
Фоновая активность нервных центров при которой реализуется конкретная деятельность человека
показатели изменения функционирования физиологических систем организма: сердечной, дыхательной, эндокринной, двигательной и др.;
показатели изменений субъективных переживаний: появление чувства усталости, раздражительности, скуки или, наоборот, ощущение собранности, душевного подъема и т. д.;
показатели изменений в протекании основных психических процессов: восприятия, памяти, мышления и процессов в эмоционально-волевой сфере.
Способ оценки ФС через эффективность деятельности безусловно полезен для практики, так как позволяет предотвратить развитие у военнослужащего нежелательных ФС как для учебно-боевого процесса, так и для его личной безопасности. Однако метод оценки ФС через эффективность деятельности не обладает высокой чувствительностью к изменению ФС и в основном пригоден для выявления наиболее заметных и грубых его изменений.
Поэтому они предлагают оценивать ФС по комплексу взаимосвязанных физиологических реакций, а изменения ФС рассматривать как смену одного комплекса реакций другим. В свете таких представлений диагностика и идентификация ФС представляются как решение задачи распознавания многомерного вектора, компонентами которого являются различные физиологические показатели или реакции.
Учитывая значимость двух этих подходов, функциональное состояние следует рассматривать как особое психофизиологическое явление со своими закономерностями, которое заложено в архитектуре модулирующих функциональных систем и которое проявляется на биохимическом, физиологическом, поведенческом и психологическом (субъективном) уровнях.
Функциональное состояние организма - системная реакция организма, выражающаяся в виде интегрального динамического комплекса наличных характеристик тех функций и качеств индивида, которые прямо или косвенно обусловливают выполнение деятельности.
Подчеркнем основные моменты в этом определении:
системность реакции;
интегральный характер комплекса изменений;
непосредственная связь с эффективностью деятельности.
Общие положения теории функциональных состояний.
Решающую роль в изучении теории функциональных состояний сыграло открытие И.М. Сеченовым (1908) явления центрального торможения. Далее - И.П. Павлов выдвинул идею о целевой функции, определяющей целостность ответа организма.
Адекватное определение понятия ФС должно разрабатываться на основе системных представлений о формировании целостного ответа организма. Это предполагает, что в основе ФС лежит формирование своеобразной системы.
Система - совокупность взаимодействующих между собой элементарных структур или процессов, объединенных в целое выполнением общей функции, которую не может осуществить ни один из ее компонентов в отдельности.
Описывая ФС как целостную реакцию организма, в качестве основных структур или звеньев системы выделяются функции и процессы разных уровней: физиологического, психологического, поведенческого.
на физиологическом уровне структуры обеспечивающие двигательные и вегетативные компоненты состояния;
на психологическом уровне - характеристики основных психических процессов (восприятие, внимание, память, мышление, эмоционально-волевые процессы);
на поведенческом уровне - скоростные показатели и показатели точности, особенности двигательного и речевого сопровождения деятельности.
В ФС различают 5 групп компонентов:
Энергетические компоненты, то есть функции, обеспечивающие требуемый уровень энерготрат. Сюда входят практически все вегетативные системы организма.
Сенсорные компоненты, которые характеризуют наличие возможности приема и первичной обработки поступающей информации. Сюда входит вся последовательность процессов обработки информации. На психологическом уровне сенсорные компоненты изучаются в форме процессов ощущения и восприятия.
Информационные компоненты, обеспечивают дальнейшую обработку поступившей информации и принятие решения.
Эффекторные компоненты - компоненты деятельности, отвечающие за реализацию принятых решений в поведенческих актах.
Активационные компоненты деятельности. Характеризуют способность человека к реализации имеющихся у него качеств и личностных свойств. В эту группу входит целый ряд физиологических и психологических показателей: особенности нервной и гуморальной регуляции, уровень внимания, установки человека, специфика его мотивационной сферы.
Вам хорошо известно, что нейроны могут находиться в нескольких состояниях: поляризованы, частично или полностью деполяризованы и гиперполяризованы. От того, каков заряд мембраны нейрона относительно мембранного потенциала покоя зависит его возбудимость. В нервных центрах нейроны организованы в группы, и состояние нервного центра – его способность к возбуждению или торможению уже зависит не только от состояния отдельных нейронов, но и от взаимодействия нейронов внутри таких групп. Нам знакомо явление иррадиации – распространение возбуждения в дивергентных нейронных сетях. Явление суммации и конвергенции возбуждения в конвергентных сетях, продолжительная реверберация импульса в локальных сетях. Возможно модулирование процесса возбуждения тормозным процессом.
Центральная нервная система, и головной мозг в частности, представляют собой сложный комплекс различных нервных центров, каждый из которых во-первых, выполняет одну или несколько определенных функций, а во-вторых, объединен сложной системой восходящих и нисходящих связей с другими центрами и корой головного мозга. Понятно, что способность головного мозга осуществлять то или иное поведение, выполнять функции запоминания и извлечения памяти, мышления, концентрировать внимание изменяется и зависит и от состояния отдельных нейронов, и от состояния и взаимного влияния нервных центров.
Наиболее часто функциональное состояние мозга определяют как фоновую активность нервных центров, при которой реализуется та или иная конкретная деятельность человека.
Выбор центров и включение их в функциональную систему поведения можно считать качественной стороной функционального состояния мозга. Мы немного погрешим против истины в пользу ясности изложения, если скажем, что есть и количественная сторона функционального состояния. Дело в том, что в головном мозге есть и неспецифические системы, которые могут изменить активность головного мозга в целом, и его коры вне зависимости от вида деятельности, а просто по факту наличия или отсутствия какой-либо деятельности.
К таким неспецифическим системам относятся: желатинозная субстанция спинного мозга, ретикулярная формация ствола мозга, неспецифические ядра таламуса, базальные отделы переднего мозга и новая кора. Общий принцип функционирования этих неспецифических структур заключается в том что, получив возбуждающую информацию, они активируют все вышележащие структуры мозга.
Важнейшей частью общей неспецифической регуляторной системы является ретикулярная формация продолговатого мозга, моста и среднего мозга. Активирующая роль ретикулярной формации доказана экспериментально: стимуляция электрическими импульсами приводит к генерализованной активации коры, десинхронизации биопотенциалов и поведенческому возбуждению; разрушение способствует глубокому сну и блокирует пробуждение, которое вызвано соматосенсорной стимуляцией. Активация осуществляется несколькими основными группами ядер. Серотонинергические нейроны сосредоточены в ядрах шва,которые образуют непрерывную структуру от каудального отдела продолговатого мозга к покрышке моста и среднего мозга. Гигантоклеточное ядро ретикулярной формации, которое получает информацию из спинного мозга о болевом воздействии, в этом ядре обнаружено большое количество опиатных рецепторов и эндогенных пептидов, это компоненты в антиболевой системы. Ядро одиночного пучка принимает информацию от барорецепторов сосудов, рецепторов растяжения легких, осморецептрово и передает сигналы в гипоталамус. Основной медиатор здесь – норадреналин. Наиболее крупное объединение норадренергических нейронов обнаружено в голубом пятне моста. Нейроны этой области получают информацию от многих областей мозга. Вентральное поле покрышкиобеспечивает дофаминергическую иннервацию структур гипоталамуса, подкорковых ядер и новой коры. Стимуляция этой области вызывает увеличение двигательной активности.
Основные медиаторные системы ретикулярной формации
| ядра шва | серотонин |
| голубое пятно | НА |
| гигантоклеточное ядро | АХ |
| вентральное поле покрышки | дофамин |
В экспериментах установлено, что ретикулярная формация оказывает активирующее влияние практически на все области коры головного мозга. Однако, есть неспецифическая система, которая работает более тонко: активирует определенные зоны коры в соответствии с потоком сенсорной информации. Кроме того, эта система обладает не только активирующим, но и тормозным влиянием на кору. Такой системой являются неспецифические ядра таламуса. Сюда приходит информация от соматической, висцеральной, слуховой, зрительной систем, а так же от ретикулярной формации ствола мозга и гипоталамуса. Удалось экспериментально доказать, что стимуляция таламуса низкой частотой в режиме пачечной активности (группа импульсов) приводит к торможению коры, а повышение частоты стимуляции одиночными импульсами возбуждает активирующую систему таламуса и передается коре головного мозга. Свойства неспецифических ядер таламуса возбуждаться в ответ на пришедший поток афферентных импульсов и передавать это активирующее влияние коре связывают с наличием в таламических ядрах специальных нейронов, которые получили название нейронов-пейсмекеров. Эти нейроны очень легко вовлекаются в ритмическую активность, буквально одного импульса возбуждения достаточно для того, чтобы такой нейрон начал генерировать ритмичные импульсы, причем длительность такой ритмичной активности весьма велика.
В коре головного мозга тоже есть неспецифические системы, которые могут активировать или тормозить работу коры. Следует отметить, что как снижение, так и увеличение функционального состояния ухудшает аналитические возможности мозга и затрудняет принятие адекватных решений. Оказалось, что раздражение поясной извилины у обезьян вызывает такую же реакцию, как и раздражение ретикулярной формации. Однако порог активации коры примерно в два раза выше, чем ретикулярной формации. Порог реакции для структур неспецифического таламуса занимает промежуточное положение.
Таким образом, любое раздражение низкопороговых нейронов ретикулярной формации потоком афферентных импульсов повышает активность всех структур головного мозга и задает их функциональное состояние. Усиление потока импульсов и анализ информации от анализаторов приводит к возбуждению более высокопороговых нейронов таламуса. Передача эстафеты активирующих влияний с уровня ретикулярной формации на таламичекий уровень означает переход от генерализованной активации коры к локальной, в соответствии с потоком информации от анализаторов готовится к работе уже определенная область коры.

Рисунок2 Схема активации мозга
Во второй половине нашего столетия прямое изучение нейронов, вовлеченных в регуляцию сна-бодрствования, показало, что нормальная работа таламо-кортикальной системы мозга, обеспечивающая сознательную деятельность человека в бодрствовании, возможна только при участии определенных подкорковых, так называемых активирующих, структур. Благодаря их действиям в бодрствовании мембрана большинства кортикальных нейронов деполяризована на 10-15 мВ по сравнению с потенциалом покоя - (65-70) мВ. Только в состоянии этой тонической деполяризации нейроны способны обрабатывать информацию и отвечать на сигналы, приходящие к ним от других нервных клеток (рецепторных и внутримозговых). Как сейчас ясно, таких систем тонической деполяризации, или активации мозга (условно "центров бодрствования"), несколько - вероятно, пять или шесть. Располагаются они на всех уровнях мозговой оси: в ретикулярной формации ствола, в области голубого пятна и дорзальных ядер шва, в заднем гипоталамусе и базальных ядрах переднего мозга. Нейроны этих отделов выделяют медиаторы - глутаминовую и аспарагиновую кислоты, ацетилхолин, норадреналин, серотонин и гистамин, активность которых регулируют многочисленные пептиды, находящиеся с ними в одних и тех же везикулах. У человека нарушение деятельности любой из этих систем не компенсируется за счет других, несовместимо с сознанием и приводит к коме.
Проявление функционального состояния.
Выделяют три основные группы физиологических реакций, по которым судят об изменении функционального состояния
1. Двигательная активность: интенсивность и количество различных движений, уровень мышечного тонуса, определенная поза.
2. Вегетативные показатели: частота сердечных сокращений, частота дыхания, артериальное давление, скорость кровотока, кожно-гальваническая реакция (изменение электрического сопротивления кожи).
3. Электроэнецефалография: реакции изменения основных ритмов.
Каждому функциональному состоянию соответствует определенный тип инстинктивного поведения. В таблице суммированы представления о соотношении функциональных состояний, инстинктивного поведения и субъективного переживания. Становится ясно, что биологическое значение изменения функционального состояния мозга заключается в настройке его работы на определенное поведение в соответствии с удовлетворением конкретной потребности. Поведение же определяется взаимодействием врожденных (инстинкты) и приобретенных форм.
| Уровень ретикулярной активации | Инстинктивное поведение | Субъективное переживание |
| Бодрствование | Оборонительно-агрессивное поведение | Эмоции |
| Сексуальное инстинктивное поведение | Настороженность | |
| Пищевое инстинктивное поведение | Спокойное бодрствование | |
| Критический уровень для осознанного поведения | Подготовка ко сну | Дремота |
| Сон | Сновидения | |
| Кома |
Электроэнцефалография
Электроэнцефалография (ЭЭГ) это метод исследования функционального состояния головного мозга, основанный на графической регистрации его биопотенциалов.
По современным представлениям, ЭЭГ, регистрируемая через неповрежденные покровы головы с помощью наложенных электродов, является суммарной, усредненной биоэлектрической активностью мозга. Эта активность обусловлена синхронной активностью множества нейронов. Суммарная ЭЭГ представляет собой организованный колебательный процесс, в котором визуально, а особенно четко при машинном анализе, можно выделить регулярные частотные составляющие.
ЭЭГ характеризуется такими показателями как частота, амплитуда и фаза колебаний. В ЭЭГ выделяют следующие основные ритмы:
Основные ритмы ЭЭГ человека
| Ритм | Частота (Гц) | Амплитуда (микровольт) |
| Дельта- ритм ( d ) | 1 – 3 | 40 – 70 мкв |
| Тета -ритм (q) | 4 - 7 | 40 – 70 мкв |
| Альфа (a) | 8 – 13 | 40 – 70 мкв |
| Бета- ритм (b) низкий высокий | 14 – 20 20 - 30 | 15 – 20 мкв |
| Гамма –ритм (g) | Выше 30 | 8 – 12 мкв |
Наиболее характерным является альфа ритм. Наибольшую выраженность альфа-ритм имеет в затылочной, затылочно-височной и затылочно-теменной областях мозга. У 75-90% здоровых людей этот ритм является доминирующим. Альфа ритм регистрируется в состоянии покоя при закрытых глазах и расслаблении мышц.
Открывание глаз во время записи ЭЭГ, воздействие света вызывают четкую реакцию десинхронизации альфа-ритма. В настоящее время эту реакцию называют реакцией активации. Наличие или отсутствия реакции активации, ее выраженность и скорость восстановления альфа-ритма используют и в диагностических целях, и в экспертной практике как критерий реактивности ЦНС.

Рисунок 3 Основные ритмы электроэнцефалограммы человека.
Бета-ритм – частота выше, а амплитуда меньше, чем у альфа-ритма. Наиболее выражен этот ритм в центральных и лобных областях коры. Бета-ритм отражает активированное состояние подкорковых образований и коры головного мозга.
Тета-ритм равен по амплитуде альфа-ритму, но частота его ниже. Тета-ритм отражает активность срединно-стволовых образований головного мозга, этот ритм изменяется по мере созревания и окончательного формирования корково-стволовых связей. Изменения проявляются в снижении амплитуды ритма. При поражении стволовых структур мозга амплитуда ритма увеличивается до 400-500 мкв, что отражает состояние чрезмерного и стойкого повышения активности структур ствола мозга и снижения активности коры.
Дельта ритм – ритм с наименьшей частотой. Появление на ЭЭГ волн дельта-ритма указывает на снижение уровня функциональной активности коры, например, при дремоте, утомлении. В норме дельта волны регистрируются кратковременно и диффузно во всех отведениях. При появлении раздражителя любой модальности дельта-ритм исчезает и сменяется альфа-ритмом.
Гамма-ритмом называют колебания с частотой от 30 до 100 Гц и амплитудой 8 -12 мкв. Закономерные изменения гамма-ритма отмечаются при различных видах умственной деятельности. При обычной, чернильной записи ЭЭГ этот ритм выявить не удается. Для этого нужна специальная обработка ЭЭГ записи.
Итак, основным ритмом у здорового человека в состоянии бодрствования, при расслабленной мускулатуре и закрытых глазах является альфа-ритм. При переходе к более активному бодрствованию – открытые глаза, поток афферентной информации любой модальности приводят к десинхронизации ритма и переходу к бета-ритму. Синхронизация ритма – снижение частоты и повышение амплитуды характеризуют переход ко сну. Однако эти соотношения между уровнем функциональной активности и ритмом ЭЭГ не должны трактоваться упрощенно. Дело в том, что вся психическая деятельность человека прямого отражения в волнах ЭЭГ не находит. Необходимы дальнейшие комплексные электрофизиологические и психофизиологические исследования для раскрытия электрофизиологической картины активного бодрствования.


Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.
Рассматриваемые ниже свойства нервных центров связаны с некоторыми особенностями распространения возбуждения в ЦНС, особыми свойствами химических синапсов и свойствами мембран нервных клеток. Основными свойствами нервных центров являются следующие.
А. Инерционность — сравнительно медленное возникновение возбуждения всего комплекса нейронов центра при поступлении к нему импульсов и медленное исчезновение возбуждения нейронов центра после прекращения входной импульсации. Инерционность центров связана с суммацией возбуждения и последействием.
Явление суммации возбуждения в ЦНС открыл И. М. Сеченов (1868) в опыте на лягушке: раздражение конечности лягушки слабыми редкими импульсами не вызывает реакции, а более частые раздражения такими же слабыми импульсами сопровождаются ответной реакцией - лягушка совершает прыжок. Различают временную (последовательную) и пространственную суммацию (рис. 5.9).
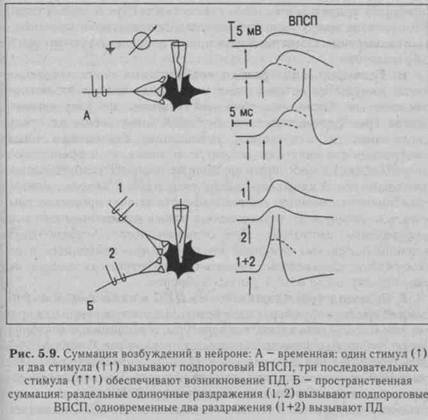
Последействие — это продолжение возбуждения нервного центра после прекращения поступления к нему импульсов по афферентным нервным путям. Основной причиной последействия является циркуляция возбуждения по замкнутым нейронным цепям (см. рис. 5.8), которая может продолжаться минуты и даже часы.
Б. Фоновая активность нервных центров (тонус) объясняется: 1) спонтанной активностью нейронов ЦНС; 2) гуморальными влияниями биологически активных веществ (метаболиты, гормоны, медиаторы и др.), циркулирующих в крови и влияющих на возбудимость нейронов; 3) афферентной импульсацией от различных рефлексогенных зон; 4) суммацией миниатюрных потенциалов, возникающих в результате спонтанного выделения квантов медиатора из аксонов, образующих синапсы на нейронах; 5) циркуляцией возбуждения в ЦНС. Значениефоновой активности нервных центров заключается в обеспечении некоторого
исходного уровня деятельного состояния центра и эффекторов. Этот уровень может увеличиваться или уменьшаться в зависимости от колебаний суммарной активности нейронов нервного центра-регулятора.
В. Трансформация ритма возбуждения — это изменение числа импульсов, возникающих в нейронах центра на выходе относительно числа импульсов, поступающих на вход данного центра. Трансформация ритма возбуждения возможна как в сторону увеличения, так и в сторону уменьшения. Увеличению числа импульсов, возникающих в центре в ответ на афферентную импульсацию, способствуют иррадиация процесса возбуждения и последействие. Уменьшение числа импульсов в нервном центре объясняется снижением его возбудимости за счет процессов пре-и постсинаптического торможения, а также избыточным потоком афферентных импульсов. При большом потоке афферентных влияний, когда уже все нейроны центра или нейронного пула возбуждены, дальнейшее увеличение афферентных входов не увеличивает число возбужденных нейронов.
Г. Большая чувствительность ЦНС к изменениям внутренней среды, например, к изменению содержания глюкозы в крови, газового состава крови, температуры, к вводимым с лечебной целью различным фармакологическим препаратам. В первую очередь реагируют синапсы нейронов. Особенно чувствительны нейроны ЦНС к недостатку глюкозы и кислорода. При снижении содержания глюкозы в 2 раза ниже нормы (до 50% от нормы) могут возникнуть судороги. Тяжелые последствия для ЦНС вызывает недостаток кислорода в крови. Прекращение кровотока всего лишь на 10 с приводит к очевидным нарушениям функций мозга, человек теряет сознание. Прекращение кровотока на 8-12 мин вызывает необратимые нарушения деятельности мозга - погибают многие нейроны, в первую очередь корковые, что ведет к тяжелым последствиям.
Д. Пластичность нервных центров — способность нервных элементов к перестройке функциональных свойств. Основные проявления пластичности следующие.
1. Синаптическое облегчение - это улучшение проведения в синапсах после короткого раздражения афферентных путей. Степень выраженности облегчения возрастает с увеличением частоты импульсов, оно максимально, когда импульсы поступают с интервалом в несколько миллисекунд.
Длительность синаптического облегчения зависит от свойств синапса и характера раздражения - после одиночных стимулов оно невелико, после раздражающей серии облегчение в ЦНС может
продолжаться от нескольких минут до нескольких часов. По-видимому, главной причиной возникновения синаптического облегчения является накопление Са 2+ в пресинаптических окончаниях, поскольку Са 2+ , который входит в нервное окончание во время ПД, накапливается там, так как ионная помпа не успевает выводить его из нервного окончания. Соответственно увеличивается высвобождение медиатора при возникновении каждого импульса в нервном окончании, возрастает ВПСП. Кроме того, при частом использовании синапсов ускоряется синтез рецепторов и медиатора и ускоряется мобилизация пузырьков медиатора, напротив, при редком использовании синапсов синтез медиаторов уменьшается - важнейшее свойство ЦНС. Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах. Значение синаптического облегчения заключается в том, что оно создает предпосылки улучшения процессов переработки информации на нейронах нервных центров, что крайне важно, например, для обучения в ходе выработки двигательных навыков, условных рефлексов.
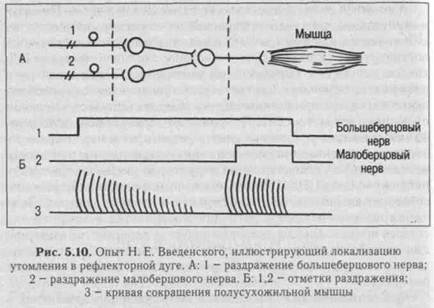
2. Синаптическая депрессия - это ухудшение проведения в синапсах в результате длительной посылки импульсов, например, при длительном раздражении афферентного нерва (утомляемость центра). Утомляемость нервных центров продемонстрировал Н. Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения п. тлЫаНз и п. регопеиз. В этом случае ритмическое раздражение одного нерва вызывает ритмические сокращения мышцы, приводящие к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги (рис. 5.10). Ослабление реакции центра на афферентные импульсы выражается в снижении постси-наптических потенциалов. Оно объясняется расходованием медиатора, накоплением метаболитов, в частности, закислением среды при длительном проведении возбуждения по одним и тем же нейронным цепям.
3. Доминанта - стойкий господствующий очаг возбуждения в ЦНС, подчиняющий себе функции других нервных центров. Доминанта - это более стойкий феномен облегчения. Явление доминанты открыл А. А. Ухтомский (1923) в опытах с раздражением двигательных зон большого мозга и наблюдением сгибания конечности животного. Как выяснилось, если раздражать корковую двигательную зону на фоне избыточного повышения возбудимости другого
нервного центра, сгибания конечности может не произойти. Вместо^ сгибания конечности раздражение двигательной зоны вызывает реакцию тех эффекторов, деятельность которых контролируется господствующим, т. е. доминирующим в данный момент в ЦНС, нервным центром.
Значение доминантного очага возбуждения в ЦНС заключается в том, что на его базе формируется конкретная приспособительная деятельность, ориентированная на достижение полезных результатов, необходимых для устранения причин, поддерживающих тот или иной нервный центр в доминантном состоянии. Например, на базе доминантного состояния центра голода реализуется пищедобывательное поведение, на базе доминантного состояния центра жажды запускается поведение, направленное на поиск воды. Успешное завершение данных поведенческих актов в конечном итоге устраняет физиологические причины доминантного состояния центров голода или жажды. Доминантное состояние центров ЦНС обеспечивает автоматизированное выполнение двигательных реакций.
Функциональное состояние определяется общим уровнем активации организма. От функционального состояния зависит успешность школьника и работника, внимание водителя на дорожной трассе и кассира, считающего сдачу. Но как создается фоновая активность организма? Какие структуры мозга отвечают за этот процесс? Как мы можем пронаблюдать за функциональными состояниями на ЭЭГ? Попытаемся разобраться.
Функциональное состояние часто определяется как фоновая активность нервных центров, при которых реализуется любая деятельность человека. Функциональное состояние может меняться в зависимости от времени суток, сезона года, выпитого нами кофе, принятого амфетамина и других факторов.

Функциональное состояние проявляется в том, насколько бодро мы себя чувствуем и насколько качественно выполняем нашу работу. Оно зависит от времени суток – у нас бывают пики и спады продуктивности. Так, например, время с 16:00 до 17:00 считается периодом, когда человек совершает наибольшее количество ошибок. Именно в этот временной диапазон было зафиксировано больше всего аварий и несчастных случаев, произошедших из-за человеческого фактора. Что же касается пика активности, то у большинства он приходится на десять часов утра. В это время люди мобилизуются и работают наиболее продуктивно.
Однако нельзя сказать, что пики и спады активности одинаково работают для всех людей – здесь нужно учитывать индивидуальные особенности человека. В зависимости от того, в какое время суток нам удобнее всего работать, выделяют различные хронотипы. Знание своего хронотипа может быть полезно для людей, которые хотят быть продуктивными. В настоящее время наиболее известным тестом для определения хронотипа является тест Хорна Остберга, который поможет определить время сна и бодрствования психологически удобное для конкретной личности.
Наше функциональное состояние регулируют модулирующие системы головного мозга. В их состав входят ретикулярная формация и лимбическая система.
Особое внимание при исследовании модулирующих систем головного мозга было оказано ретикулярной формации и таламусу – одной из частей лимбической системы. Так, при высокочастотной стимуляции ретикулярной формации электрическим током испытуемым повышали уровень бодрствования, если же стимуляция была низкочастотная, то эффект оказывался противоположным.

Как наблюдают за функциональными состояниями психофизиологи?
Для определения функционального состояния используют следующие показатели:
- Двигательные – уровень двигательной активности, фоновый мышечный тонус.
- Вегетативные – частота дыхания, пульс, кожно-гальванический рефлекс.
- ЭЭГ показатели – наиболее часто применяется в клинике и в исследованиях.
Поговорим подробнее о главном психофизиологическом методе измерения ФС – электроэнцефалограмме.
ЭЭГ регистрирует активность нейронов. Разные уровни этой активности называют ритмами. Эта ритмическая активность создаёт условия для взаимодействия удаленных нейронов при обучении и других задачах. Каждый ритм имеет свой диапазон частот.
Какими же они бывают?
· Дельта-ритм фиксируется во время глубокого сна. При стимуляции этого ритма может пропадать ощущение боли. Частота – 1-4 Гц.
· Тета-ритм наблюдается во время медитации или сна. Состояние спокойствия и полного расслабления. Частота – 4-8 Гц
Дельта- и Тета-ритмы связаны с падением уровня функционального состояния.
· Альфа-ритм появляется, когда мы расслабляемся, например, во время просмотра телевизор или прослушивания фоновой музыки. Чилл-ритм. Частота – 8-13 Гц.
· Мю-ритм регистрируется в роландичекой области головного мозга. Никакого чилла, мю-ритм связан с умственными нагрузками. Частота – 8-13 Гц.
· Каппа-ритм зафиксирован в височной области при умственной деятельности. Частота – 8-13 Гц.
· Бета-ритм регистрируется в области передних центральных извилин. При бета-ритме мы более активны, нежели при альфа. Мы готовы осознанно воспринимать и перерабатывать информацию. Появляется, когда мы читаем серьёзную литературу, отвечаем на вопросы, ведём лекцию или участвуем в дискуссии. Стимулируется кофе, энергетиками, чтением увлекательной литературы или никотином. Частота – 14-40 Гц.
· Гамма-ритм выражен при активном обучении. Когда до экзамена два дня, а материала много, то, вероятнее всего, в вашей комнате от стен будут отражаться именно гамма-волны. С гамма-ритмом связан один интересный факт. Он редкий гость в нашей повседневной жизни, и лишь йоги, регулярно практикующие медитацию, научились вызывать его самостоятельно. Однако некоторые ученые ставят под сомнение существование гамма-ритма, так как его сложно отделить от электрической активности мышц. Его частота – 30-60 Гц, а по некоторым данным может достигать и больших значений. 
Как функциональные состояния определяются нашей физиологией?
Как мы уже сказали, за поддержание фоновой активности ответственны модулирующие системы. Важность этого влияния на функциональное состояние были показаны в экспериментах профессора Зингера. Он обнаружил, что свойства нейронов, реагирующих на отдельные зрительные признаки, зависят от сохранности стволовой и таламической активирующих систем мозга. Ученый получил такие данные, мучая глазки и мозг маленьких котят.
Суть эксперимента была следующей: первой группе котиков разрушали таламус, а второй группе модулирующую систему оставляли в сохранности. В период становления зрения зверькам закрывали один глаз, а другой оставляли открытым. Оказалось, что у котят с разрушенным таламусом нейроны сохранили свои бинокулярные свойства и реагировали на стимуляцию обоих глаз, несмотря на процедуру депривации. У котят с таламусом, наоборот, наблюдались нарушения зрения.
Эти опыты показывают, что пластические изменения в коре, наступающие под влиянием индивидуального опыта, требуют как зрительной стимуляции, так и функционирования структур, которые модулируют кортикальную возбудимость и определяют внимание.
При изучении функциональных состояний, как и при изучении чего-либо в области психофизиологии, нужно ставить эксперименты, и порой они не совсем гуманны. Так, для определения роли уровня функционального состояния на выполнение той или иной деятельности, учёным Котляром были взяты в качестве подопытных кроликов – кролики.
Эксперименты Котляра – апгрейд Павлова. Они очень похожи на эксперименты с собаками и выработкой условного рефлекса, только теперь мы перешли на новый уровень – уровень нейронов головного мозга.
Учёный вырабатывал условные рефлексы у нейронов гиппокампа кролика при сочетании звукового стимула с электрокожным раздражением. Примерно у 40% нейронов гиппокампа после обучения условный стимул приобретал способность вызывать ту реакцию, которую до обучения вызывало лишь электрокожное раздражение. Параллельно со становлением этого условного рефлекса ученый наблюдал у обучаемого нейрона повышенную активность в межстимульные интервалы. Этот эффект был назван ассоциативным тоническим ответом. Как выяснилось, ассоциативный тонический ответ определяет успешность выработки условно-рефлекторной связи и уровень активности обучаемого нейронов для каждой деятельности.
Кроме того, что Котляр подтвердил значимость фоновой активности нейронов для формирования условного рефлекса, он также выявил, что во всех наших активностях участвуют свои специфические структуры головного мозга. Механизмы регуляции функционального состояния необходимы для любой деятельности. Получается, что от функционирования модулирующих систем мозга, зависит не только обучение, но осуществление врожденного поведения.
Как полученные экспериментальные данные применяются на практике и какие выводы мы можем извлечь для себя?
Как известно, в школах не все ученики имеют одинаковый уровень развития, внимания, запоминания. Эксперименты Котляра показали, что такие дети не совсем безнадежны, и их возможно обучить математике, физике, языкознанию – достаточно лишь повысить их функциональное состояние и предложить им индивидуальный подход к обучению.
Для повышения функционального состояния можно использовать несложные приёмы. Например, прослушивание ритмичной музыки, нормализация питания, в случае необходимости принятие витаминов. Основным источником повышения оптимизации работы в случае психических заболеваний является психофармакология.
Какой же вывод вы можете вынести из этой статьи? Без оптимального функционального состояния мы не можем быть продуктивными. Если организм вымотан, истощён, то мы будем допускать ошибки, а поступающая информация, будет пролетать мимо ушей, а главное – мимо нашего мозга.
Поэтому для эффективной работы выберите для себя нужное время, хорошенько отдохните, поставьте любимую пластинку или вовсе отрубите все звуки вокруг.
Читайте также:
