
Это морфологическое изменение нервной ткани
Нейроны и нейроглия
Нейроны и нейроглия
Нервную систему формирует нервная ткань. Клетки образующие нервную ткань делятся на две группы – это нейроны и клетки глии.
Определение_1:
Нейрон - это структурно-функциональная клеточная единица нервной системы, которая с помощью своих отростков осуществляет контакты с другими нейронами и участвует в образовании рефлекторных дуг.
Нейроны отличаются от других клеток организма следующими свойствами:
способностью приходить в состояние возбуждения, т.е. реагировать на раздражение изменением обмена веществ и свойств клеточной мембраны,
способность принимать сигнал от другой клетки;
способностью проводить возбуждение (импульс) по нервным волокнам, и только в аксопетальном, т.е. от дендритов к аксону, направлении;
генерировать нервное возбуждение;
нейроны - высокоспециализированные клетки;
в течение жизни нейроны не делятся;
- структуры нейрона могут регенерировать (например, аксон, или дендриты).
В зависимости от выполняемой функции в составе рефлекторной дуге различают:
– нейроны рецепторы (чувствительные, афферентные),
– вставочные (ассоциативные) нейроны,
– двигательные (эфферентные) нейроны.
Афферентные нейроны воспринимают импульс, эфферентные передают его на ткани рабочих органов, побуждая их к действию, а ассоциативные осуществляют связь между нейронами.
Нейроны отличаются большим разнообразием форм и размеров. Диаметр тел клеток-зерен коры мозжечка 4-6 мкм, а гигантских пирамидных нейронов двигательной зоны коры большого мозга – 130-150 мкм.

В нейроне выделяют перикарион (тело) и отростки. Перикарион состоит из клеточной оболочки (мембраны), ядра и цитоплазмы. Основная функция перикариона состоит в осуществлении обмена веществ. Перикарионы образуют серое вещество нервной ткани.
Ядро занимает центральное положение, содержит мало хроматина, хорошо выраженное ядрышко. Из-за высокой активности метаболизма нейронов хроматин в их ядрах находится в деспирализованом (раскрученном) состоянии, в результате чего происходит непрерывное считывание генетической информации (транскрипция) и образование (трансляция) нейропептидов и белков.
Снаружи нейрон покрыт клеточной мембраной (нейролеммой), которая обеспечивает следующие функции:
- транспортная функция состоит в обеспечении транспорта ионов и биологически активных веществ (нейромедиаторов и гормонов);
- рецепторная функция состоит в восприятии внешних сигналов;
- обеспечивает проведение импульса вдоль нервного волокна.
Цитоплазматическая мембрана нейрона обладает способностью генерировать и проводить импульс, за счет встроенных в нее интегральных белков. Они работают как ионно-избирательные каналы – пропуская одни ионы и не позволяя проходить в клетку другим ионам. В результате на мембране формируется разность потенциалов, создающая возможность проведения нервного импульса.
Цитоплазма нейронов состоит из гиалоплазмы, органоидов и включений. Основные органоиды нейрона:
- элементы цитоскелета (микротрубочки, микрофиламенты и микрофибриллы)
- рибосомы и полисомы
В качестве включений в нейронах встречаются липидные (жировые) капли, гранулы липофусцина и меланина.
Дендриты – это выпячивания перикариона. Они содержат те же органеллы, что и перикарион:
гранулярная эндоплазматическая сеть,
полисомы (скопления рибосом),
нейротубулы (микротрубочек) и нейрофиламентов.
На поверхности дендритов характерно наличие тонких шипикообразных отростков длиной 2-3 мкм. За счет дендритов рецепторная поверхность нейрона увеличивается в 1000 и более раз.
Определение_2:
Аксон (или нейрит) – отросток, по которому импульс передается от тела клетки на орган эффектор.
Длина аксона может достигать 1 м (например, клеток блуждающего нерва). Аксон имеет нитевидную форму и является основой нервного волокна. Аксон начинается в виде не покрытого дополнительной оболочкой участка, который называется аксональный холмик, который затем переходит в продолжение – осевой цилиндр. Аксональный холмик наиболее возбудимый участок аксона, является местом генерации нервных импульсов. Совокупность нескольких аксонов формирует нервное волокно, направленное к рабочему органу (эффектору).
Цитоплазма аксона (аксоплазма) содержит микротрубочки, нейрофиламенты, митохондрии, ЭПС, синаптические пузырьки, заполненные нейромедиаторами и плотные тельца. Перемещение аксоплазмы в нейронах идет со скоростью 1-5 мм/сутки, что способствует непрерывному обновлению структурных белков аксона.
По количеству отростков различают:
– униполярные нейроны, имеющие только аксон,
– биполярные, имеющие аксон и один дендрит (встречаются в органах чувств);
– псевдоуниполярный – от тела отходит один общий вырост – отросток, разделяющийся затем на дендрит и аксон (присутствуют в спинальных ганглиях);
– мультиполярные, имеющие аксон и много дендритов (большинство нейронов мультиполярные).
Аксоны образуют белое вещество нервной ткани.
Аксон оканчиваются разветвлениями, которые называются телодендронами (telodendron). Каждый телодендрон заканчиваются утолщением – нервным окончанием.
Нейроны – высокоспециализированные клетки, существующие и функционирующие в строго определенной среде. Такую среду им обеспечивает нейроглия. Нейроглия выполняет следующие функции:
поддержание постоянства среды вокруг нейронов,
Различают глию центральной и периферической нервной системы. Клетки глии центральной нервной системы делятся на:
К макроглии относятся:
Эпендимоциты выстилают желудочки головного мозга и центральный канал спинного мозга. Большинство эпендимоцитов имеют подвижные реснички, вызывающие ток цереброспинальной жидкости. Эпендимный эпителий сосудистых сплетений желудочков мозга продуцирует цереброспинальную жидкость.
Астроциты – клетки отростчатой формы. Они выполняют в основном опорную и разграничительную функции. Астроциты накапливают и передают вещества от капилляров к нейронам, захватывают избыток экстрацеллюлярного калия и других веществ, таких как нейромедиаторы, из экстрацеллюлярного пространства после интенсивной нейрональной активности.
Олигодендроциты (oligodendrocyte) имеют мелкие ядра и немногочисленны отростки. Олигодендроглиоциты присутствуют как в сером, так и в белом веществе. В сером веществе они локализуются вблизи перикарионов. В белом веществе их отростки образуют миелиновый слой в нервных волокнах.
Глия периферической нервной системы (периферическая нейроглия) включает нейролеммоциты (Шванновские клетки) и глиоциты ганглиев (мантийные глиоциты).
Нейролеммоциты формируют оболочки отростков нервных клеток в нервных волокнах периферической нервной системы. Глиоциты ганглиев окружают тела нейронов в нервных узлах и участвуют в обмене веществ нейронов.
Цель:познакомиться с различными видами нейронов, особенностями их строения и типами соединения.
Содержание занятия:
- Строение нервной клетки. Перикарион, отростки нервной клетки
- Классификация нейронов
- Синапсы.
Навыки и умения:
Перечень основных анатомических понятий и образований, которые студент должен уметь: а) показать на препарате или на рисунке; б) назвать по-русски; в) определить по строению и функции:
¾ Везикулы с медиатором
Методические указания:
Рассмотреть в микроскоп гистологические препараты нервных клеток на маленьком увеличении (×8) и большом увеличении (×40). Прочитать описания препаратов. Найти соответствие описания и препарата. Зарисовать изображения препаратов в тетрадь с короткими описаниями.
Познакомится с описаниями клеточных контактов нервных клеток – синапсами. Рассмотреть на таблицах и рисунках анатомические особенности синапсов. Зарисовать и описать химический синапс с кратким описанием.
Рис. 1. Нервные клетки спинномозгового ганглия собаки.
Чувствительные нейроны млекопитающих и человека располагаются вне центральной нервной системы, чаще всего они входят в состав спинномозговых узлов, расположенных в непосредственной близости от спинного мозга. При малом увеличении видно, что спинномозговой узел покрыт капсулой из плотной соединительной ткани с элементами рыхлой соединительной ткани, содержащей жировые клетки. Плотная соединительная ткань проникает в глубь органа, образуя его остов, в ней проходят кровеносные сосуды. Тела нейронов (1) расположены под капсулой спинномозгового узла и в его центральной части по ходу нервных волокон (2), являющихся нейритами и дендритами этих нейронов. Спинномозговой узел спаян с пучками нервных волокон, образованных нейритами мотонейронов спинного мозга. Среди этих нервных волокон нейроны отсутствуют. При большом увеличении видны значительных размеров перикарионы округлой формы (1). Крупное пузыревидное ядро (2) с четко выраженной ядерной мембраной, небольшим количеством мелких глыбок хроматина, крупным оксифильным ядрышком (3) располагается центрально. Если срез прошел вне зоны расположения ядра или по касательной к телу нейрона, клетки (4) выглядят безъядерными. Чем дальше от экваториальной плоскости нейрона прошел срез, тем более мелким выглядит нейрон. Вокруг ядра находятся мелкая зернистость и небольшие глыбки (5). Это глыбки Ниссля, выявляющиеся в чувствительных нейронах при обычной гистологической обработке. Неодинаковая степень хроматофилии нейроплазмы (наличие темных и светлых клеток) служит морфологическим выражением различного функционального состояния нейроцитов. Вокруг перикариона находятся круглые, бедные хроматином ядра (6) мантийных глиоцитов, представляющих олигодендроглию. Глиальная оболочка покрыта тонкой соединительнотканной капсулой. Овальные ядра (7) соединительнотканных клеток богаты хроматином. В результате фиксации нейроциты сжимаются сильнее соединительнотканной капсулы, между ними образуется щель, представляющая артефакт. Обычная окраска не выявляет отростки нейронов. Импрегнация серебром позволяет выявить, что афферентные нейроны являются псевдоуниполярными (ложноодноотростчатыми). В процессе эмбриогенеза дендрит и нейрит биполярного нейробласта сближаются, их основания сливаются. Дендрит и нейрит начинаются от общего выроста тела нейроцита, создающего впечатление одного отростка, с последующим Т-образным делением. Несмотря на изменение формы тела, биполярность афферентного нейрона сохраняется в постэмбриональном периоде. Впечатление об этих нейронах как об униполярных ложно, поэтому они и называются ложноодноотростчатыми. Дендрит чувствительного нейрона большой протяженности (более 1 м), выходит из спинномозгового узла в составе периферического нерва и может закончиться в любой ткани воспринимающим нервным аппаратом—рецептором. По дендриту чувствительные нервные импульсы передаются к телу его нейрона. Нейрит афферентного нейрона входит в спинной мозг и образует синапс—структуру, с помощью которой нервный импульс передается нейронам центральной нервной системы. Таким образом, вследствие большой длины дендрита псевдоуниполярные рецепторные нейроны имеют огромную протяженность. В результате воздействия внешней или внутренней среды чувствительные нейроциты генерируют нервные импульсы и передают их нейронам центральной нервной системы. Рецепторным нейроном начинается любая рефлекторная дуга.
Рис.2. Тигроид в двигательных нейронах
Препарат представляет поперечный срез спинного мозга, окрашенный основным красителем, толлундиновым синим, выявляющим базофильные структуры нейронов. Эти структуры находятся в основном в нейроплазме, поэтому окрашиваются преимущественно нейроны. Остальные тканевые элементы не окрашиваются, вследствие чего срез светлый и слабо заметен на предметном стекле. При малом увеличении надо найти в передних рогах спинного мозга крупные мультиполярные нейроциты, выделяющиеся голубой окраской на бледном фоне среза. Выбрав нервные клетки с хорошо окрашенной базофильной зернистостью, с отростками и ядрами, попавшими в срез, надо изучить их при большом увеличении. Двигательные нейроны имеют бедное хроматином пузырьковидное ядро (1) и 1—2 базофильных ядрышка (2). В нейроплазме видны глыбки базофильного вещества, названного в честь описавшего их нейрогистолога веществом Ниссля (3). Крупные, неправильной угловатой формы глыбки Ниссля располагаются наиболее плотно вокруг ядра. На периферии перикариона и в дендритах (4) они мельче, несколько вытянуты в длину и расположены реже. Базофильное вещество никогда не встречается в аксоплазме (5), а также в месте отхождения аксона от перикариона — аксональном бугорке (6). Особенность расположения глыбок Ниссля позволяет отличить аксон от дендритов. Крупные глыбки Ниссля придают перикариону пятнистый вид, отдаленно напоминающий рисунок тигровой шкуры. Поэтому вещество Ниссля нередко называют тигроидным веществом. Электронное микроскопирование нейроплазмы показывает, что глыбки базофильного вещества представляют участки, соответствующие гранулярной эндоплазматической сети и скоплениям свободных рибосом, РНК которых обусловливает базофилию этих участков цитоплазмы. Установлено, что в этих участках нейроплазмы происходит активный синтез белков. Хотя глыбки Ниссля специфичны для нейронов, электронно-микроскопическое исследование показывает, что специфическим является лишь своеобразное расположение конгломератов гранулярной цитоплазматической сети и рибосом в нейроплазме. Содержание, распределение и величина глыбок тигроидного вещества подвержены функциональным колебаниям. Повышение интенсивности специфической деятельности нейрона сопровождается увеличением его базофилии. В условиях перенапряжения, травмы, кислородного голодания и т. п. глыбки Ниссля распадаются на пылевидные частицы и исчезают. Эти морфологические изменения находятся в зависимости от характера поражения и дают основание судить о состоянии нейрона. Нормализация нейрона сопровождается восстановлением структуры базофильного вещества. Между нейронами видны косые сечения их отростков (7), мелкие голубые ядра глиоцитов (8), кровеносные сосуды (9).
Препарат 3. Спинной мозг кролика. Серебрение по Гросс-Бильшовскому.
Степень выраженности этого компонента нейроплазмы зависит от функционального состояния нейрона. Электронно-микроскопическое изучение нейроплазмы не обнаружило структур, соответствующих нейрофибриллам; обнаружены нейрофиламенты, или протонейрофибриллы, и микротрубочки, неразличимые при световой микроскопии. Считают, что они представляют комплексы белковых молекул, вследствие фиксации и обработки азотнокислым серебром склеиваются в пучки, на поверхности которых откладываются частицы восстановленного серебра. Протонейрофибриллы выполняют в нейроне, в особенности в его отростках, опорную функцию. Вокруг нейронов видны мелкие, округлые ядра (6) клеток нейроглии и тонкие глиальные волокна (7), образованные отростками глиоцитов. От глиальных волокон отличаются толстые, интенсивноокрашенные отростки нейронов, тела которых лежат в других отделах спинного мозга или за его пределами. В тонких прослойках рыхлой соединительной ткани проходят кровеносные сосуды (8). Белое вещество спинного мозга образовано миелиновыми нервными волокнами (9), проходящими в своем большинстве по его длинной оси. На поперечном срезе эти волокна имеют округлую форму, на косом — овальную. В центре мякотного волокна находится светлоокрашенный отросток нейрона — осевой цилиндр (10), окруженный миелиновой оболочкой (11). Пучки нервных волокон разделены глиальными перегородками, образованными отростками глиальных клеток серого вещества. Физиологическая регенерация нейронов проявляется постепенной сменяемостью ультраструктур нейроплазмы (внутриклеточная регенерация). При их повреждении репаративная регенерация возможна также на уровне ультраструктур нейроплазмы. В случае гибели нейроцитов репаративная регенерация не происходит.
Рис.4. Двигательные нейроны серого вещества спинного мозга
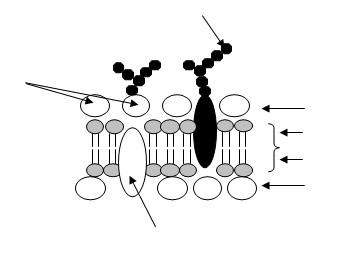
Синапсы.Нервные клетки своими отростками контактируют с другими нейронами или с клетками, не принадлежащими к нервной системе. Места таких контактов называют синапсами. По структуре и локализации синапсы подразделяются на три группы: межнейронные, рецепторно-нейрональные и нейроэффекторные. Межнейронные синапсы подразделяются на аксодендритические, аксосоматические и аксоаксональные.
Несмотря на разнообразие форм синапсов, в их строении наблюдаются общие черты. Конечные участки аксонов и дендритов в области синапса не имеют мякотной оболочки и расширены. В этой части аксона располагаются многочисленные митохондрии и синаптические пузырьки. Последние окружены цитоплазматическими мембранами. Диаметр пузырьков составляет 40—50 нм. Синаптические пузырьки содержат медиаторы (mediator — посредник) — биогенные амины типа ацетилхолина, адреналина, гамма-аминомасляной и глутаминовой кислот. В аминэргических синапсах, помимо синаптических пузырьков, имеются мелкие электронно-плотные гранулы, а в нейросекреторных нейронах — более крупные гранулы.
Нейрилеммы в области аксодендритических синапсов часто бывают утолщены за счет отложения электронно-плотного материала. В таких синапсах нейрилемма аксона называется пресинаптической мембраной, а контактирующая с ней нейрилемма дендрита—постсинаптической мембраной. Постсинаптической мембраной в иных синапсах может быть нейрилемма перикариона, аксона другой нервной клетки или плазмалемма клетки, не принадлежащей к нервной системе. Пресинаптической мембраной может быть нейрилемма аксона, плазмалемма рецепторной клетки либо эпителиальных и нейроглиальных клеток.
Между пресинаптической и постсинаптической мембранами имеется промежуток 12—30 нм, называемый синаптической щелью, которая является не чем иным, как межклеточным пространством. Содержимое синаптических пузырьков выделяется в синаптическую щель не путем экзоцитоза, а через поры диаметром 4—5 нм, имеющиеся в пресинаптической мембране. В таком синапсе передача нервного импульса осуществляется при помощи химического вещества — медиатора. Эти синапсы получили название синапсов с химической передачей. В некоторых синапсах синаптическая щель отсутствует и структурной основой его является плотный контакт. Возбуждение в таком синапсе может передаваться непосредственно, без участия медиатора, так как мембраны клеток соприкасаются. Эти синапсы называются синапсами с электрической передачей. В синапсах такого строения пресинаптическая мембрана также имеет поры, но они в 5 раз меньше, чем в синапсах с химической передачей возбуждения. Поры электрических синапсов являются межклеточными диффузионными каналами, соединяющими соприкасающиеся клетки.
Межнейронные синапсы, как свидетельствует их наименование, являются синапсами между нервными клетками. В том случае, если аксон одного нейрона контактирует с дендритом другого постсинаптического нейрона, то такие синапсы называются аксодендритическими. Эти синапсы очень распространены и имеют весьма разнообразное строение. Если аксон одного нейрона контактирует с перикарионом другого постсинаптического нейрона, то такой синапс называется аксосоматическим. В том случае, если аксон одного нейрона контактирует с аксоном другого постсинаптического нейрона, то такой синапс называется аксо-аксональным.
Межнейронные синапсы могут быть очень многочисленными. Подсчитано, что на поверхности перикариона и отростков одного пирамидального нейрона и коре больших полушарий головного мозга имеется около 10 тыс. синапсов.
Рецепторно-нейрональные (рецепторно-дендритные) синапсы, как свидетельствует их название, являются синапсами между рецепторными клетками, сходными с нейронами, специализированными эпителиальными, нейроглиальными клетками, с одной стороны и дендритами чувствительных нейронов —с другой. Примером синапсов такого типа у позвоночных являются синапсы вкусовых почек, боковой линии, внутреннего уха, кожи, соединительной ткани, причем рецепторы могут быть особо чувствительными ко многим формам энергии.
Нейроэффекторные (аксоэффекторные) синапсы,как свидетельствует их наименование, являются синапсами между аксоном двигательных (эффекторных) нейронов и клетками, не принадлежащими к нервной системе. У человека и млекопитающих животных хорошо изучены двигательные и секреторные нейроэффекторные синапсы, или эффекторные нервные окончания. Первые представляют собой синаптическое соединение между аксоном двигательного нейрона и поперечнополосатыми мышечными волокнами, поперечнополосатыми и гладкими мышечными клетками, а вторые—между аксоном двигательного нейрона и секреторными клетками. Кроме того, существуют многочисленные синапсы между аксоном эффекторного нейрона и другими клетками — жировыми, ресничными и пр.
Рис. 5. Центральный синапс:
1-аксон; 2-микротрубочки; 3-синаптический пузырек; 4-синаптическая щель; 5-дендрит; 6-рецептор для медиатора; 7-постсинаптическая мембрана; 8-пресинаптическая мембрана; 9-митохондрия
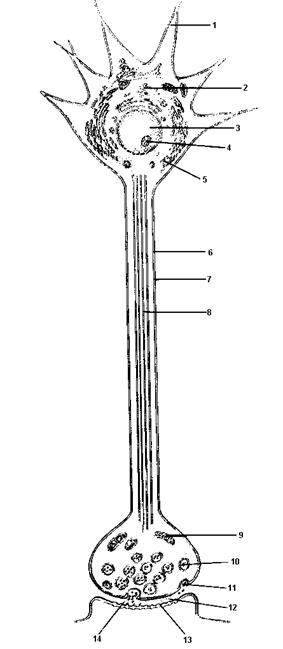
Рис.6. Внутреннее строение типичного нейрона:
1-дендрит; 2-гладкий эндоплазматический ретикулум; 3-ядро; 4-ядрышко; 5-шероховатый эндоплазматический ретикулум; 6-клеточная мембрана (нейролемма); 7-аксон; 8-микротрубочки; 9-митохондрия; 10-синаптический пузырек;11-обратное поглощение медиатора; 12-синапс; 13-рецепторы для медиатора; 14-выброс медиатора.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Нервная система человека в зависимости от онтофилогенетических и анатомо-физиологических особенностей подразделяется на центральную (головной и спинной мозг), периферическую (нервные корешки, узлы — ганглии, сплетения, черепные и спинномозговые нервы и т. д.), а также вегетативную (автономную), регулирующую деятельность органов.
Головной мозг в свою очередь подразделяется на несколько отделов. Конечный, или большой, мозг (telencephalon, cerebrum) состоит из полушарий большого мозга, долей, боковых желудочков, мозолистого тела, свода, базальных (подкорковых) ядер, внутренней капсулы. В большом мозге доли отделены друг от друга бороздами, из которых наиболее выражены центральная (роландова), разделяющая лобную и теменную доли, латеральная (сильвиева), отграничивающая лобную и теменную доли от височной, и теменно-затылочная, разделяющая теменную и затылочную доли. Благодаря
наличию борозд и извилин общая площадь коры большой мозга составляет около 2500 см 2 .
Промежуточный мозг (diencephalon) включает эпиталамус, задний таламус, метаталамус, передний таламус, гипоталамус, III желудочек. Средний мозг (mesencephalon) состоит из ножек мозга, черного вещества; покрышки, крыши и водопровода среднего мозга (сильвиева). Мозжечок (cerebellum) состоит из средней части (червя), двух боковых частей — полушарий и трех пар ножек. В глубине белого вещества мозжечка имеется несколько ядер — зубчатое, пробковидное, шаровидное и ядро шатра. Мост (pons) — задний мозг (metencephalon) включает ретикулярную формацию, вестибулярные ядра, ядра тройничного и лицевого нервов и другие образования. Продолговатый мозг (medulla oblongata) включает оливу, вестибулярные ядра, ядро подъязычного нерва, двойное ядро, перекрест пирамид, перекрест медиальных петель и др. В области заднего мозга и продолговатого мозга формируется IV желудочек, который включает ромбовидную ямку, латеральный карман, лицевой бугорок, крышу, сосудистые сплетения и т. д.
Спинной мозг
(medulla spinalis) состоит из передних, задних и боковых рогов, переднего, бокового и заднего канатиков, центрального канала, сегментов (шейных, грудных, поясничных, крестцовых, копчиковых) и других образований. Головной мозг взрослого человека весит около 1100—1200 г, спинной мозг — 30-40 г.
Центральная нервная система состоит из серого и белого вещества, а также ретикулярной формации, студенистого вещества и эпендимы. Серое вещество образовано скоплением тел нейронов и глии, белое — покрытыми миелиновой оболочкой отростками нейронов — аксонами и дендритами (пути и пучки). Серое вещество находится в коре большого мозга и мозжечка, в ядрах подкорковых узлов и ствола мозга, в передних и задних рогах спинного мозга.
Основной анатомо-функциональной структурой всей нервной системы является нервная к летка —нейрон, который состоит из тела и отростков — дендритов и аксона (рис. 6). В соответствии с законом Рамон-и-Кахаля (динамической поляризации нервной клетки) нервный импульс может распространяться в одном направлении—по нескольким дендритам к телу клетки и от него по единственному аксону к мышце, органу или дендриту следующего нейрона. Все нейроны разделяются на афферентные (рецепторные), передающие информацию от органов чувств в центральные отделы нервной системы, эфферентные (двигательные), посылающие импульсы (команды) к мышцам, органам и тканям, и вставочные (интернейроны), служащие для переработки и передачи импульсов. Первые клетки афферентных нейронов расположены обычно в вынесенных на периферию узлах (ганглиях), т. е. вне ЦНС. Клетки последующих нейронов, воспринимающих и передающих информацию в другие центры регуляции, сгруппированы в задних рогах спинного мозга, зрительном бугре и теменной доле мозга.
Эфферентные нейроны находятся главным образом в передних рогах спинного мозга, двигательных ядрах ствола и подкорковых узлов и в передней центральной извилине. Вставочные нейроны наиболее многочисленны и расцоложены во всех отделах ЦНС.
Аксоны большинства нервных клеток имеют миелиновую оболочку, которая через каждые 2—3 мм прерывается (так называемые перехваты Ранвье). Скорость проведения импульса в миелинизированных волокнах значительно большая (до 120 мс), чем в немиелинизированных (до 1 мс). В зависимости от некоторых особенностей строения аксонов все нервные клетки делятся на два типа. Клетки первого типа характеризуются длинными аксонами, которые отдают боковые ветви (коллатерали). Клетки второго типа имеют короткий аксон, делящийся вблизи его начала на концевые разветвления. К клеткам первого типа относятся двигательные (пирамидные) клетки в коре большого мозга, клетки передних рогов спинного мозга. К клеткам второго типа принадлежат вставочные нейроны. Особое строение % имеют афферентные (рецепторные) нейроны, имеющие два длинных отростка — аксон и аксоноподобно вытянутый дендрит.
Места стыков (контактов) между нейронами называют синапсами. Каждый из них состоит из нескольких элементов: пресинаптической мембраны, где происходит выделение медиатора передачи нервного возбуждения, синаптической щели и постсинаптической мембраны, обладающей избирательной чувствительностью к медиаторам нервного возбуждения. Передача импульсов в синапсах нервной системы осуществляется с помощью ряда веществ (ацетил-холина, норадреналина, серотонина, дофамина, ГАМК и да.). Одни синапсы (центральная нервная система, вегетативные узлы, окончания парасимпатических и двигательных нервов) возбуждаются холинергическими, другие (центральная нервная система, постганглионарные окончания симпатических нервов) норадренергическими медиаторами. Распространение их в ЦНС неравномерное (созданы специальные карты с указанием концентрации этих веществ в различных областях мозга).
В мембране нервных клеток есть рецепторы, которые взаимодействуют только с определенными, нужными клетке медиаторами и гормонами (принцип ключ—замок). Так, в лимбической системе, подкорковых узлах, ретикулярной формации, вестибулярном аппарате и на других уровнях ЦНС обнаружены специфические рецепторы — бензодиазепиновые (взаимодействуют с транквилизаторами), опиатные (взаимодействуют с наркотическими анальгетиками), дофа-минергические (взаимодействуют с дофамином) и др.
Нервные клетки, выполняющие различные функции, имеют известные различия в размерах и строении. Пирамидные клетки коры большого мозга имеют треугольную форму, клетки межпозвоночных узлов—овальную, клетки передних рогов спинного мозга — мультиполярную и т. д. Самые крупные размеры имеют находящиеся в двигательной области коры клетки Беца (до 120 μ ). Величина находящихся в афферентных областях коры клеток-зерен около 4 μ .
Основная функция нервной клетки — генерация возбуждения, восприятие и переработка импульсов (информативных сигналов), их проведение и передача на другие нервные клетки, орган или мышцу. Каждая нервная клетка содержит ядро и протоплазму, состоящие в свою очередь из множества специальных образований (органелл). В их число входят синаптические пузырьки, транспортирующие медиаторы— переносчики нервного возбуждения и ряд других. По программе, зашифрованной в молекулах ДНК в виде последовательности определенных химических реакций, в каждой нервной клетке ежеминутно происходит синтез и деградация сотен сложнейших соединений. Образующиеся в результате этих реакций на свободных и прикрепленных полисомах белковые молекулы обладают, вероятно, способностью к фиксации следов различных раздражений и являются таким образом важнейшей материальной основой нервно-психических процессов. При этом имеется цепь взаимосвязанных электрохимических явлений. Каждая нервная клетка покрыта мембранной оболочкой, избирательно проницаемой для ионов К + и Na + . В протоплазме нервной клетки по сравнению с внеклеточной жидкостью содержится приблизительно в 40—50 раз больше ионов К + , в 8—10 раз меньше ионов Na + и в 40—50 раз меньше ионов Сl — . Разность концентрации ионов и создает так называемый мембранный потенциал покоя (около 50—70 мВ).
Эта ионная асимметрия между вне- и внутриклеточной жидкостью поддерживается так называемым натрий-калиевым насосом, обеспечивающим удаление из протоплазмы
нервных клеток избыточных ионов Na + и поступление нужного количества ионов К + . Энергетическое обеспечение этого процесса осуществляется в основном за счет гликоли-тического расщепления глюкозы с образованием молекул АТФ. В ответ на всякое раздражение нейрон деполяризуется, т.е. возникает разность потенциалов между точкой раздражения (заряжается отрицательно) и окружающими участками
(заряжаются положительно). При этом проницаемость мембраны для ионов Na + резко возрастает и приток этих ионов внутрь .нервной клетки значительно повышается. Ионная асимметрия сдвигается в сторону Na + . Затем проницаемость мембраны для ионов Na + вновь понижается, а для ионов К + повышается и происходит реполяризация мембраны и т. д. Состояние распространяющейся деполяризации и реполяризации называется потенциалом действия (около 80—100 мВ). Он может перемещаться по нервному волокну и является началом нервного импульса.
Обычно по цепи из невронов в направлении от дендрина к телу нервной клетки и далее к ее аксону передаются залпы различных по частоте и продолжительности импульсов. Под их влиянием в синаптических бляшках происходит выделение медиаторов (ацетилхолина, симпатина и др.). Молекулы медиаторов взаимодействуют с рецепторами постсинаптической мембраны и в клетке открываются каналы для ионов К + и Na + . Возникающий поток ионов приводит нервную клетку в состояние возбуждения. Рождается электрический импульс, который передается следующему нейрону и т. д. В мякотных нервах распространение нервного импульса совершается скачкообразно от одного перехвата Ранвье к другому. Суммарная электрическая активность клеток мозга регистрируется с помощью электроэнцефалографа.
Глия вместе с сосудистой соединительной тканью представляет собой опорную ткань головного и спинного мозга, участвует в процессах обмена веществ в нервной системе, обладает трофической и барьерной функциями. По морфо-функциональным особенностям ее принято разделять на астроцитарную глию, олигодендроглию и микроглию (или мезоглию). Астроцитарная глия состоит из так называемых фиброзных и протоплазменных астроцитов — клеток, имеющих звездчатую или паукообразную форму. Фиброзные астроциты встречаются как в белом, так и в сером веществе мозга. Они образуют сетевидную строму (синцитий), волокна которой пронизывают всю паренхиму нервной ткани, крепко фиксируясь на кровеносных сосудах.
Протоплазменные астроциты имеют зернистую цитоплазму и более разветвленные и разнообразные по форме отростки. Встречаются преимущественно в сером веществе коры большого мозга. Они представляют собой разновидность сателитной невроглии и участвуют в заполнении пространства между элементами паренхимы. Олигодендроглия представлена мелкими округлой формы с немногочисленными отростками клетками, располагающимися около нервных клеток и сосудов. Ее роль заключается в образовании специального изолирующего материала — миелиновых оболочек отростков нервных клеток. Вне ЦНС эту функцию выполняют леммоциты (шванновские клетки). Микроглия состоит из клеток веретенообразной формы с длинными ветвящимися отростками и обладает большой спо собностью к фагоцитозу, образуя так называемые зернистые шары, или решетчатые клетки. При различных патологических состояниях глия вместе с сосудистой соединительной тканью подвергается гиперплазии и гипертрофии и активно участвует в освобождении нервной ткани от продуктов распада и процессах рубцевания.
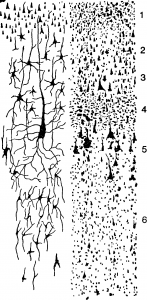
Сложное шестислойное строение имеет кора полушарий большого мозга . Первый (I) слой —молекулярный (lamina molecularis), II —наружный зернистый (lamina granih laris externa), III — пирамидный (lamina pyramidalis), IV —внутренний зернистый (lamina granulans interna), V — ганглионар ный (lamina ganglionaris) и VI — по диморфный (lamina multiformis). В афферентных зонах преобладают II и IV слои, в эфферентных — III и V.
В соответствии с особенностями морфологического строения предложено несколько вариантов классификации корковых полей. По Бродману, поверхность большого мозга разделяется на 11 областей и 52 поля: лобная область включает поля 8—12, 44—47; прецентральная область—поля 4, 6; постцентральная область — поля 1—3, 43; теменная область — поля 5, 7, 39, 40; височная область-поля 20-22, 36-38, 41, 42, 52; затылочная область—поля 17—19; островок — по ля 13—16; поясная область—поля 23—25, 31—33; ретроспленальная область—поля 26, 29, 30; область гиппокампа — поля 27, 28, 34, 35, 48; обонятельная область—поле 51 (рис.).
Нервная система человека содержит несколько сот миллиардов нервных и глиозных клеток. Только в коре полушарий большого мозга имеется около 15 млрд нервных и до 130 млрд глиозных клеток.
Статья на тему Морфология нервной системы
Читайте также:
