головные отделы нервной трубки дают начало трем мозговым пузырям. Из переднего пузыря формируется конечный и промежуточный мозг, средний пузырь дифференцируется в средний мозг, а из заднего пузыря образуются продолговатый
мозг и мозжечок. Сходный план онтогенетического развития мозга сохраняется и у других классов позвоночных.
Нейрофизиологические исследования мозга круглоротых показывают, что его главный интегративный уровень сосредоточен в среднем и продолговатом мозгу, т. е. на данном этапе развития ЦНС доминирует бульбомезенцефальная система интеграции, пришедшая на смену спинальной.
Передний мозг круглоротых длительное время считали чисто обонятельным. Однако исследования недавнего времени показали, что обонятельные входы в передний мозг не являются единственными, а дополняются сенсорными входами других модальностей. Очевидно, уже на ранних этапах филогенеза позвоночных передний мозг начинает участвовать в переработке информации и управлении поведением.
Вместе с тем энцефализация как магистральное направление развития мозга не исключает эволюционных преобразований в спинном мозгу круглоротых. В отличие от бесчерепных нейроны кожной чувствительности выделяются из спинного мозга и концентрируются в спинно-мозговой ганглий. Наблюдается совершенствование проводниковой части спинного мозга. Проводящие волокна боковых столбов имеют контакты с мощной дендритной сетью мотонейронов. Формируются нисходящие связи головного мозга со спинным через мюллеровские волокна — гигантские аксоны клеток, лежащих в среднем и продолговатом мозгу.
Появление более сложных форм двигательного поведения у позвоночных сопряжено с совершенствованием организации спинного мозга. Так, например, переход от стереотипных ундулирующих движений круглоротых к локомоции с помощью плавников у хрящевых рыб (акулы, скаты) связан с разделением кожной и мышеч-но-суставной (проприоцептивной) чувствительности . В спинальных ганглиях появляются специализированные нейроны для выполнения этих функций.
В эфферентной части спинного мозга хрящевых рыб также наблюдаются прогрессивные преобразования. Укорачивается путь моторных аксонов внутри спинного мозга, происходит дальнейшая дифференциация его проводящих путей. Восходящие пути боковых столбов у хрящевых рыб доходят до продолговатого мозга и мозжечка. Вместе с тем восходящие пути задних столбов спинного мозга еще не дифференцированы и состоят из коротких звеньев.
Нисходящие пути спинного мозга у хрящевых рыб представлены развитым ретикулоспинальным трактом и путями, соединяющими вестибулолатеральную систему и мозжечок со спинным мозгом (вестибулоспинальный и церебеллоспинальный тракты).
Одновременно в продолговатом мозгу наблюдается усложнение системыядер вестибулолатеральной зоны. Этот процесс сопряжен с дальнейшей дифференциацией органов боковой линии и с появлением в лабиринте третьего (наружного) полукружного канала в Дополнение к переднему и заднему.
Развитие общей двигательной координации у хрящевых рыб связано с интенсивным развитием мозжечка. Массивный мозжечок акулы имеет двусторонние связи со спинным, продолговатым мозгом и покрышкой среднего мозга. Функционально он разделяется на две части: старый мозжечок (архицеребеллум), связанный с вестибуло-латеральной системой, и древний мозжечок (палецоцеребеллум), включенный в систему анализа проприоцептивной чувствительности. Существенным моментом структурной организации мозжечка хрящевых рыб является его многослойность. В сером веществе мозжечка акулы идентифицированы молекулярный слой, слой клеток Пуркинье и зернистый слой.
Другой многослойной структурой стволовой части мозга хрящевых рыб является крыша среднего мозга, куда подходят афференты различных модальностей (зрительные, соматические). Сама морфологическая организация среднего мозга свидетельствует о его важной роли в интегративных процессах на данном уровне филогенетического развития.
В промежуточном мозгу хрящевых рыб происходит дифференциация гипоталамуса, который является наиболее древним образованием этой части мозга. Гипоталамус имеет связи с конечным мозгом. Сам конечный мозг разрастается и состоит из обонятельных луковиц и парных полушарий. В полушариях у акул находятся зачатки старой коры (архикортекса) и древней коры (палеокортекса).
Палеокортекс, тесно связанный с обонятельными луковицами, служит главным образом для восприятия обонятельных стимулов. Архикортекс, или гиппокампальная кора, предназначен для более сложной обработки обонятельной информации. Вместе с тем электрофизиологические исследования показали, что обонятельные проекции занимают только часть полушарий переднего мозга акул. Кроме обонятельной здесь обнаружено представительство зрительной и соматической сенсорных систем. Очевидно, старая и древняя кора может участвовать в регуляции поисковых, пищевых, половых и оборонительных рефлексов у хрящевых рыб, многие из которых являются активными хищниками.
Таким образом, у хрящевых рыб складываются основные черты ихтиопсидного типа организации мозга. Его отличительной чертой является присутствие надсегментарного аппарата интеграции, координирующего работу моторных центров и организующего поведение. Эти интегративные функции осуществляют средний мозг и мозжечок, что позволяет говорить о мезэнцвфалоцеребеллярной системе интеграции на данном этапе филогенетического развития нервной системы. Конечный мозг остется преимущественно обонятельным, хотя и участвует в регуляции функций нижележащих отделов.
Переход позвоночных от водного к наземному образу жизни связан с целым рядом перестроек в ЦНС. Так, например, у амфибий в спинном мозгу появляется два утолщения, соответствующие верхнему и нижнему поясам конечностей. В спинальных ганглиях вместо биполярных чувствительных нейронов сосредоточиваются униполярные с Т-образно ветвящимся отростком, обеспечивающим более высокую скорость проведения возбуждения без участия клеточного тела. На периферии в коже земноводных формируются специализированные рецепторы и рецепторные поля, обеспечивающие дискриминационную чувствительность.
В мозговом стволе также происходят структурные изменения в связи с перераспределением функциональной значимости различных отделов. В продолговатом мозгу наблюдаются редукция ядер боковой линии и формирование кохлеарного, слухового ядра, осуществляющего анализ информации от примитивного органа слуха .
По сравнению с рыбами у амфибий, имеющих довольно стереотипную локомоцию, наблюдается значительная редукция мозжечка. Средний мозг, так же как и у рыб, представляет собой многослойную структуру, в которой наряду с передним двухолмием — ведущим отделом интеграции зрительного анализатора — появляются дополнительные бугорки — предшественники задних холмов четверохолмия.
Наиболее существенные в эволюционном плане изменения происходят в промежуточном мозгу амфибий. Здесь обособляется зрительный бугор — таламус, появляются структурированные ядра (наружное коленчатое тело) и восходящие пути, связывающие зрительный бугор с корой (таламокортикальный тракт).
В полушариях переднего мозга происходит дальнейшая дифференциация старой и древней коры. В старой коре (архикортексе) обнаруживаются звездчатые и пирамидные клетки. В промежутке между старой и древней корой появляется полоска плаща, которая является предтечей новой коры (неокортекса).
В целом развитие переднего мозга создает предпосылки для перехода от свойственной рыбам мозжечково-мезэнцефальной системы интеграции к диэнцефало-телэнцефальной, где ведущим отделом становится передний мозг, а зрительный бугор промежуточного мозга превращается в коллектор всех афферентных сигналов. В полной мере эта система интеграции представлена в зауропсидном типе мозга у рептилий и знаменует собой следующий этап мор-фофункциональной эволюции мозга .
Развитие таламокортикальной системы связей у рептилий приводит к формированию новых проводящих путей, как бы подтягивающихся к филогенетически молодым формациям мозга.
В боковых столбах спинного мозга рептилий появляется восходящий спиноталамический тракт, который проводит к головному мозгу информацию о температурной и болевой чувствительности. Здесь же в боковых столбах формируется новый нисходящий тракт — руброспинальный (Монакова). Он связывает мотонейроны спинного мозга с красным ядром среднего мозга, которое включено в древнюю экстрапирамидную систему двигательной регуляции. Эта многозвенная система объединяет влияние переднего мозга, мозжечка, ретикулярной формации ствола, ядер вестибулярного комплекса и координирует двигательную активность.
У рептилий, как истинно наземных животных, возрастает роль зрительной и акустической информации, возникает необходимость
сопоставления этой информации с обонятельной и вкусовой, В соответствии с этими биологическими изменениями в стволовой части мозга рептилий происходит целый ряд структурных изменений. В продолговатом мозгу дифференцируются слуховые ядра, помимо кохлеарного ядра появляется угловое, связанное со средним мозгом. В среднем мозгу двухолмие преобразуется в четверохолмие, в задних холмах которого локализованы акустические центры.
Наблюдается дальнейшая дифференциация связей крыши среднего мозга со зрительным бугром — таламусом, который является как бы преддверием перед входом в кору всех восходящих сенсорных путей. В самом таламусе происходит дальнейшее обособление ядерных структур и установление между ними специализированных связей.
Конечный мозг рептилий может иметь два типа организации:
кортикальный и стриатальный. Кортикальный тип организации, свойственный современным черепахам, характеризуется преимущественным развитием полушарий переднего мозга и параллельным 'развитием новых отделов мозжечка. В дальнейшем это направление в эволюции мозга сохраняется у млекопитающих.
Стриатальный тип организации, характерный для современных ящериц, отличается доминирующим развитием находящихся в глубине полушарий базальных ганглиев, в частности полосатого тела. По этому пути далее идет развитие головного мозга у птиц. Представляет интерес, что в полосатом теле у птиц имеются клеточные объединения или ассоциации нейронов (от трех до десяти), разделенные олигодендроглией. Нейроны таких ассоциаций получают одинаковую афферентацию, и это делает их сходными с нейронами, объединенными в вертикальные колонки в новой коре млекопитающих. В то же время в полосатом теле млекопитающих идентичные клеточные ассоциации не описаны. Очевидно, это является примером конвергентной эволюции, когда сходные образования развились независимо у различных животных.
У млекопитающих развитие переднего мозга сопровождалось бурным ростом новой коры, находящейся в тесной функциональной связи со зрительным бугром промежуточного мозга . В коре закладываются эфферентные пирамидные клетки, посылающие свои длинные аксоны к мотонейронам спинного мозга.
Таким образом, наряду с многозвенной экстрапирамидной системой появляются прямые пирамидные пути, которые обеспечивают непосредственный контроль над двигательными актами. Корковая регуляция моторики у млекопитающих приводит к развитию филогенетически наиболее молодой части мозжечка — передней части задних долей полушарий, или неоцеребеллума. Неоцеребеллум приобретает двусторонние связи с новой корой.
Рост новой коры у млекопитающих происходит настолько интенсивно, что старая и древняя кора оттесняется в медиальном направлении к мозговой перегородке. Бурный рост коры компенсируется формированием складчатости. У наиболее низко организованных однопроходных (утконос) на поверхности полушария закладываются первые две постоянные борозды, остальная же поверхность остается гладкой (лиссэнцефальный тип коры).
Как показали нейрофизиологические исследования, мозг однопроходных и сумчатых млекопитающих лишен еще соединяющего полушария мозолистого тела и характеризуется перекрытием сенсорных проекций в новой коре. Четкая локализация моторных, зрительных и слуховых проекций здесь отсутствует.
У плацентарных млекопитающих (насекомоядных и грызунов) * отмечается развитие более четкой локализации проекционных зон в коре. Наряду с проекционными зонами в новой коре формируются ассоциативные зоны, однако границы первых и вторых могут перекрываться. Мозг насекомоядных и грызунов характеризуется наличием мозолистого тела и дальнейшим увеличением общей площади новой коры.
В процессе параллельно-адаптивной эволюции у хищных млекопитающих появляются теменные и лобные ассоциативные поля, ответственные за оценку биологически значимой информации, мотивацию поведения и программирование сложных поведенческих актов. Наблюдается дальнейшее развитие складчатости новой коры.
И наконец, приматы демонстрируют наиболее высокий уровень организации коры головного мозга. Кора приматов характеризуется шестислойностью, отсутствием перекрытия ассоциативных и проекционных зон. У приматов формируются связи между фронтальными и теменными ассоциативными полями и, таким образом, возникает целостная интегративная система больших полушарий.
В целом, прослеживая основные этапы эволюции мозга позвоночных, следует отметить, что его развитие не сводилось просто к линейному увеличению размеров. В различных эволюционных линиях позвоночных могли иметь место независимые процессы увеличения размеров и усложнения цитоархитектоники различных отделов мозга. Примером тому может служить сравнение стриа-тального и кортикального типов организации переднего мозга позвоночных.
В процессе развития наблюдается тенденция к перемещению ведущих интегративных центров мозга в ростральном направлении от среднего мозга и мозжечка к переднему мозгу. Однако эту тенденцию нельзя абсолютизировать, так как мозг представляет собой целостную систему, в которой стволовые части играют важную функциональную роль на всех этапах филогенетического развития позвоночных. Кроме того, начиная с круглоротых в переднем мозгу обнаруживаются проекции различных сенсорных модальностей, свидетельствующие об участии этого отдела мозга в управлении поведением уже на ранних стадиях эволюции позвоночных.
Список литературы
1. Самусев Р.П. Анатомия человека.М., 1995.
2. Анатомия человека/ Под ред. М.Р. Сапина. М.,1986.
3. Общий курс физиологии человека и животных в 2-х кн. Под ред. А.Д.Ноздрачёва. М., “Высшая школа”,1991.
Читайте также:
