
Что такое ганглий и анализатор
Висцеральный анализатор. интерорецепторы
Боль. ноцицептивный анализатор
Физиология анализаторов
Анализатор - это совокупность рецепторов и нейронов мозга, участвующих в обработке информации о сигналах внешнего или внутреннего мира и в получении о них представления (ощущения, восприятия). Все анализаторы состоят из трех основных отделов: периферического (в нем происходит превращение сигнала внешнего мира в электрический процесс), проводникового - в нем происходит обработка информации и проведение ее в высшие отделы мозга и, наконец, центрального или коркового отдела, в котором происходит окончательная обработка сенсорной информации и возникает ощущение - субъективный образ сигнала.
Рассмотрим принцип работы анализатора.
Рецептор - это специализированная структура (клетка или окончание нейрона), которая в процессе эволюции приспособилась к восприятию соответствующего раздражителя внешнего или внутреннего мира. Например, адекватным раздражителем для фоторецепторов является квант видимого света, для фонорецепторов - звуковые колебания воздушной или водной среды, для терморецепторов - воздействие температуры. Под влиянием адекватного раздражителя в рецепторной клетке или в нервном специализированном окончании происходит изменение проницаемости для ионов (например, под влиянием растяжения в рецепторе растяжения мышц рака происходит увеличение открытия натриевых каналов, что вызывает деполяризацию, степень которой пропорциональна степени растяжения), что приводит к генерации рецепторного потенциала.
Афферентные нейроны - это первые нейроны, которые участвуют в обработке сенсорной информации. Как правило, афферентные нейроны лежат в ганглиях (спинномозговые ганглии, ганглии головы и шеи, например, вестибулярный ганглий, спиральный ганглий, коленчатый ганглий и т.п.). Исключением являются фоторецепторы - их афферентные нейроны лежат непосредственно на сетчатке.
Следующий нейрон, принимающий участие в обработке информации, расположен в спинном, продолговатом или в среднем мозге. Отсюда идут пути к таламусу. Исключением из этого правила является обонятельный анализатор - после обонятельной луковицы информация направляется сразу же в обонятельную кору, не заходя в таламус. От общего сенсорного коллектора (таламуса) информация поступает в соответствующие проекционные и ассоциативные зоны коры. Для каждого анализатора имеются свои конкретные участки, куда приходят импульсы от рецепторного аппарата. В проекционных зонах происходит декодирование информации, возникает представление о модальности сигнала, о его силе и качестве, а в ассоциативных участках коры - определение "что это такое?" - акцепция сигнала. Это происходит с участием процессов памяти.
Высшие отделы наряду с собственными рецепторными механизмами регулируют и процессы адаптации в рецепторах - привыкание. В основном, все рецепторы - быстро адаптирующиеся, поэтому они реагируют на начало воздействия стимула и на окончание его действия. Часть рецепторов - медленно адаптирующиеся, поэтому постоянно реагируют на стимул. Например, быстро адаптируются рецепторы обоняния, вкуса, но медленно адаптируются рецепторы боли (ноцицепторы).
Проприоцептивная чувствительность. мышечные рецепторы
Если закрыть глаза и попытаться написать текст, то буквы будут все-таки написаны достаточно четко. Таким способом просто убедиться, что мы умеем пользоваться информацией, идущей от мышц и суставов. Известно, что имеется 3 вида рецепторов, осуществляющих восприятие положения мышцы, ее состояние и положение сустава: это мышечные веретена, сухожильные рецепторы Гольджи и суставные рецепторы.
Мышечные веретена представляют собой инкапсулированные мышечные волокна (интрафузальныс волокна), на которые намотаны нервные волокна, представляющие собой окончания дендритов афферентного нейрона, расположенного в спинномозговом ганглии. Это рецепторы растяжения: когда мышца растягивается под влиянием силы тяжести или искусственно (например, ударом неврологического молоточка), то возникает растяжение мышечных волокон веретена, что влечет за собой растяжение нервных окончаний.
Сухожильные рецепторы тоже относятся к первичным рецепторам: в момент сокращения мышц в этих рецепторах возникает деполяризация, величина которой пропорциональна силе, развиваемой мышцей.
Вестибулярный анализатор
Адекватным раздражителем для рецепторов вестибулярного аппарата - для волосковых клеток макул (они расположены в вестибулюме) и волосковых клеток гребешков (находятся в расширенной части ампул полукружных каналов) являются соответственно линейное и угловое ускорения (ускорение Кориолиса). Сигнал от рецепторов идет в продолговатый мозг. Здесь расположены 4 вестибулярных ядра.
От вестибулярных ядер продолговатого мозга начинаются пути:
1. Вестибулоспинальный, который передает информацию от вестибулярного аппарата на мотонейроны спинного мозга и тем самым способствует сохранению равновесия при движении.
2. Вестибулоокулярный путь - этот путь используется для регуляции активности мышц глаза во время движения. Благодаря этому, несмотря на всевозможные перемещения тела, на сетчатке сохраняется объект наблюдения.
3. Вестибуломозжечковый путь - идет к мозжечку и несет туда информацию о положении тела в пространстве.
4. Лемнисковый путь - от вестибулярных ядер информация идет также к специфическим ядрам таламуса (по лемнисковому пути), а от них - в кору - в сенсорные зоны, расположенные в постцентральной извилине (в области проекции лица).
Афферентные связи вестибулярного аппарата. Г - глаз; М - мышца; Ж - желудок; ТК - тонкая кишка; ПМ - продолговатый мозг; CM- спинной мозг.
Висцеральный анализатор. интерорецепторы
Рецепторы, расположенные в органах, называются висцерорецепторами, или интерорецепторами.
Импульсация от интерорецепторов поступает в продолговатый мозг по волокнам IX (языкоглоточного) и Х (блуждающего) нервов, проходя через чувствительные ганглии - верхний и нижний ганглии языкоглоточного нерва, верхний и нижний ганглии блуждающего нерва. Затем она достигает ядра одиночного нерва (ядро солитарного тракта или висцеро-сенсорнос ядро), расположенного в продолговатом мозге. Отсюда начинается путь, идущий через вентробазальное (специфическое) ядро таламуса к коре, лимбической системе. В продолговатом и в среднем мозге часть информации используется для процессов регуляции деятельности органов. Часть импульсов от висцерорецепторов поступает в ретикулярную формацию, от нее - в неспецифические ядра таламуса, затем - диффузно к нейронам коры и лимбической системы. Поэтому при нарушении деятельности внутренних органов у человека возникают неосознанные эмоциональные состояния негативной окраски, например, "беспричинный страх" и т.п. И.М. Сеченов, называя это темным чувством, придавал большое значение потоку импульсов от рецепторов внутренних органов в определении настроения человека, его поступков, действии.
Рассмотрим отдельные виды висцерорецепторов.
Сердечно-сосудистая система. В сердце имеются механорецепторы, реагирующие на растяжение - в эндокарде, эпикарде, миокарде. Кроме этого имеются хеморецепторы, которые возбуждаются при недостатке кислорода или избытке углекислого газа (соответственно - гипоксемия, гиперкапния) и при избытке водородных ионов (ацидоз).
Легкие. В легких имеются три вида мехапорецепторов. В регуляции деятельности системы внешнего дыхания принимают участие и хеморсцспторы сосудистых областей.
Механорецепторы легких - это:
1) рецепторы растяжения,
2) ирритантные рецепторы и 3) рецепторы типа J- юкстаальвеолярные рецепторы капилляров.
Рецепторы растяжения возбуждаются во время глубокого вдоха.
Ирритантные рецепторы расположены в эпителиальном и субэпителиальном слоях всех воздухоносных путей. Особенно их много в области корней легких. Они не являются "чистыми" мехацоренепторами: частично реагируют на пары едких веществ - аммиака, эфира, табачного дыма, двуокиси серы, а также на химические вещества типа гистамина. Ирритантные рецепторы возбуждаются при быстром вдохе и быстром выдохе, наличии во вдыхаемом воздухе частичек пыли, содержании в воздухоносных путях слизи, содержании во вдыхаемом воздухе паров едких веществ, ряда химических веществ. Это возбуждение порождает явление одышки - частое и поверхностное дыхание, а также остановку дыхания, например, при наличии паров аммиака, кашель. Их возбуждение вызывает неприятные ощущения першения и жжения.
Рецепторы типа J - или юкстаальвеолярные рецепторы капилляров - находятся вблизи (юкста) капилляров малого круга кровообращения в интерстициальной ткани альвеол. Они возбуждаются в ответ на выделение ряда БАВ, в ответ на отек ткани и вызывают одышку.
Почки. Кровь. Для поддержания основных констант организма (гомеостаза) требуются непосредственно органы-исполнители и рецепторы, улавливающие гомеостатические показатели. Об этих рецепторах известно мало.
А) Осморецспторы. Они расположены во многих тканях и органах и чувствительны к изменению осмотического давления внутренней среды организма, являются разновидностью механорецепторов.
Б) Волюморецепторы: они предназначены для оценки объема жидкости, циркулирующей и находящейся и органе.
В) В последние годы подтверждено существование натриорецепторов - они реагируют на изменение уровня натрия в крови - и глюкозорецепторов, реагирующих на изменение уровня глюкозы в крови.
Спиральный ганглий располагается в канале Розенталя, который находится в модиолусе улитки. Он содержит тела афферентных нейронов, дендриты, которых возбуждаются под действием нейромедиаторов, секретируемых клетками кортиева органа, и аксоны, которые направляются в улитковые ядра расположенные в стволе головного мозга. Большинство (приблизительно 95%) афферентных волокон являются толстыми, покрыты миелиновой оболочкой и берут начало от нейронов I типа.
Эти волокна иннервируют исключительно внутренние волосковые клетки. Оставшиеся афферентные волокна тонкие, не имеют миелиновой оболочки и исходят из нейронов II типа, эти волокна контактируют с наружными волосковыми клетками. Около десятка нейронов I типа иннервируют каждую внутреннюю волосковую клетку (сходящаяся иннервационная модель), в то время как афферентные нервные волокна II типа разделяются на несколько ветвей и контактируют с несколькими наружными волосковыми клетками (расходящаяся иннервационная модель).
Вся слуховая информация переносится к стволу мозга по афферентной системе. Слуховой и вестибулярный нервы соединяются друг с другом в преддверно-улитковом нерве (VIII черепной нерв).
Эфферентные волокна берут начало в стволе головного мозга от нейронов, расположенных в комплексе верхней оливы и посылают информацию к улитке посредством синаптических соединений с наружными волосковыми клетками, а также с афферентными волокнами внутренних волосковых клеток. Эфферентная система позволяет центральной нервной системе регулировать работу улитки.
Иннервационная структура Кортиева органа отражает функциональное различие двух типов улитковых волосковых клеток.
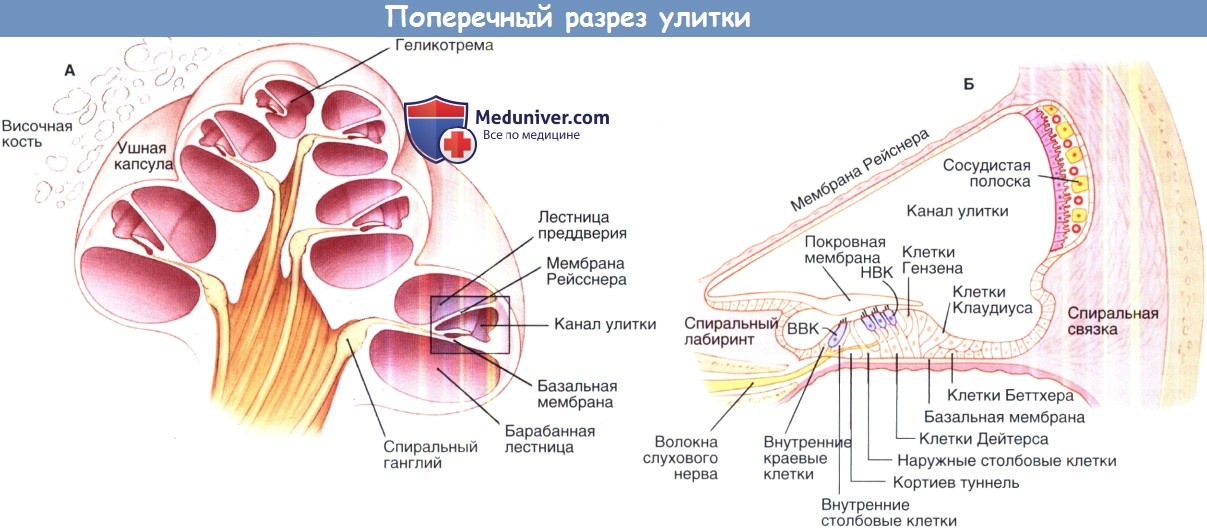
Поперечный разрез улитки. А, схематичное изображение улитки в разрезе.
Выделенная область, показанная на рисунке Б, изображает улитковый проток и окружающие структуры, такие как кортиев орган и сосудистую полоску в разрезе.
ВВК — внутренние волосковые клетки; НВК — наружные волосковые клетки.
а) Нейропроцессинг слуховой информации и синапсы внутренних волосковых клеток. Афферентная нейропередача, осуществляемая внутренними волосковыми клетками, инициируется синапсами ленточного типа количеством от 5 до 30, где поток ионов кальция через потенциал-зависимые кальциевые каналы приводит к четко дозированному высвобождению синаптических пузырьков в пресинаптическую область. В результате экзоцитоз нейротрансмиттеров прямо пропорционален пресинаптическому току кальция, который в свою очередь зависит от изменения потенциала, обусловленного механотрансдукцией.
Информация, кодируемая на афферентных синапсах, поразительно точна, что позволяет получить высокую временную точность, так же как и значительный динамический диапазон с пятикратной амплитудой от 0 до более 100 дБ.
Каждый ленточный синапс состоит из пресинаптического плотного тела (схожего с ленточным синапсом фоторецепторных клеток) в окружении пузырьков с нейротрансмиттерами, плотной полоски пресинаптической мембраны, синаптической щели и постсинаптической области, содержащей АМРА-глютаминэргические рецепторы афферентных нейронов. Считается что глутамат или вещество, близкое по составу, является нейротрансмиттером афферентных связей внутренних волосковых клеток, хотя в этом процессе могут также быть задействованы другие, еще нераспознанные трансмиттеры.
Тонотопическая организация кортиева органа происходит благодаря афферентной системе, где деполяризация внутренних волосковых клеток в определенных участках ведет к возбуждению афферентных нейронов спирального ганглия. Каждое афферентное волокно характеризуется определенной настроечной кривой, описывающей необходимый для получения ответа на заданной частоте уровень звукового давления. Особенность настроечных кривых заключается в отражении частоты наивысшей чувствительности нервного волокна, т.е. частотной характеристики.
Стимулы выше или ниже характерной частоты могут также вызывать ответ, но только при наличии более высокой интенсивности. Форма и пороги чувствительности настроечных кривых афферентных нервных волокон зависят от множества факторов, включая морфологию кортиева органа и активные процессы, связанные с улитковой амплификацией. Активность эфферентной системы играет важную роль в модуляции характеристик афферентных нервов. Потеря улитковой амплификации, в частности, в результате утраты наружных волосковых клеток, ведет к расширению настроечных кривых и увеличению порогов ответа нервного волокна.
Тонотопическая организация улитки является основой частотной кодировки в волокнах слухового нерва (кодирование позиции). Кроме того, выходные характеристики частот кодируются волокнами слухового нерва, это явление известно, как фазовая синхронизация. Волокна слухового нерва посылают сигнал в определенной фазе с частотным стимулом, приводящим к регулярному ответу с характерными интервалами между пиками, равными или кратными длине волны стимула. Фазовая синхронизация возникает только на низких частотах.
Свыше 5 кГц возбуждение, идущее от волокон слухового нерва, возникает с хаотическими интервалами. Тонотопическая организация и фазовая синхронизация крайне важны для выделения сигнала нужной частоты. Выходная частота волокон слухового нерва определяется не только частотными характеристиками, но также и интенсивностью стимулов. По мере увеличения интенсивности стимула увеличивается импульсная частота слухового нерва. Аналогичным образом количество волокон слуховых нервов, активированных на заданных частотах, увеличивается по мере усиления стимула.
Это является результатом вовлечения в процесс большего количества нервных волокон в результате того, что волокна слухового нерва при одинаковых частотных характеристиках имеют разные пороги частотного ответа. Кроме того, при росте интенсивности стимула активируются и другие афферентные нервные волокна с похожими частотными характеристиками. Эти важные физиологические характеристики определяют ряд правил, обязательных при изготовлении и применении имплантатов улитки.
Таким образом, частотно-зависимая стимуляция волосковых клеток ведет к увеличению синаптической передачи в ленточных синапсах внутренних волосковых клеток. Последующее возбуждение постсинаптических токов в волокнах афферентного нерва ведет к временной разрядке потенциала действия в слуховом нерве. В действительности, временное разрешение и динамический диапазон слуховой системы не имеют равных среди сенсорных систем.
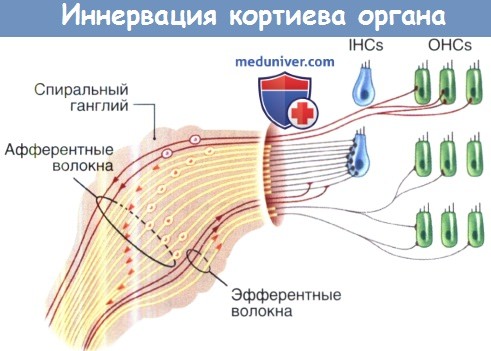
Иннервация кортиева органа.
Схема отображает афферентную и эфферентную иннервацию внутренних и наружных волосковых клеток.
Сверху вниз показаны немиелиновые и миелиновые афферентные волокна II типа,
немиелоновые эфферентные волокна латеральных оливокохлеарных путей (ЛОК) и миелиновые эфферентные волокна медиальных оливокохлеарных путей (МОК).
б) Эфферентная иннервация улитки. В головном мозге четко различают две группы эфферентных нервных волокон. Первые, миелиновые медиальные оливокохлеарные (МОК) эфферентные волокна, берут начало от нейронов, расположенных около верхнего медиального оливного ядра. МОК волокна направляются к контраи ипсилатеральной улитке, где они образуют холинергические синапсы с наружными волосковыми клетками. Вторая группа волокон представлена безмиелиновыми латеральными оливокохлеарными (ЛОК) эфферентными волокнами, которые берут начало от нейронов с маленькими телами, локализующимися около латерального верхнего ядра оливы.
ЛОК волокна направляются преимущественно к ипсилатеральной улитке, где они оканчиваются на дендритах афферентных нейронов I типа у основания внутренних волосковых клеток. Синапсы ЛОК волокон являются сложным химическим комплексом и используют холинэргическую, ГАМК-эргическую и дофаминэргическую передачу, а также различные нейропептиды.
Эффект от стимуляции МОК волокон был изучен подробнее, чем стимуляция ЛОК волокон. В целом стимуляция МОК системы ведет к увеличению порогов чувствительности из-за снижения степени улитковой амплификации наружными волосковыми клетками. Это обусловленная звуком обратная связь, следовательно, чувствительность слухового аппарата снижается в ситуациях, когда нет нужды в энергозатратных механизмах метаболической амплификации.
Функция ЛОК нервных волокон представляется более сложной. Их непосредственный вход в афферентные нейроны позволяет предположить, что они регулируют афферентную активность, тем самым оказывая влияние на динамический диапазон. Исследование различных патологических состояний подтверждает эту точку зрения тем, что снижение специфических нейротрансмиттеров или разрушение тел нейронов в стволе головного мозга ведет либо к усилению, либо к супрессии ответа слухового нерва.
Эффекты обратной связи ЛОК волокон медленны и обычно проявляются в течение минут. Еще одна функция ЛОК системы заключается в обеспечении медленной интеграции и настройки бинауральной функции, необходимой для точного различения локализации звука.
В итоге, активация МОК и ЛОК эфферентной системы, по-видимому, имеет защитный эффект, направленный против акустического повреждения и такая обратная связь может быть важна в шумной окружающей среде.
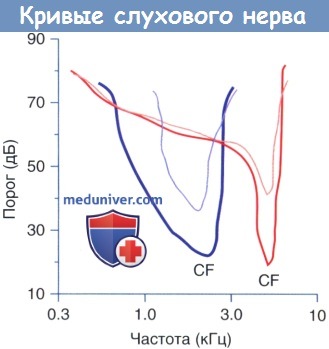
Характерные настроечные кривые слухового нерва (толстая линия).
Настроечная кривая слухового нерва с частотной характеристикой около 2 кГц (синяя) и 5 кГц (красная) (тонкие линии).
Значительное снижение порогов слуховой чувствительности в ответ на стимуляцию МОК системы.
Изменения специфической формы настроечных кривых зависят от частотной характеристики (CF) отдельно взятого волокна.
- Вернуться в оглавление раздела "отоларингология"
ГАНГЛИИ (ganglia нервные узлы)— скопления нервных клеток, окруженные соединительной тканью и клетками глии, расположенные по ходу периферических нервов.
Различают Г. вегетативной и соматической нервной системы. Г. вегетативной нервной системы делятся на симпатические и парасимпатические и содержат тела постганглионарных нейронов. Г. соматической нервной системы представлены спинномозговыми узлами и Г. чувствительных и смешаных черепных нервов, содержащих тела чувствительных нейронов и дающих начало чувствительным порциям спинномозговых и черепных нервов.
Содержание
- 1 Эмбриология
- 2 Анатомия
- 3 Гистология
- 4 Физиология
- 5 Патология
- 6 Краткая анатомическая характеристика нервных ганглиев (узлов)
Эмбриология
Зачатком спинномозговых и вегетативных узлов является ганглиозная пластинка. Она образуется у эмбриона в тех отделах нервной трубки, которые граничат с эктодермой. У зародыша человека на 14—16-й день развития ганглиозная пластинка располагается по дорсальной поверхности замкнувшейся нервной трубки. Затем она расщепляется по всей длине, обе ее половины перемещаются вентрально и в виде нервных валиков лежат между нервной трубкой и поверхностной эктодермой. В дальнейшем соответственно сегментам дорсальной стороны зародыша в нервных валиках возникают очаги пролиферации клеточных элементов; эти участки утолщаются, обособляются и превращаются в спинномозговые узлы. Из ганглиозной пластинки развиваются также чувствительные ганглии У, VII—X пар черепных нервов, аналогичные спинномозговым ганглиям. Зародышевые нервные клетки, нейробласты, формирующие спинномозговые ганглии, представляют собой биполярные клетки, т. е. имеют два отростка, отходящих от противоположных полюсов клетки. Биполярная форма чувствительных нейронов у взрослых млекопитающих и человека сохраняется только в чувствительных клетках преддверно-улит кового нерва, преддверного и спирального ганглиев. В остальных, как спинномозговых, так и черепных чувствительных узлах, отростки биполярных нервных клеток в процессе их роста и развития сближаются и сливаются в большинстве случаев в один общий отросток (processus communis). По этому признаку чувствительные нейроциты (нейроны) называют псевдоуниполярными (neurocytus pseudounipolaris), реже протонейронами, подчеркивая древность их происхождения. Спинномозговые узлы и узлы в. н. с. отличаются характером развития и строения нейронов. Развитие и морфология вегетативных ганглиев — см. Вегетативная нервная система.
Анатомия
Основные сведения об анатомии Г. приведены в таблице.
Гистология


Возраст и перенесенные заболевания сказываются на структуре спинномозговых ганглиев — в них появляется значительно большее, чем у здоровых, количество различных атипичных нейронов, в особенности с дополнительными отростками, снабженными колбовидными утолщениями, как, напр., при ревматическом пороке сердца (рис. 2), стенокардии и др. Клинические наблюдения, а также экспериментальные исследования на животных показали, что чувствительные нейроны спинномозговых узлов реагируют значительно быстрее интенсивным ростом дополнительных отростков на различные эндогенные и экзогенные вредности, нежели моторные соматические или вегетативные нейроны. Эта способность чувствительных нейронов иногда выражена значительно. В случаях хрон, раздражений вновь образованные отростки могут обвиваться (в виде намотки) вокруг тела собственного или соседнего нейрона, напоминая кокон. Чувствительные нейроны спинномозговых узлов, как и другие типы нервных клеток, имеют ядро, различные органеллы и включения в цитоплазме (см. Нервная клетка). Т. о., отличительным свойством чувствительных нейронов спинномозговых и узлов черепных нервов является их яркая морфол, реактивность, выражающаяся в изменчивости их структурных компонентов. Это обеспечено высоким уровнем синтеза белков и различных активных веществ и свидетельствует об их функциональной подвижности.
Физиология
У беспозвоночных Г. играют ту же роль, что и ц. н. с. у позвоночных, являясь высшими центрами координации соматических и вегетативных функций. В эволюционном ряду от червей до головоногих моллюсков и членистоногих Г., перерабатывающие всю информацию о состоянии окружающей и внутренней среды, достигают высокой степени организации. Это обстоятельство, а также простота анатомической препаровки, относительно большие размеры тел нервных клеток, возможность введения в сому нейронов под прямым визуальным контролем одновременно нескольких микроэлектродов сделали Г. беспозвоночных общераспространенным объектом нейрофизиол, экспериментов. На нейронах круглых червей, октапод, декапод, брюхоногих и головоногих моллюсков методами электрофореза, прямого измерения активности ионов и фиксации напряжения проводят исследования механизмов генерации потенциалов и процесса синаптической передачи возбуждения и торможения, зачастую невыполнимые на большинстве нейронов млекопитающих. Несмотря на эволюционные различия, основные электрофизиол, константы и нейрофизиол, механизмы работы нейронов во многом одинаковы у беспозвоночных и высших позвоночных животных. Поэтому исследования Г., беспозвоночных имеют общефизиол. значение.
У позвоночных соматосенсорные черепные и спинномозговые Г. функционально однотипны. Они содержат тела и проксимальные части отростков афферентных нейронов, передающих импульсацию от периферических рецепторов в ц. н. с. В сомато-сенсорных Г. нет синаптических переключений, эфферентных нейронов и волокон. Так, нейроны спинномозговых Г. у жабы характеризуются следующими основными электрофизиол, параметрами: специфическим сопротивлением — 2,25 кОм/см 2 для деполяризующего и 4,03 кОм/см 2 для гиперполяризующего тока и удельной емкостью 1,07 мкФ/см 2 . Полное входное сопротивление нейронов сомато-сенсорных Г. существенно ниже, чем соответствующий параметр аксонов, поэтому при высокочастотной афферентной импульсации (до 100 импульсов в 1 сек.) проведение возбуждения может блокироваться на уровне тела клетки. В этом случае потенциалы действия, хотя и не регистрируются от тела клетки, продолжают проводиться с периферического нерва в задний корешок и сохраняются даже после экстирпации тел нервных клеток при условии интактности Т-образных ветвлений аксонов. Следовательно, возбуждение сомы нейронов сомато-сенсорных Г. для передачи импульсов от периферических рецепторов к спинному мозгу не обязательно. Эта особенность впервые появляется в эволюционном ряду у бесхвостых амфибий.
Вегетативные Г. позвоночных в функциональном плане принято разделять на симпатические и парасимпатические. Во всех вегетативных Г. происходит синаптическое переключение с преганглионарных волокон на постганглионарные нейроны. В подавляющем большинстве случаев синаптическая передача осуществляется хим. путем с помощью ацетилхолина (см. Медиаторы). В парасимпатическом цилиарном Г. птиц обнаружена электрическая передача импульсов при помощи так наз. потенциалов соединения, или потенциалов связи. Электрическая передача возбуждения через один и тот же синапс возможна в двух направлениях; в процессе онтогенеза она формируется позже химической. Функциональное значение электрической передачи пока не ясно. В симпатических Г. амфибий выявлено небольшое количество синапсов с хим. передачей нехолинергической природы. В ответ на сильное одиночное раздражение преганглионарных волокон симпатического Г. в постганглионарном нерве прежде всего возникает ранняя отрицательная волна (O-волна), обусловленная возбуждающими постсинаптическими потенциалами (ВПСП) при активации н-холинорецепторов постганглионарных нейронов. Тормозной постсинаптический потенциал (ТПСП), возникающий в постганглионарных нейронах под действием катехоламинов, выделяемых хромаффинными клетками в ответ на активацию их м-холинорецепторов, формирует следующую за 0-волной положительную волну (П-волна). Поздняя отрицательная волна (ПО-волна) отражает ВПСП постганглионарных нейронов при активации их м-холинорецепторов. Завершает процесс длительная поздняя отрицательная волна (ДПО-волна), возникающая вследствие суммации ВПСП нехолинергической природы в постганглионарных нейронах. В нормальных условиях на высоте О-волны при достижении ВПСП величины 8—25 мв возникает распространяющийся потенциал возбуждения с амплитудой 55—96 мв, длительностью 1,5—3,0 мсек, сопровождаемый волной следовой гиперполяризации. Последняя существенно маскирует волны П и ПО. На высоте следовой гиперполяризации возбудимость снижается (период рефрактерности), поэтому обычно частота разрядов постганглионарных нейронов не превышает 20—30 импульсов в 1 сек. По основным электрофизиол. характеристикам нейроны вегетативных Г. идентичны большинству нейронов ц. н. с. Нейрофизиол. особенностью нейронов вегетативных Г. является отсутствие истинной спонтанной активности при деафферентации. Среди пре- и постганглионарных нейронов преобладают нейроны групп В и С по классификации Гассера — Эрлангера, основанной на электрофизиол, характеристиках нервных волокон (см.). Преганглионарные волокна обширно ветвятся, поэтому раздражение одной преганглионарной ветви приводит к возникновению ВПСП во многих нейронах нескольких Г. (феномен мультипликации). В свою очередь на каждом постганглионарном нейроне оканчиваются терминали многих преганглионарных нейронов, отличающихся по порогу раздражения и скорости проведения (феномен конвергенции). Условно мерой конвергенции можно считать отношение количества постганглионарных нейронов к количеству преганглионарных нервных волокон. Во всех вегетативных Г. оно больше единицы (за исключением цилиарного ганглия птиц). В эволюционном ряду это отношение возрастает, достигая в симпатических Г. человека величины 100:1. Мультипликация и конвергенция, обеспечивающие пространственную суммации) нервных импульсов, в сочетании с временной суммацией являются основой интегрирующей функции Г. при обработке центробежной и периферической импульсации. Через все вегетативные Г. проходят афферентные пути, тела нейронов которых лежат в спинномозговых Г. Для нижнего брыжеечного Г., чревного сплетения и некоторых интрамуральных парасимпатических Г. доказано существование истинных периферических рефлексов. Афферентные волокна, проводящие возбуждение с малой скоростью (ок. 0,3 м/сек), входят в Г. в составе постганглионарных нервов и оканчиваются на постганглионарных нейронах. В вегетативных Г. обнаружены окончания афферентных волокон. Последние информируют ц. н. с. о происходящих в Г. функционально-хим. изменениях.
Патология
В клин, практике наиболее часто встречается ганглионит (см.), называемый также симпато-ганглионитом, — заболевание, связанное с поражением ганглиев симпатического ствола. Поражение нескольких узлов определяют как полиганглионит, или трунцит (см.).
Спинальные ганглии нередко вовлекаются в патологический процесс при радикулитах (см.).
Читайте также:
