
Что такое нервный субстрат
Только широкое применение реакции самораздражения, о которой мы писали выше, позволило его последователям приступить к изучению истинно эмоциональных пунктов мозга, раздражение которых приводило к возникновению у животных эмоциональных реакций. Многочисленные эксперименты, проведенные в этом направлении, показали, что эмоциональные аппараты тесно связаны с деятельностью лимбических образований мозга, таких, как гипоталамус, гиппокамп, перегородка, миндалина, поясная извилина, височная кора и т. д. Именно при нахождении раздражающих электродов в этих образованиях мозга животные наиболее отчетливо проявляют реакции самораздражения.
Каким же образом объединяются различные отделы головного мозга в построении эмоциональных реакций?
Многочисленные исследования, в том числе и работы нашей лаборатории, показывают, что, возникая в гипоталамических и других лимбических образованиях мозга, эмоциональное возбуждение не ограничивается только этими структурами, а в форме восходящих активирующих влияний начинает широко распространяться на другие отделы мозга, вплоть до коры больших полушарий. Такую реакцию можно отчетливо наблюдать на электроэнцефалограмме в виде генерализованной реакции десинхронизации электрической активности при раздражении любого эмоционального центра гипоталамуса. Кроме того, при исследовании электрической активности отдельных нейронов в различных областях коры мозга можно обнаружить их участие в возбуждениях, вызванных раздражением тех пунктов гипоталамической области, которые вызывают у животных поведенческие реакции самораздражения или избегания (В. И. Бадиков, 1969).
Все эти наблюдения показывают, что эмоциональное возбуждение действительно является глобальной реакцией целого мозга, объединяющей различные корково-подкорковые структуры в интегрированный комплекс. Этот комплекс, формирующий потребности, может возбуждаться как внутренними, так и внешними факторами.
Первоочередная задача современной нейрофизиологии — выяснение механизмов формирования таких комплексов эмоциональных возбуждений. Особый интерес представляют наблюдения, показывающие, что положительные и отрицательные эмоциональные возбуждения могут адресоваться к одним и тем же нейронам коры больших полушарий. В связи с этим возникает следующая задача изучения эмоций — исследование особенностей химических механизмов, с помощью которых на одних и тех же нейронах коры больших полушарий реализуются эмоциональные реакции различного биологического качества.
Другой важной особенностью формирования эмоциональных возбуждений в центральной нервной системе является то, что внутри головного мозга, особенно в лимбических образованиях — главном субстрате эмоций имеется множество замкнутых кольцевых путей, по которым эмоциональный разряд может длительное время циркулировать.
Каковы же пути перехода отрицательной эмоции в застойное состояние?
Одной из первостепенных причин этого является невротическое состояние. Именно невроз становится поставщиком самых разнообразных заболеваний — гипертонии, экземы, спастических состояний и т. д.
Нервная система. Вопросы по нервной системе. Экспресс контроль лекции по теме: Введение в неврологию.. Строение ЦНС, ПНС, нейроны, синапсы…
1. Функции нервной системы
1) Регуляции всех функций организма, а также обеспечивает целостность организма, интеграцию организма (взаимосвязь всех органов и систем).
2) Координация, согласует функции всех органов и систем, связь организма с внешней средой. В процессе эволюции нервная система в первую очередь возникла для связи с внешней средой.
3) Кора головного мозга является основой мышления. У животных образное мышление, у человека мысли в речевой оболочке.
4) Память – хранение информации.
2. Основные этапы эволюции нервной системы
Сначала — гуморальная регуляция — это способность некоторых клеток воспринимать раздражение и проводить импульсы. Затем:
• Сетевидная (диффузная) нервная система (гидра).
• Узловая нервная система. Нервные клетки стали концентрироваться и специализироваться, следовательно, начинается образование нервных узлов и нервов.
• Трубчатая нервная система (хордовые).
• Цефализация – появление головного мозга. Впервые — у низших рыб.
• Кортикализация – на поверхности полушарий большого мозга образуется кора.
Отличие человеческого мозга — речевые центры (сенсорный и моторный), развитие логического мышления. Лобные доли отвечают за развитие интеллекта.
3. Какие факторы обусловили формирование трубчатой нервной системы, цефализацию и кортикализацию?
• Трубчатая нервная система (хордовые). Возникла из-за усложнения двигательной активности.
• Цефализация – появление головного мозга. Впервые — у низших рыб (из-за формирования лидирующего переднего конца, там — органы чувств, это привело к усиленному развитию и появлению головного мозга).
• Кортикализация – на поверхности полушарий большого мозга образуется кора из-за изменения среды обитания (земноводные). У птиц меньше в сравнении с рептилиями.
4. По каким причинам и как осуществляется классификация нервной системы.
По топографии:
• ЦНС – находятся нервные центры.
• ПНС – 31 пара спинномозговых нервов + 12 пар черепных нервов (связь ЦНС с организмом).
По функции:
• соматическая (сознательная) — регуляция функций скелетной мускулатуры
• вегетативная (бессознательная) — регуляция функций внутренних органов, желез, ССС.
СНС и ВНС имеют:
— центры в головном мозге
— нервы в составе черепных нервов
— нервы в составе спинномозговых нервов.
5. Что такое нейрон? Его строение.
Нервная система состоит из нервной ткани. Ткань образуется нервными клетками – нейроны и нейроглии.
Нейрон – структурно-функциональная единица нервной системы.
Составляет основу строения нервной системы и обеспечивает возбуждение и проведение.
Нейрон имеет:
• тело (нейролемма, нейроплазма, специфические органоиды). Содержит темный пигмент – меланин (нейроплазма) серого цвета.
а) Дендриты – древовидно ветвящиеся. Их может быть много. Импульс проводит к телу (центростремительно).
б) Аксон – осевой отросток. Есть только конечное ответвление. Импульс проводит от тела. (центробежно).
Отростки заключены в миелиновую оболочку белого цвета (продукт нейроглий).
6. Классификация нейронов по строению.
1) Одноотросчатые (униполярные) – от тела один отросток: палочки и колбочки сетчатки.
2) Двуотросчатые (биполярные) – в сетчатке.
3) Ложные одноотросчатые (псевдоуниполярные) – один отросток делится на дендрит и аксон. Чувствительные узлы спинномозговых и черепных нервов.
4) Многоотросчатые (мультиполярные).
5) Безотросчатые – стволовые нервные клетки эмбриона.
7. Классификация нейронов по функциям.
1) Чувствительные нейроны (афферентные).
- псевдоуниполярные,
- тела — в чувствительных узлах спинномозговых и черепных нервов,
- дендриты на периферии — заканчиваются рецепторами (восприятие раздражения и преобразование в импульс),
- дендриты проводят импульс центростремительно.
2) Двигательные нейроны (эфферентные).
- мультиполярные,
- тела — в двигательных ядрах спинномозговых и черепных нервов,
- аксоны заканчиваются в мышцах,
- аксон проводит импульс, происходит сокращение мышцы.
3) Вставочные нейроны (ассоциативные).
- мультиполярные,
- тела — в ядрах спинного мозга, ствола конечного мозга, коре,
- обеспечение связи двух нейронов, тела вставочных нейронов
- образуют нервные центры (кроме двигательных ядер)
4) Нейросекреторные нейроны – выработка гормонов и регуляция всех функций организма.
8. Узлы, ядра, кора: их сходство и отличия.
Скопление тел имеет три разновидности: узлы, ядра, кора.
Они отличаются по локализации:
- Узлы – скопление тел на периферии в составе ПНС (за пределами ЦНС).
- Ядра – скопление тел внутри головного и спинного мозга.
- Кора – скопление тел на поверхности полушарий.
- чувствительные,
- вегетативные.
- чувствительные,
- вегетативные,
- двигательные.
- чувствительные зоны,
- двигательные зоны,
- ассоциативные поля.
9. Что такое нервное волокно. Как образуются нервы и проводящие пути, их назначение.
Скопление отростков образует белое вещество. Существует в виде проводящих путей и нервов.
Проводящие пути – скопление отростков внутри спинного и головного мозга. Связывают различные нервные центры друг с другом. Бывают чувствительными и двигательными.
Нервы – скопление отростков на периферии вне спинного и головного мозга.
Связывают нервные центры со всем организмом. По составу волокон нервы: двигательные, чувствительные, смешанные.
Нервные волокна — это скопление отростков нервных клеток, которые окружены оболочкой из олигодендроцитов (клетки Шванна).
10. На какие делятся по составу волокон нервы и проводящие пути.
• Нисходящие – двигательные нервы:
11. Что такое синапс? Его разновидности.
Синапсы — места контактов нейронов.
Виды (морфологические + функциональные контакты):
- Аксосоматические,
- Аксодендритические,
- Аксоаксиальные,
- Дендродендритические.
12. Что такое рефлекс? Что является его морфологическим субстратом?
Основа деятельности нервной системы — рефлекс. Это ответная реакция организма на раздражение.
Виды ответной реакции:
Морфологический субстрат рефлексов — рефлекторная дуга. Это цепь нейронов, контактирующих друг с другом в области синапсов.
По количеству нейронов дуги:
• Простые – два или три нейрона,
• Сложные – из большого количества.
13. Начертите схему 3-х нейронной рефлекторной дуги. Чем отличается от рефлекторной дуги рефлекторное кольцо?
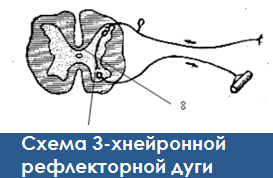
В любой рефлекторной дуге есть обратная связь – образуется рефлекторное кольцо, это обеспечивает анализ полученных данных.
Физиологический субстрат эмоций
В эмоциональном реагировании участвует не только психика, но и тело. Эмоциональные состояния сопровождаются физиологическим возбуждением, которое, если оно заметно, указывает на эмоциональную вовлеченность в произошедшее событие. За последние годы были сделаны важные открытия в области изучения физиологических механизмов эмоций, а также индивидуальных различий в их физиологии. В большинстве работ делается попытка выявить определенные физиологические механизмы и процессы, связанные с разными эмоциональными состояниями.
В определенном смысле заявление о том, что разные эмоции опосредуются разными физиологическими системами, банально. Эмоциональные реакции, как и другие физиологические феномены, являются продуктом активности нервной системы. Разные эмоциональные состояния с неизбежностью должны быть связаны с разными видами физиологической активации. Трудность состоит в том, чтобы определить сущность и функции физиологических систем при разных эмоциях и выявить индивидуальные различия в физиологии, обусловливающие различия в эмоциональных переживаниях.
В процессе поиска эмоционально-специфичных физиологических механизмов исследуются разнообразные физиологические системы, в том числе автономная нервная система и ее влияние на периферическую физиологию; подкорковые структуры центральной нервной системы, особенно лимбическая система; и корковая активность мозга.
Эмоции связаны с возбуждением. Мы живо ощущаем изменения в частоте сердечных сокращений, артериальном давлении и активности потовых желез, свидетельствующие о нашем эмоциональном возбуждении. Эти изменения в периферической физиологии связаны с деятельностью автономной нервной системы. Симпатическая ветвь автономной нервной системы подготавливает организм к энергичному поведению, активизируя физиологические механизмы, обеспечивающие потенциальные действия (Levenson, 1992). Разные эмоции предрасполагают к разным видам действия, поэтому паттерны возбуждения также эмоционально специфичны. Поскольку гнев, к примеру, предрасполагает к нападению, он связан с периферическим возбуждением, которое специфически обеспечивает агрессивное, атакующее поведение. Поэтому один из потенциальных источников эмоционально-специфической физиологии — это эмоционально специфические паттерны возбуждения в физиологических механизмах, которыми управляет автономная нервная система.
Данные о том, что разным эмоциям сопутствуют специфические паттерны автономного возбуждения, были получены главным образом при использовании двух экспериментальных парадигм (Levenson, 1992). Например, Экман с соавторами (Ekman et al, 1983) просили профессиональных актеров изобразить на своем лице гнев, страх, грусть, радость, отвращение и удивление. Во втором задании актеры должны были оживить в памяти последний эпизод, в котором они испытали каждую из перечисленных эмоций. При выполнении этих заданий были зафиксированы значимые надежные различия в активации автономной нервной системы. Изображение гнева и страха повышало частоту сердечных сокращений по сравнению с радостью, грустью и отвращением. Это и понятно, учитывая тот факт, что реакция борьбы при гневе и реакция бегства при страхе требуют мобилизации энергии, для которой, в свою очередь, необходимо повышение уровня сердечных сокращений. Хотя при гневе и страхе уровень сердечных сокращений сходен, эти эмоции различаются по другим показателям деятельности автономной нервной системы. Температура кожи (на пальцах) при гневе оказалась выше, чем при страхе, что объясняется направлением крови при страхе от периферии к крупной мускулатуре для обеспечения передвижения, необходимого для избегания угрожающей ситуации. Поэтому температура кожи на периферии не повышается, несмотря на повышение частоты сердечных сокращений (Levenson, 1992). Эти результаты были воспроизведены и с испытуемыми не актерами (Levenson, Ekman, & Friesen, 1990), а также в одной из незападных культур — с народностью минангкабау (Западная Суматра) (Levenson, 1992). Однако следует отметить, что различия в деятельности автономной нервной системы при выполнении заданий на изображение мимики оказались более достоверными, чем при выполнении задания на оживление в памяти соответствующих эпизодов (Ekman et al, 1983). Наряду с гневом и страхом грусть также оказалась связанной с повышением частоты сердечных сокращений (Ekman et al, 1983; Schwartz et al, 1981); это свидетельствует о том, что грусть также стрессовое состояние с высоким уровнем возбуждения. И напротив, такая негативная эмоция, как отвращение оказалась связанной с более низкой частотой сердечных сокращений (Levenson, 1992).
Хотя некоторые эмоциональные состояния можно дифференцировать по периферическим физиологическим реакциям, не все эмоции можно определить таким способом. В действительности результаты, полученные при регистрации деятельности автономной нервной системы, не особенно убедительны. Очевидно, что уровень активации автономной нервной системы не может полностью определять эмоциональный опыт. В действительности, поскольку автономная нервная система отвечает за обеспечение организма энергией, а энергетические расходы на разные эмоции могут быть одинаковыми, паттерны возбуждения автономной нервной системы, вероятно, не лучший показатель, по которому можно судить об эмоционально-специфической физиологии (Gray, 1994). Более информативны в отношении биологических механизмов, обеспечивающих разные эмоции, данные о функционировании центральной нервной системы.

Кафедра физиологии и психофизиологии
Нервные механизмы эмоционального и мотивированного поведения
Мурик Сергей Эдуардович
В настоящее время становится всё более очевидно, что в нейропсихологии одним из наименее разработанных вопросов остаётся проблема механизма эмоций. Несмотря на кажущуюся простоту явления, наличие у эмоций специфических психофизиологических черт и выделения их давно в самостоятельный психический процесс- нейрофизиологический субстрат эмоций остается, очевидно, также мало понятен сегодня, как и 50 и 100 лет назад. Со времен Джеймса и Ланге мало что изменилось в понимании этого вопроса. В настоящее время вряд ли кто осмелится безаппелляционно утверждать, что мы сегодня знаем больше, чем они знали в своё время (конец 19 века) о том, где в нервной системе рождаются эмоции и каков механизм субъективного переживания мозгом объективной реальности.
На наш взгляд, главной причиной, тормозящей разработку нейрофизиологического механизма эмоций, является отрыв их нервного субстрата от сенсорных процессов. До сих пор разработка этого вопроса шла в русле поиска специфического центра эмоций. Начиная с работ Кеннона ( P. Cannon) и Барда (P.Bard) и заканчивая современными работами (П.В.Симонов, 1966-1999), центр эмоций перемещали из одной структуры в другую (таламус, гипоталамус) иногда, отводя под него системы структур (круг Пейпеца, лимбический мозг), но всегда эмоции пытались локализовать, найти своеобразный эмоциональный “центрэнцефалон”. Поиски в этом направлении так ни к чему и не привели. Более того, мы можем с большой долей уверенности, в свою очередь, утверждать, что такой центр никогда и не будет найден, поскольку этот путь- путь поиска “седалища” эмоций порочен по своей сути и поэтому бесперспективен.
С точки зрения нервного субстрата, эмоции, как субъективное переживание биологической значимости раздражителей из внешней и внутренней среды организма, на наш взгляд, не существуют, как самостоятельное явление. Они неразрывно связаны с сенсорными процессами, сопровождая их, и локализуются соответственно в сенсорных системах.
Если вопрос о “субстрате эмоций” при таком подходе снимается, то нейрофизиологический механизм эмоций остается не очевидным. Например, непонятно, в чем специфика нервных процессов в сенсорных системах, когда при анализе раздражителей мы не испытываем эмоции или, когда действие раздражителя субъективно переживается в виде негативной или положительной эмоции.
Ответы на эти вопросы мы пытаемся найти, развивая поляризационную теорию эмоций и мотиваций (Мурик, 1994-1999), согласно которой нейрофизиологическая основа эмоций и мотиваций тесно слита и связана с сенсорными процессами, являясь своеобразной формой отражения биологической значимости раздражителей ввиде изменения возбудимости нейронов, участвующих в этих процессах. Приэтом, снижение возбудимости нейронов при относительно длительной деполяризации их мембраны под действием стимулов из внешней или внутренней среды субъективно переживается ввиде негативной эмоции. Обратные процессы, связанные с восстановлением мембранного потенциала клеток и гиперполяризацией, и повышение при этом их возбудимости, субъективно оцениваются ввиде положительных эмоций.
Деполяризационные процессы, по-нашему мнению, лежат не только в основе негативных эмоций, но и являются нейрофизиологической основой мотиваций, так как деполяризованное состояние нейронов отражает либо их неблагоприятное функциональное состояние (как при гипоксии), либо наличие повреждающих факторов. В том и другом случае живые системы должны избегать этого состояния, что и лежит в основе мотивированного поведения.
Из развиваемого нами подхода становится очевидным, что нервный субстрат мотиваций и эмоций тесно слит не только с афферентными системами (сюда мы включаем сенсорные и ассоциативные системы), но и между собой. Положительные эмоции, в таком случае, отражают процессы редукции мотивационного (деполяризационного) очага, вплоть до развития гиперполяризованного состояния.
Нами сделаны первые шаги в экспериментальном исследовании роли поляризационных процессов нервной ткани в нейрофизиологическом механизме мотивированного и эмоционального поведения. В частности, мы изучали влияние искусственной поляризации нервной ткани на субъективное состояние животных. Нами было показано, что поляризация анодом постоянного тока зрительной коры крыс уменьшает, а катодом постоянного тока- увеличивает эмоцию страха в открытом поле (Murik S.E. The relation of emotions to polarization proсesses in sensory systems. Intern J. Neuroscience. 8/3-4:185-197. 1997). Действие тока, в тоже время влияет, и на информационные процессы (Murik S.E. Polarization processes in the nervous system and behavior. Intern J. Neuroscience. 94:213-221. 1998).
Изучение роли поляризационных процессов в механизме эмоций и мотиваций, на наш взгляд, возможно, в частности, посредством исследования изменений уровня постоянного потенциала головного мозга. Нами начаты эксперименты в этом направлении и получены первые результаты (Мурик,1999). В частности, показано, что формирование пищевой мотивации и её удовлетворение отражается на уровне постоянного потенциала ряда структур головного мозга. Полученные результаты мы рассматриваем как свидетельство участия поляризационных процессов в механизме пищевого поведения.
Мы были бы благодарны всем, кто примет участие в обсуждении данной проблемы и развиваемого нами подхода к её решению.
Физиологической основой эмоций является прежде всего деятельность подкорковых отделов мозга – таламуса, гипоталамуса, лимбической системы.
Аналогичное явление наблюдается в клинике там, где интенсивность корковой деятельности ослаблена в силу некоторых заболеваний: у больных появляется чрезмерная вспыльчивость, гневливость и прочие подобные явления.
Сложные эмоциональные переживания, чувства невозможны без участия коры головного мозга. И.П.Павловым было показано, что именно кора регулирует протекание и выражение эмоций.
Существенную роль в эмоциональных переживаниях человека играет вторая сигнальная система: переживания могут возникнуть не только при непосредственном воздействии внешней среды, но могут быть вызваны словами, мыслями.
Теории эмоций.
Кроме того, было установлено, что важную роль в регуляции эмоций играет также ретикулярная формация, активность которой определяет многие динамические параметры эмоций – силу, продолжительность, изменчивость. Внешние или внутренние стимулы порождают импульсы, активирующие ствол мозга. Ретикулярная формация, в свою очередь, активирует гипоталамус и кору мозга. В конечном итоге происходит трансформация первичного стимула в поведение, сопровождающееся эмоциональным возбуждением.
Когнитивные теории эмоций.
Теория когнитивного диссонанса Л.Фестингера. Положительная эмоция возникает тогда, когда ожидания человека подтверждаются, т.е. когда реальные результаты деятельности согласуются с ранее намеченным планом (находятся в консонансе).
Отрицательные эмоции возникают в противоположной ситуации, когда ожидаемые и действительные результаты расходятся (находятся в диссонансе).
Информационная теория П.В.Симонова. Степень эмоционального напряжения определяется потребностью и дефицитом информации, необходимой для удовлетворения этой потребности.
Воля.
Воля – это психический процесс сознательного управления и регулирования своего поведения, обеспечивающий преодоление трудностей и препятствий на пути к поставленной цели.
Низшие уровни воли, такие как произвольное движение или задержка импульсивного действия, выделяются уже у животных, однако развитая воля является специфической человеческой функцией.
Воля является социальным новообразованием психики, возникла и формируется в процессе трудовой деятельности человека.
Воля обеспечивает выполнение двух взаимосвязанных функций: побудительной и тормозной.
Побудительная функция воли обеспечивает активизацию действий человека, преодоление препятствий на пути к достижению цели.
Тормозная функция воли заключается в возможностях человека удержаться от совершения каких-либо действий, а если они начались, то замедлить или остановить их. Человек способен затормозить побуждения и выполнение действий, которые не соответствуют его убеждениям, идеалам, мировоззрению. По мнению И.П.Павлова, волевое торможение человеком своей деятельности не менее, а часто более сложное волевое усилие, чем активизация.
В своём единстве тормозная и побудительная функции воли обеспечивают преодоление трудностей на пути к достижению цели, т.е. обеспечивают волевую регуляцию поведения человека.
Говоря о волевой регуляции поведения, обычно упоминают о том, как человек решает вопрос контроля над происходящими в его жизни событиями. Локализация контроля (или локус контроля) - это склонность человека приписывать ответственность за происходящее внешним или внутренним силам.
По этому принципу люди делятся на интерналов и экстерналов.
Интерналы чаще всего чувствуют личную ответственность за свои действия, объясняя их своими личностными особенностями, поэтому они стремятся работать над собой, самосовершенствоваться, в том числе и развивать свою волю.
Экстерналы же всё стараются объяснить внешними обстоятельствами, снимая с себя всяческую ответственность и тем самым снижая актуальность волевой активности и волевой тренировки.
Сомато-вегетативные реакции, наблюдаемые при эмоциональном состоянии, должно быть, интегрируются в виде поведенческих актов особым нервным субстратом, который тесно связан с субстратом эмоционального переживания. Координирующие центры же этих сомато-вегетативных реакций, должно быть, находятся в разных областях промежуточного мозга, главным образом в гипоталамусе. Детальными исследованиями Гесса (Гесс, 21) было показано, что точечным раздражением разных частей гипоталамуса кошки можно вызвать движение волос, расширение или сужение зрачка, дефекацию, одышку, обнюхивательные движения, засыпание, атонию, аффективную защитную реакцию, опускание головы и передней части тела, поднятие головы и передней части тела и т. д. Такие разнообразные внешние реакции наблюдались также другими авторами (Каада, 2; Коган, 22). Все эти разнообразные внешние реакции по-разному объединяются в поведенческих актах.
Спрашивается, где же собственно лежит тот нервный субстрат, который продуцирует эмоциональные переживания и интегрирует сомато-вегетативные реакции, т. е. производит всю эмоциональную реакцию с ее субъективными и объективными проявлениями в неокортексе или архипалеокортексе. Есть предположение, что местом эмоциональных переживаний, как и эмоциональных внешних выражений, является гипоталамус — часть промужуточного мозга (Гельгорн, 13; Гесс, 21, и др.). Лешли еще в 1938 г. (8) критически рассмотрел подобные предположения и на основании определенных клинических фактов пришел к выводу, что те центры промежуточного мозга, которые повинны в возникновении эмоциональных внешних выражений, не являются источником кортикопетальных импульсов, которые определяют характер субъективных переживаний. Он же выступает против наличия особых центров в промежуточном мозге для эмоциональных переживаний. Известно, например, что конвульсивный смех или плач, наблюдаемый при разных заболеваниях ствола головного мозга, не сопровождается эмоциональным переживанием.
Такая диссоциация эмоциональных переживаний и выразительных движений лучше всего свидетельствует о том, что обе стороны эмо-
циональной реакции не находятся в неразрывной связи друг с другом, не исходят из одних и тех же таламических или иных центров.
Опыты Массермана (23) еще лучше показали, что эмоциональные эффекты, вызванные у кошек прямым раздражением гипоталамуса, не всегда полноценны, даже если данное раздражение наносится нормальным животным. Эти эмоциональные эффекты не направлены на определенные объекты внешней среды, они прекращаются сейчас же после прекращения раздражения, не бывает последействия (в виде мяукания, дрожания, убегания и т. д.), как это наблюдается при действительных эмоциональных реакциях. Кроме того, когда животное производило нормальные поведенческие акты еды, умывания, чистки, лакания молока, то эти эмоционально выраженные поведенческие акты не менялись при раздражениях гипоталамуса, если только не наступали очень сильные двигательные эффекты, которые прерывали их.
Опыты на животных с удалением неокортекса ясно показывают, что такие животные не лишены полноценных эмоциональных реакций. Так, например, по опытам Орджоникидзе и Нуцубидзе (24, 25), кошки после оперативного удаления неокортекса обнаруживают чувство голода и насыщения. В голодном состоянии они становятся беспокойными, мяукают и даже неистово кричат. При насыщении они успокаиваются, перестают мяукать. Эти кошки мясо предпочитают хлебу. Когда предлагают им мясо и хлеб одновременно, они, как правило, поедают сначала мясо. Если дать им кусок воска "и хлеба, захватывают только хлеб. Если прилепить воск к хлебу, они съедают хлеб и не берут воск в рот.
Если исходить из приведенных фактов, нужно полагать, что нервные комплексы, производящие субъективные эмоциональные переживания и интегративные внешние эмоциональные реакции, должны находиться в ростральной части головного мозга, а именно в палеокортексе и еще в подкорковом образовании — в миндалевидных ядрах. Эти последние ядра анатомически тесно связаны с важнейшими отделами архипалеокортекса, как грушевидная извилина. Кроме того, известно, что миндалевидные ядра, подобно грушевидной извилине и гиппокампу, получают афферентную импульсацию от рецепторов главным образом через неспецифическую систему (см. Глур, 27).
Но насчет тесной топографической локализации эмоциональных функций в этих ростральных отделах мозга существует большое расхождение. Одни авторы считают, что участки мозга, вызывающие разные моторные, вегетативные и сложно поведенческие реакции, перекрывают друг друга и поэтому не намечается топографическая локализация функции (Глур, 27). Но другие авторы, как Каада и Урсин (28, 29), Брейди (30), Гриин и Ардуини (31), Эди (32) и др., считают, что ростральные отделы так называемой лимбической
системы принимают более или менее дифференцированное участие в образовании сложных эмоциональных форм поведения.
И так, не может быть сомнения, что мотивационно-эмоционалъные функции принадлежат не просто архипалеокортексу, а лимбической системе, куда входят как отделы архипалеокортекса, так и подкорковые образования в виде миндалевидных ядер и гипоталамуса. Ниже будет подробно изложено, какую роль могут играть все эти отделы головного мозга в происхождении эмоционального поведения.
Последнее изменение этой страницы: 2017-02-21; Нарушение авторского права страницы
Читайте также:
