
Этапы проведения нервного импульса
Возникновение нервного импульса связано с состоянием мембраны нейрона. Биологические мембраны, будучи липидно-белковым комплексом, непроницаемы для заряженных ионов. Перемещение ионов через мембрану осуществляется с помощью специальных транспортных систем: натриевого насоса, работающего с затратой энергии (Na + , К + -АТФ-аза) и ионпроводящих каналов – натриевых и калиевых.
В состоянии физиологического покоя по разные стороны мембраны имеет место разность концентраций ионов, главным образом Na + и К + . Концентрация ионов К + выше в клетке, ионов Na + – в межклеточной жидкости. Это влияет и на распределение других ионов. Разность концентраций ионов поддерживается с помощью натриевого насоса, перекачивающего ионы против градиента концентрации (в противном случае диффузия ионов по градиенту концентрации привела бы к выравниванию концентраций ионов по обе стороны мембраны и гибели клетки) Таким образом, внутри аксона образуется избыток (–) зарядов, снаружи – (+) зарядов, то есть возникает разность электрических потенциалов – потенциал покоя (ПП). Его значение составляет – 60-70 мВ. ПП одинаков по всей длине нервного волокна (рис. 4).
Раздражение нервного волокна приводит сначала к открытию Na + - и К + - каналов. Раньше открываются Na + -каналы, и в клетку устремляется поток ионов натрия. Это изменяет трансмембранный потенциал: сначала он становится равным 0 (деполяризация мембраны), затем происходит перезарядка мембраны, внутренняя сторона мембраны приобретает (+) заряд, наружная (-). Разность потенциалов при этом достигает 40 мВ. Эта величина и есть потенциал действия (ПД)
Натриевые каналы закрываются, открываются калиевые, начинается выход калия из клетки, потенциал изменяется от – 40 мВ до – 70 мВ, то есть идет реполяризация мембраны. После закрытия ионных каналов ионный насос восстанавливает исходное распределение ионов по сторонам мембраны.
Механизмы памяти.
Концепции биохимического кодирования индивидуального опыта в памяти опираются на две группы фактов:
· возможность передачи приобретенной информации необученному мозгу с помощью этих факторов.
Первые гипотезы, связывающие запечатление информации с биохимическими изменениями в нервной ткани родились на основе широко известных в 60-е гг. опытов Г. Хидена, когда было показано, что образование следов памяти сопровождается изменениями свойств РНК и белка в нейронах.
Выяснилось, что раздражение нервной клетки увеличивает в ней содержание РНК и оставляет длительные биохимические следы, сообщающие клетке способность резонировать в ответ на повторные действия одних и тех же раздражителей.
Таким образом, было установлено, что РНК играет важную роль в механизмах формирования и сохранения следов памяти.
Однако в более поздних работах было показано, что в консолидации энграмм памяти ведущую роль играет ДНК, которая может служить хранилищем не только генетической, но и приобретенной информации, а РНК обеспечивает передачу специфического информационного кода.
В настоящее время идея существования биохимических факторов, способных к сохранению и переносу информации, большинством исследователей воспринимается критически. Считается, что гипотеза молекулярного кодирования индивидуального опыта не имеет прямых фактических доказательств.
Медиаторам - химическим посредникам в синаптической передаче информации - придается большое значение в обеспечении механизмов долговременной памяти. Основные медиаторные системы головного мозга принимают самое непосредственное участие в обучении и формировании энграмм памяти. Так, экспериментально установлено, что уменьшение количества норадреналина замедляет обучение, вызывает амнезию и нарушает извлечение следов из памяти.
Р.И. Кругликов (1986) разработал концепцию, в соответствии с которой в основе долговременной памяти лежат сложные структурно-химические преобразования на системном и клеточном уровнях головного мозга. Показано, что под влиянием обучения увеличивается количество холинорецепторов, то есть рецепторов, расположенных на теле нейрона и отвечающих за обнаружение медиатора ацетилхолина. В процессе образования условного рефлекса повышается чувствительность соответствующих нейронов к ацетилхолину, что облегчает обучение, ускоряет запоминание и способствует более быстрому извлечению следа из памяти. В то же время вещества, препятствующие действию ацетилхолина, нарушают обучение и воспроизведение, вызывая амнезию (потерю памяти).
Биохимические методы, которые позволяют проникнуть в последовательность процессов, происходящих в синаптических мембранах с последующим синтезом новых белков, привлекают многих исследователей памяти. На этом пути ожидаются новые яркие открытия. Предполагается, например, что для различных видов памяти в ближайшем будущем будут выявлены различия в биохимических процессах.
Тем не менее следует подчеркнуть, что интенсивные биохимические исследования привели к явной переоценке и автономизации клеточно-молекулярного уровня изучения механизмов памяти. Как указывает С. Роуз, эксперименты, проводимые только на клеточном уровне, слишком ограничены и, по-видимому, не способны ответить на вопрос, как мозг человека запоминает, например, сложные симфонические партитуры или извлекает из памяти данные, необходимые для разгадывания простого кроссворда.
Для более полного знания специфики функционирования процессов памяти необходим переход на уровень сложных мозговых систем, где многие нейроны соединены между собой морфологическими и функциональными связями. При этом психофизиологические исследования на здоровых людях позволяют изучать процессы переработки и хранения информации, а изучение больных с различного рода амнезиями, возникающими после повреждения мозга, позволяет глубже проникать в тайны памяти.
Березов стр. 641-642
Дата добавления: 2019-07-15 ; просмотров: 739 ;
Синапс - это функциональный контакт специализированных участков плазматических мембран двух возбудимых клеток. Синапс состоит из пресинаптической мембраны, синаптической щели и постинаптической мембраны. Мембраны клеток в месте контакта имеют утолщения в виде бляшек - нервных окончаний. Нервный импульс, достигший нервного окончания, не в состоянии преодолеть возникшее перед ним препятствие - синаптическую щель. После этого электрический сигнал преобразуется в химический. Пресинаптическая мембрана содержит специальные канальные белки, подобные белкам, формирующим натриевый канал в мембране аксона. Они тоже реагируют на мембранный потенциал, изменяя свою конформацию и формируют канал. В результате ионы Са2+ проходят через пресинаптическую мембрану по градиенту концентраций в нервное окончание. Градиент концентраций Са2+ создается работой Са2+-зависимой.
АТФазы - кальциевым насосом. Повышение концентрации Са2+ внутри нервного окончания вызывает слияние 200-300 имеющихся там везикул, заполненных ацетилхолином, с плазматической мембраной. Далее ацетилхолин секретируется в синаптическую щель путем экзоцитоза, и присоединяется к рецепторным белкам, расположенным на поверхности постсинаптической мембраны.
Ацетилхолиновый рецептор представляет собой трансмембранный олигомерный гликопротеиновый комплекс, состоящий из 6 субъединиц: 2-, 2-бета, 1-гамма и 1-дельта. Плотность расположения белков-рецепторов в постсинаптической мембране очень велика - около 20000 молекул на 1 мкм2. Пространственная структура рецептора строго сооответствует конформации медиатора. При взаимодействии с ацетилхолином белок-рецептор так изменяет свою конформацию, что внутри него формируется натриевый канал. Катионная селективость канала обеспечивается тем, что ворота канала сформированы отрицательно заряженными аминокислотами. Таким образом повышается проницаемость постсинаптической мембраны для натрия и возникает новый импульс (или сокращение мышечного волокна). Деполяризация постсинаптической мембраны выеывает диссоциацию комплекса "ацетилхолин-белок-рецептор" и ацетилхолин освобождается в синаптическую щель. Как только ацетилхолин оказывается в синаптической щели, он за 40 мкс подвергается быстрому гидролизу под действием фермента ацетилхолинэстеразы.
Во время гидролиза ацетилхолина образуется промежуточный фермент-субстратный комплекс, в котором ацетилхолин связан с активным центром фермента через серин.
Необратимое ингибирование холинэстеразы вызывает смерть. Ингибиторами холинэстеразы являются фосфорорганические соединения (хлорофос, дихлофос, табун, зарин, зоман, бинарные яды). Эти вещества связываются ковалентно с серином в активном центре фермента. Некоторые из них синтезированы в качестве инсектицидов, а некоторые - в качестве боевых отравляющих веществ (нервно-паралитические яды). Смерть наступает в результате остановки дыхания. Обратимые ингибиторы холинэстеразы используются как лечебные препараты. Например, при лечении глаукомы и атонии кишечника.
Катехоламины: норадреналин и дофамин. Адренэргические синапсы встречаются в постганглионарных волокнах, в волокнах симпатической нервной системы, в различных отделах головного мозга. Катехоламины в нервной ткани синтезируются по общему механизму из тирозина. Ключевой фермент синтеза - тирозингидроксилаза, ингибируемая конечными продуктами.
Катехоламины, как и ацетилхолин, накапливаются в синаптических пузырьках и тоже выделяется в синаптическую щель при поступлении нервного импульса. Но регуляция в адренэргическом рецепторе происходит иначе. В пресинаптической мембране здесь имеется специальный регуляторный белок - -ахромогранин (Мм = 77 кДа), который в ответ на повышение концентрации медиатора в синаптической щели связывает уже выделившийся медиатор и прекращает его дальнейший экзоцитоз. Фермента, разрушающего медиатор, в адренэргических синапсах нет. После передачи импульса молекулы медиатора перекачивается специальной транспортной системой путем активного транспорта с участием АТФ обратно через пресинаптическую мембрану и включается вновь в везикулы. В пресинаптическом нервном окончании излишек медиатора может быть инактивирован моноаминоксидазой, а также катехоламин-О-метилтрансферазой путем метилирования по оксигруппе. Кокаин тормозит активный транспорт катехоламинов.
Связывание медиатора с постсинаптическим рецептором почти мгновенно вызывает повышение концентрации ц-АМФ, что приводит к быстрому фосфорилированию белков постсинаптической мембраны. В результате изменяется генерация нервных импульсов постсинаптической мембраной (тормозится). В некторых случаях непосредственной причиной этого является повышение проницаемости постсинаптической мембраны для калия, либо снижением проводимости для натрия (эти события приводят к гиперполяризации).
ГАМК - тормозной медиатор. Повышает проницаемость постсинаптических мембран для ионов калия. Это ведет к изменению мембранного потенциала.
Глицин - тормозной медиатор, по вызываемым эффектам подобен гамк.
Образование нейромедиаторов – ацетилхолина, адреналина, дофамина, серотонина.
Информация, благодаря которой нейроны устанавливают только определенные связи с определенными нейронами, кодируется в структуре полисахаридных веточек мембранных гликопротеинов. Образование таких связей, не заложенных в период эмбрионального развития, является результатом опыта индивидуального организма и составляет материальную основу для хранения информации, определяющей особенности поведения данного организма.
Нейромедиаторы - это вещества, которые характеризуются следующими признаками. Накапливаются в пресинаптической структуре в достаточной концентрации. Освобождаются при передаче импульса. Вызывают после связывания с постсинаптической мембраной изменение скорости метаболических процессов и возникновение электрического импульса. Имеют систему для инактивации или транпортную систему для удаления из синапса, обладающие к ним высоким сродством. Таким образом, нейромедиаторы играют важную роль в функционировании нервной ткани, обеспечивая синаптическую передачу нервного импульса. Их синтез происходит в теле нейронов, а накопление - в особых везикулах, которые постепенно перемещаются с участием систем нейрофиламентов и нейротрубочек к кончикам аксонов.
Химическая классификация нейромедиаторов: Аминокислоты (и их производные) - К ним относят таурин, норадреналин, ДОФАминГАМК, глицин, ацетилхолин, гомоцистеин и некоторые другие (адреналин, серотонин, гистамин, серотонин).
Таурин - образуется из аминокислоты цистеина. Сначала происходит окисление серы в SH-группе до остатка серной кислоты (процесс идет в несколько стадий), а затем происходит декарбоксилирование. Таурин - это необычная кислота, в которой нет карбоксильной группы, а имеется остаток серной кислоты. Таурин принимает участие в проведении нервного импульса в процессе зрительного восприятия.
Ацетилхолин - Для синтеза холина требуются аминокислоты серин, метионин. Этаноламин может быть использован и в готовом виде. Но, как правило, из крови в нервную ткань пступает уже готовый холин. Второй же предшественник этого нейромедиатора - Ацетил-КоА, синтезируется в нервных окончаниях. Продукт этой реакции ацетилхолин участвует в синаптической передаче нервного импульса. Он накапливается в синаптических пузырьках, образуя комплексы с отрицательно заряженным белком везикулином. Передача возбуждения с одной клетки на другую осуществляется с помощью специального синаптического механизма.
Норадреналин - медиатор в постганглионарных волокнах симпатической и в различных отделах ЦНС.
Дофамин- медиатор проводящих путей, тела нейронов которого расположены в отделе мозга, который отвечает за контроль произвольных движений. Поэтому при нарушении дофаминэргической передачи возникает заболевание паркинсонизм.
Последнее изменение этой страницы: 2016-08-16; Нарушение авторского права страницы
Структура нервного волокна. Проведение нервных импульсов является специализированной функцией нервных волокон, т.е. отростков нервных клеток.
Нервные волокна разделяют намякотные, или миелинизированные, и безмякотные, или немиелинизированные. Мякотные, чувствительные и двигательные волокна входят в состав нервов, снабжающих органы чувств и скелетную мускулатуру; они имеются также в вегетативной нервной системе. Безмякотные волокна у позвоночных животных принадлежат в основном симпатической нервной системе.
Нервы обычно состоят как из мякотных, так и из безмякотных волокон, причем их соотношение в разных нервах различное. Например, во многих кожных нервах преобладают безмякотные нервные волокна. Так, в нервах вегетативной нервной системы, например в блуждающем нерве, количество безмякотных волокон достигает 80—95%. Наоборот, в нервах, иннервирующих скелетные мышцы, имеется лишь относительно небольшое количество безмякотных волокон.
Как показали электронно-микроскопические исследования, мие- линовая оболочка создается в результате того, что миелоцит (шван- новская клетка) многократно обертывает осевой цилиндр (рис. 2.27'), слои ее сливаются, образуя плотный жировой футляр — миелиновую оболочку. Миелиновая оболочка через промежутки равной длины прерывается, оставляя открытыми участки мембраны шириной примерно 1 мкм. Эти участки получили название перехватов Ранвье.

Рис. 2.27. Роль миелоцита (шванновской клетки) в образовании миелиновой оболочки в мякотных нервных волокнах: последовательные стадии спиралеобразного закручивания миелоцита вокруг аксона (I); взаимное расположение миелоцитов и аксонов в безмякотных нервных волокнах (II)
Длина межперехватных участков, покрытых миелиновой оболочкой, примерно пропорциональна диаметру волокна. Так, в нервных волокнах диаметром 10—20 мкм длина промежутка между перехватами составляет 1—2 мм. В наиболее тонких волокнах (диаметром
1—2 мкм) эти участки имеют длину около 0,2 мм. [1]
Безмякотные нервные волокна не имеют миелиновой оболочки, они изолированы друг от друг только шванновскими клетками. В простейшем случае одиночный миелоцит окружает одно безмякот- ное волокно. Часто, однако, в складках миелоцита оказывается несколько тонких безмякотных волокон.
Миелиновая оболочка выполняет двоякую функцию: функцию электрического изолятора и трофическую функцию. Изолирующие свойства миелиновой оболочки связаны с тем, что миелин как вещество липидной природы препятствует прохождению ионов и потому обладает очень высоким сопротивлением. Благодаря существованию миелиновой оболочки возникновение возбуждения в мякот- ных нервных волокнах возможно не на всем протяжении осевого цилиндра, а только в ограниченных участках — перехватах Ранвье. Это имеет важное значение для распространения нервного импульса вдоль волокна.
Трофическая функция миелиновой оболочки, по-видимому, состоит в том, что она принимает участие в процессах регуляции обмена веществ и роста осевого цилиндра.
Прямые электрофизиологические исследования, проведенные Като (1924), а затем Тасаки (1953) на одиночных миелинизированных нервных волокнах лягушки, показали, что потенциалы действия в этих волокнах возникают только в перехватах, а участки между перехватами, покрытые миелином, являются практически невозбудимыми.
В состоянии покоя наружная поверхность возбудимой мембраны всех перехватов Ранвье заряжена положительно. Разности потенциалов между соседними перехватами не существует. В момент возбуждения поверхность мембраны перехвата С становится заряженной электроотрицательно по отношению к поверхности мембраны соседнего перехвата D. Это приводит к возникновению местного (ло 
Рис. 2.28. Сальтаторное распространение возбуждения в мякотном нервном волокне от перехвата к перехвату:
А — немиелинизированное волокно; В — миелинизированное волокно. Стрелками показано направление тока
кального) электрического тока, который идет через окружающую волокно межтканевую жидкость, мембрану и аксоплазму в направлении, показанном на рисунке стрелкой. Выходящий через перехват D ток возбуждает его, вызывая перезарядку мембраны. В перехвате С возбуждение еще продолжается, и он на время становится рефрактерным. Поэтому перехват D способен привести в состояние возбуждения только следующий перехват и т.д.
СТРУКТУРА НЕРВНЫХ ВОЛОКОН
Проведение нервных импульсов является специализированной функцией нервных волокон, т. е. отростков нервных клеток.
Нервные волокна разделяют на мякотные, или миелинизированные, и безмякотные, немиелинизированные. Мякотные, чувствительные и двигательные волокна входят в состав нервов, снабжающих органы чувств и скелетную мускулатуру; они имеются также в вегетативной нервной системе. Безмякотные волокна у позвоночных животных принадлежат в основном симпатической нервной системе.
Нервы обычно состоят как из мякотных, так и из безмякотных волокон, причем соотношение между числом тех и других в разных нервах различное. Например, во многих кожных нервах преобладают безмякотные нервные волокна. Так, в нервах вегетативной нервной системы, например в блуждающем нерве, количество безмякотных волокон достигает 80—95 %. Наоборот, в нервах, иннервирующих скелетные мышцы, имеется лишь относительно небольшое количество безмякотных волокон.
На рис. 42 схематически показано строение миелинизированного нервного волокна. Как видно, оно состоит из осевого цилиндра и покрывающей его миелиновой оболочки. Поверхность осевого цилиндра образована плазматической мембраной, а его содержимое представляет собой аксоплазму, пронизанную тончайшими (диаметром 10—40 нм) нейрофибриллами (и микротубулами), между которыми находится большое количество митохондрий и микросом. Диаметр нервных волокон колеблется от 0,5 до 25 мкм.
Как показали электронно-микроскопические исследования, миелиновая оболочка создается в результате того, что миелоцит (шванновская клетка) многократно обертывает осевой цилиндр (рис. 43, I), слои ее сливаются, образуя плотный жировой футляр — миелиновую оболочку. Миелиновая оболочка через промежутки равной длины прерывается, оставляя открытыми участки мембраны шириной примерно 1 мкм. Эти участки получили название перехватов (перехваты Ранвье).
Длина межперехватных участков, покрытых миелиновой оболочкой, примерно пропорциональна диаметру волокна. Так, в нервных волокнах, имеющих диаметр 10— 20 мкм, длина промежутка между перехватами составляет 1—2 мм. В наиболее тонких волокнах (диаметром 1—2 мкм) эти участки имеют длину около 0,2 мм.
Безмякотные нервные волокна не имеют миелиновой оболочки, они изолированы друг от друга только шванновскими клетками. В простейшем случае одиночный миелоцит окружает одно безмякотное волокно. Часто, однако, в складках миелоцита оказывается несколько тонких безмякотных волокон (рис. 43. II).

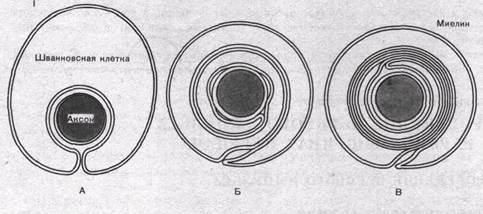
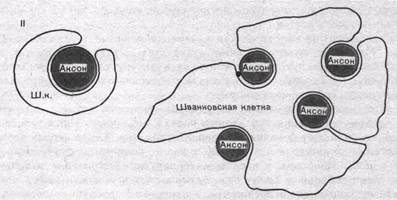
Рис.43. Роль миелоцита (шванновской клетки) в образовании миелиновой оболочки в мякотных нервных волокнах. Показаны последовательные стадии спиралеобразного закручивания миелоцита вокруг аксона (I). Взаимное расположение миелоцитов и аксонов в безмякотных нервных волокнах (II).
ФИЗИОЛОГИЧЕСКАЯ РОЛЬ СТРУКТУРНЫХ ЭЛЕМЕНТОВ МИЕЛИНИЗИРОВАННОГО НЕРВНОГО ВОЛОКНА
Можно считать доказанным, что в процессах возникновения и проведения нервного импульса основную роль играет поверхностная мембрана осевого цилиндра. Миелиновая оболочка выполняет двоякую функцию: функцию электрического изолятора и трофическую функцию. Изолирующие свойства миелиновой оболочки связаны с тем, что миелин как вещество липидной природы препятствует прохождению ионов и потому обладает очень высоким сопротивлением. Благодаря существованию миелиновой оболочки возникновение возбуждения в мякотных нервных волокнах возможно не на всем протяжении осевого цилиндра, а только в ограниченных участках — перехватах узла (перехвата Ранвье). Это имеет важное значение для распространения нервного импульса вдоль волокна.
Трофическая функция миелиновой оболочки, по-видимому, состоит в том, что она принимает участие в процессах регуляции обмена веществ и роста осевого цилиндра.
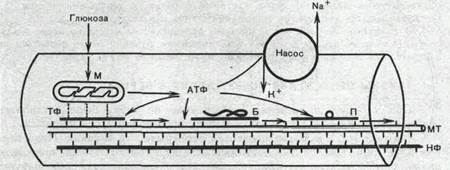
Рис.44. Гипотетический транспортный механизм нервного волокна.
Предполагается, что микротубулы (МТ) и нейрофиламенты (НФ) образованы миозином, а тонкие транспортные филаменты — актином. При расщеплении АТФ транспортные фила менты скользят вдоль микротрубочек и таким образом переносят прикрепившиеся к ним митохондрии (М), молекулы белка (Б) или пузырьки (П) с медиатором. АТФ продуцируется митохондриями в результате распада глюкозы, проникающей в волокно. Энергия АТФ частично используется также натриевым насосом поверхностной мембраны.
Нейрофибриллы, микротубулы и транспортные филаменты обеспечивают транспорт различных веществ и некоторых клеточных органелл по нервным волокнам от тела нейрона к нервным окончаниям и в обратном направлении. Так, по аксону из тела клетки на периферию транспортируются: белки, формирующие ионные каналы и насосы;
возбуждающие и тормозные медиаторы; митохондрии. Подсчитано, что через поперечный разрез среднего по диаметру аксона в течение суток перемещается примерно 1000 митохондрии.
Так же как и мышечный сократительный аппарат, транспортная система нервного волокна использует для своей работы энергию АТФ и нуждается в присутствии ионов Ca 2+ в цитоплазме.
ПЕРЕРОЖДЕНИЕ НЕРВНЫХ ВОЛОКОН ПОСЛЕ ПЕРЕРЕЗКИ НЕРВА
Нервные волокна не могут существовать вне связи с телом нервной клетки: перерезка нерва ведет к гибели тех волокон, которые оказались отделенными от тела клеток. У теплокровных животных уже через 2—3 сут после перерезки нерва периферический его отросток утрачивает способность к проведению нервных импульсов. Вслед за этим начинается дегенерация нервных волокон, причем миелиновая оболочка претерпевает жировое перерождение. Это выражается в том, что мякотная оболочка теряет миелин, который скапливается в виде капель; распавшиеся волокна и их миелин рассасываются и на месте нервных волокон остаются тяжи, образованные леммоцитом (шванновской клеткой). Все эти изменения впервые были описаны английским врачом Валлером и названы по его имени валлеровским перерождением.
Регенерация нерва происходит очень медленно. Леммоциты, оставшиеся на месте дегенерировавших нервных волокон, начинают разрастаться вблизи места перерезки по направлению к центральному отрезку нерва. Одновременно перерезанные концы аксонов центрального отрезка образуют так называемые колбы роста — утолщения, которые растут в направлении периферического отрезка. Часть этих веточек попадает в старое ложе перерезанного нерва и продолжает расти в этом ложе со скоростью 0,5—4,5 мм в сутки до тех пор, пока не дойдет до соответствующей периферической ткани или органа, где волокна образуют нервные окончания. С этого времени восстанавливается нормальная иннервация органа или ткани.
В различных органах восстановление функции после перерезки нерва наступает в разные сроки. В мышцах первые признаки восстановления функций могут появиться через 5—6 нед;
окончательное восстановление происходит много позднее, иногда через год.
ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ
При изучении проведения возбуждения по нерву было установлено несколько необходимых условий и правил (законов) протекания этого процесса.
Анатомическая и физиологическая непрерывность волокна. Проведение импульсов возможно лишь при условии анатомической целостности волокна, поэтому как церерезка нервных волокон, так и любая травма поверхностной мембраны нарушают проводимость. Непроводимость наблюдается также при нарушении физиологической целостности волокна (блокада натриевых каналов возбудимой мембраны тетродотоксином или местными анестетиками, резкое охлаждение и т. п.). Проведение нарушается и при стойкой деполяризации мембраны нервного волокна ионами К, накапливающимися при ишемии в межклеточных щелях. Механическая травма, сдавливание нерва при воспалительном отеке тканей могут сопровождаться частичным или полным нарушением функции проведения.
Двустороннее проведение. При раздражении нервного волокна возбуждение распространяется по нему и в центробежном, и в центростремительном направлениях. Это доказывается следующим опытом.
К нервному волокну, двигательному или чувствительному, прикладывают две пары электродов, связанных с двумя электроизмерительными приборами А и Б (рис. 45). Раздражение наносят между этими электродами. В результате двустороннего проведения возбуждения приборы зарегистрируют прохождение импульса как под электродом А, так и под электродом Б.
Двустороннее проведение не является только лабораторным феноменом. В естественных условиях потенциал действия нервной клетки возникает в той ее части, где тело переходит в ее отросток— аксон (так называемый начальный сегмент). Из начального сегмента потенциал действия распространяется двусторонне: в аксоне по направлению к нервным окончаниям и в тело клетки по направлению к ее дендритам.
Изолированное проведение. В периферическом нерве импульсы распространяются по каждому волокну изолированно, т. е. не переходя с одного волокна на другое и оказывая действие только на те клетки, с которыми контактируют окончания данного нервного волокна. Это имеет очень важное значение в связи с тем, что всякий периферический нервный ствол содержит большое число нервных волокон —двигательных, чувствительных и вегетативных, которые иннервируют разные, иногда далеко отстоящие друг от друга и разнородные по структуре и функциям клетки и ткани. Например, блуждающий нерв иннервирует все органы грудной полости и значительную часть органов брюшной полости, седалищный нерв — всю мускулатуру, костный аппарат, сосуды и кожу нижней конечности. Если бы возбуждение переходило внутри нервного ствола с одного волокна на другое, то в этом случае нормальное функционирование периферических органов и тканей было бы невозможно-Изолированное проведение в отдельных волокнах смешанного нерва может быть доказано простым опытом на скелетной мышце, иннервированной смешанным нервом, в образовании которого участвует несколько спинномозговых корешков. Если раздражать один из этих корешков, сокращается не вся мышца, как это было бы в случае перехода возбуждения с одних нервных волокон на другие, а только те группы мышечных волокон, которые иннервированы раздражаемым корешком. Еще более строгое доказательство изолированного проведения возбуждения может быть получено при отведении потенциалов действия от различных нервных волокон нервного ствола.
Изолированное проведение нервного импульса обусловлено тем, что сопротивление жидкости, заполняющей межклеточные щели, значительно ниже сопротивления мем-
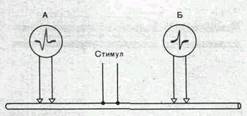 |
Рис.45. Схематическое изображение опыта для доказательства двустороннего проведения импульса в нерве. Объяснение в тексте.
браны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным (деполяризованным) и покоящимися участками возбудимой мембраны, проходит по межклеточным щелям, не заходя в соседние волокна.
Теория по нормальной физиологии на тему: Синапс. Физиология мышечных волокон. В данной статье рассматривается механизм синаптической передачи, НМС…
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Синапс — это специфическое место контакта двух возбудимых систем (клеток) для передачи возбуждения.
По способу передачи сигналов:
- механические,
- химические,
- электрические.
По виду медиатора: холинэргические и др.
Нервно-мышечный синапс (НМС) — химический, передача с помощью медиатора ацетилхолина.
Синонимы к слову НМС:
- Нервно-мышечное соединение;
- Моторная концевая пластинка.

Аксоны нервных клеток на своих окончаниях теряют миелиновую оболочку , ветвятся, и концевые веточки аксона утолщаются. Это пресинаптическая терминаль или бляшка или пуговка , которая погружается в углубление на поверхности мышечного волокна.
Покрывающая концевую веточку аксона поверхностная мембрана называется пресинаптической мембраной , т.е. это мембрана, покрывающая поверхность синаптической бляшки (терминали аксона).
Мембрана, покрывающая мышечное волокно в области синапса, называется постсинаптической мембраной , или концевой пластинкой. Она имеет извитую структуру, образуя многочисленные складки, уходящие вглубь мышечного волокна, за счет чего увеличивается площадь контакта.

На постсинаптической мембране находятся белковые структуры — рецепторы , способные связывать медиатор . В одном синапсе количество рецепторов достигает 10-20 млн.
Между пре- и постсинаптическими мембранами находится синаптическая щель , размеры ее в среднем 50 нм, она открывается в межклеточное пространство и заполнена межклеточной жидкостью .
В синаптической щели находится мукополисахаридное плотное вещество в виде полосок, мостиков и содержится фермент ацетилхолинэстераза .
В пресинаптической терминали находится большое количество пузырьков или везикул , заполненных медиатором — химическим веществом посредником, осуществляющим передачу возбуждения.
В нервно-мышечном синапсе медиатор — ацетилхолин (АХ).

АХ синтезируется из холина и уксусной кислоты (ацетил-коэнзима А) с помощью фермента холинэстеразы. Эти вещества перемещаются из тела нейрона по аксону к пресинаптической мембране. Здесь в пузырьках происходит окончательное образование АХ.
3 фракции медиатора:
- Первая фракция — доступная — располагается рядом с пресинаптической мембраной.
- Вторая фракция — депонированная — располагается над первой фракцией.
- Третья фракция — диффузно рассеянная — наиболее удаленная от пресинаптической мембраны.
Механизм синаптической передачи



Ионы Ca вызывают образование специального белкового комплекса , который включает в себя везикулу и структуры, расположенные непосредственно около пресинаптической мембраны.
Они связаны между собой так называемыми белками экзоцитоза.
Часть белков расположена на везикулах (синапсин, синаптотагмин, синаптобревин), а часть — на пресинаптической мембране (синтаксин, синапсо-ассоциированный белок). Данный комплекс получил название секретосома .



Излитию содержимого пузырька в щель способствует белок синаптопорин , формирующий канал, по которому идет выброс медиатора.
Квант медиатора — количество молекул, содержащихся в одной везикуле.
На 1 ПД выбрасывается 100 квантов АХ.





На постсинаптической мембране возникает потенциал концевой пластинки (ПКП). Он является аналогом локального ответа (ЛО).
Потенциал действия на постсинаптической мембране не возникает ! Он формируется на соседней мембране мышечного волокна.
- связывание с рецептором,
- разрушение ферментов (ацетилхолинэстеразой),
- обратное поглощение в пресинаптическую мембрану,
- вымывание из щели и фагоцитоз.
События в синапсе :
- ПД приходит к терминали аксона;
- Он деполяризует пресинаптическую мембрану;
- Ca2+ входит в терминаль, что приводит к выделению АХ;
- В синаптическую щель выделяется медиатор АХ;
- Он диффундирует в щель и связывается с рецепторами постсинаптической мембраны;
- Меняется проницаемость постсинаптической мембраны для ионов Na+;
- Ионы Na+ проникают в постсинаптическую мембрану и уменьшают ее заряд — возникает потенциал концевой пластинки (ПКП) .
На самой постсинаптической мембране ПД возникнуть не может, так как здесь отсутствуют потенциалзависимые каналы, они являются хемозависимыми!
- ПКП суммируются и достигают КУД на соседнем участке мышечного волокна, что приводит к возникновению ПД и его распространению по мышечному волокну (около 5 м/с).
Достигнув пороговой величины, то есть КУД, ПКП возбуждает соседнюю (внесинаптическую) мембрану мышечного волокна за счет местных круговых токов.

Особенности проведения возбуждения в нервно-мышечном синапсе
Одностороннее проведение возбуждения — только в направлении от пресинаптического окончания к постсинаптической мембране.
Суммация возбуждения соседних постсинаптических мембран.
Синаптическая задержка — замедление в проведении импульса от нейрона к мышце составляет 0,5-1 мс. Это время затрачивается на секрецию медиатора, его диффузию к постсинаптической мембране, взаимодействие с рецептором, формирование ПКП, их суммацию.
Низкая лабильность — она составляет 100-150 имп/с для сигнала, что в 5-6 раз ниже лабильности нервного волокна.
Чувствительность к действию лекарственных веществ, ядов, БАВ, выполняющих роль медиатора.
Утомляемость химических синапсов — выражается в ухудшении проводимости вплоть до блокады в синапсе при длительном функционировании синапса. Главная причина утомляемости — исчерпание запасов медиатора в пресинаптическом окончании.
Законы проведения возбуждения по нервам:
- Закон функциональной целостности нерва.
- Закон изолированного проведения возбуждения.
- Закон двустороннего проведения возбуждения.
В зависимости от скорости проведения возбуждения нервные волокна подразделяются на 3 группы: A, B, C. В группе A выделяют 4 подгруппы: альфа, бетта, гамма и сигма.
Физиология мышечных волокон
- скелетная (40-50% массы тела),
- сердечная (менее 1%),
- гладкая (8-9%).
Физиологические свойства скелетных мышц:
- Возбудимость — способность отвечать на действие раздражителя возбуждением.
- Проводимость — способность проводить возбуждение из места его возникновения к другим участкам мышцы.
- Лабильность — способность мышцы сокращаться в соответствии с частотой действия раздражителя (200-300 Гц для скелетной мышцы).
- Сократимость — для мышцы является специфическим свойством — это способность мышцы изменять длину или напряжение в ответ на действие раздражителя.
Физические свойства скелетных мышц:
- Растяжимость — способность мышцы изменять длину под действием растягивающей силы.
- Эластичность — способность мышцы восстанавливать первоначальную длину или форму после прекращения действия растягивающей силы.
- Силы мышц — способность мышцы поднять максимальный груз.
- Способность мышцы совершать работу.
Режимы сокращения:
- Изотонический,
- Изометрический,
- Ауксотонический.
Изотонический режим — сокращение мышцы происходит с изменением ее длины без изменения напряжения (тонуса) (напр.: сокращение мышц языка).
Изометрический режим — длина постоянная, увеличивается степень мышечного напряжения (тонуса) (напр.: при поднятии непосильного груза).
Ауксотонический режим — одновременно изменяется длина и напряжение мышцы (характерен для обычных двигательных актов).
Механизм сокращения поперечно-полосатых мышц
Любая скелетная мышца состоит из мышечных волокон, которые, в свою очередь, состоят из множества тонких нитей — миофибрилл , расположенных продольно. Каждая миофибрилла состоит из протофибрилл — нитей сократительных белков: миозина (миозиновая протофибрилла), актина (актиновая протофибрилла).
Кроме сократительных белков в миофибрилле имеются два регуляторных белка: тропомиозин и тропонин .
Миозиновые волокна соединены в толстый пучок, от которого в торону актиновых нитей отходят поперечные мостики. У каждого мостика выделяют шейку и головку.
Нить актина располагается в виде 2 скрученных ниток бус. На ней имеются актиновые центры.

Тропомиозин в виде спиралей оплетает поверхность актина, закрывая в покое ее центры. Одна молекула тропомиозина контактирует с 7 молекулами актина.
Тропонин образует утолщение на конце каждой нити тропомиозина.
Под влиянием возникшего в мышечном волокне ПД из саркоплазматического ретикулума (СПР — депо Ca2+) высвобождаются ионы Ca. Кальций связывается с тропонином, который смещает тропомиозиновый стержень, что приводит к открытию актиновых центров.

В результате, к актиновым центрам присоединяются головки поперечных миозиновых мостиков.
Процесс расслабления происходит в обратной последовательности с использованием энергии АТФ за счет функционирования кальциевого насоса.
При отсутствии повторного импульса ионы Ca не поступают из СПР. В результате отсутствия Ca-тропонинового комплекса, тропомиозин возвращается на свое прежнее место, блокируя актиновые центры актина. Актиновые протофибриллы легко скользят в обратном направлении благодаря эластичности мышцы, и мышца удлиняется (расслабляется).
Гладкие мышцы
Гладкие мышцы — это мышцы, формирующие слой стенок полых внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток без поперечной исчерченности за счет хаотичного расположения миофибрилл.
Особенности гладких мышц:
- Иннервируются волокнами вегетативной нервной системы (ВНС);
- Обладают низкой возбудимостью:
- Обладают низкой величиной МП (мембранного потенциала) — -50 — -60 мВ из-за более высокой проницаемости для ионов Na+
- ПД (потенциал действия) отличается меньшей амплитудой и большей длительностью. Он формируется в основном за счет ионов Ca2+
- Медленная проводимость:
Клетки в гладких мышцах функционально связаны между собой посредством щелевидных контактов — нексусов, которые имеют низкое электрическое сопротивление. За счет этих контактов ПД распространяется с одного мышечного волокна на другое, охватывая большие мышечные пласты, и в реакцию вовлекается вся мышца.
Гладкие мышцы способны осуществлять относительно медленные ритмические и длительные тонические сокращения.
Медленные ритмические сокращения обеспечивают перемещение содержимого органа из одной области в другую.
Длительные тонические сокращения, особенно сфинктеров полых органов, препятствуют выходу из них содержимого.
Это способность сохранять приданную им при растяжении или деформации форму. Благодаря пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии.
Особенность гладких мышц, отличающая их от скелетных. Благодаря автоматии гладкие мышцы могут сокращаться в условиях отсутствия иннервации . Важную роль в этом играет растяжение.
Растяжение является адекватным раздражителем для гладкой мускулатуры. Сильное и резкое растяжение гладких мышц вызывает их сокращение.
Сравнительная характеристика скелетных и гладких мышц:
Читайте также:
