
Функциональная классификация нервных волокон по эрлангеру и гассеру
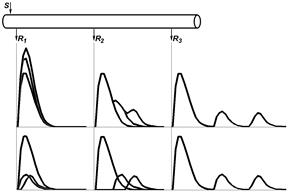
В 1939 г. американские физиологи Джозеф Эрлангер и Герберт С.Гассер зарегистрировали токи действия от целого нервного ствола седалищного нерва лягушки на разных расстояниях от стимулирующего электрода (рис. 210041905).
Было установлено, что регистрируемый суммарный потенциал имеет ряд пиков, которые были обозначены буквами латинского алфавита A, B, C (рис. , I). Пик A имел дополнительные пики, помеченные греческими буквами α, β, γ, δ (рис. , II). В 1944 г. работа Дж.Эрлангера и Г.С.Гассера была оценена присуждением Нобелевской премии.
Нервы у позвоночных состоят из трех основных групп волокон (А, В и С), различающихся по степени миелизации, диаметру волокна, длительности пика ПД (скорости развития ПД), электровозбудимости, его компенсации и скорости проведения (все эти показатели в ряду А — В — С падают).
Группа А включает наиболее толстые хорошо миелинизированные моторные и чувствительные волокна; группа В — слабомиелинизированные, преганглионарные волокна автономной нервной системы; группа С - немиелинизированные, постганглионарные (симпатические) волокна.
В группе А в ряду a, b, g, d названные показатели тоже падают. Соотношения свойств этих групп волокон демонстрируются в табл. .
Необходимо заметить, что указанные соотношения порогов электрического раздражения групп волокон не отражают точного соотношения электровозбудимости их мембран. Относительно высокие пороги тонких волокон при их раздражении в нервном стволе определяются в основном тем обстоятельством, что тонкие волокна по сравнению с толстыми обладают более высоким входным сопротивлением. В них входит такая малая часть раздражающего тока, что при пороговой силе для Аa-волокон она совершенно недостаточна для создания на мембране более тонких волокон сколько-нибудь существенной деполяризации. По этой же причине (высокое RI) отводимые от ствола (внеклеточно) ПД тонких волокон предстают значительно меньшими, чем ПД толстых волокон.

Рис. . Составные части потенциала действия смешанного нерва.
I – при относительно медленной скорости записи.
II – при относительно высокой скорости записи.
Объяснение в тексте. По оси абсцисс – время, по оси ординат амплитуда составного потенциала в мВ.
Суммарная электрическая активность нерва создается его волокнами, каждое из которых генерирует свой стандартный по амплитуде и временным параметрам ПД, распространяющийся в обе стороны от точки, к которой приложено раздражение. Суммарный электрический сигнал нерва зависит от числа активных волокон, синхронности их активности, способа отведения и других обстоятельств.
При дальнейшем увеличении силы стимула этот ПД несколько удлиняется во времени. Все изменения амплитуды и длительности пика ПД нерва при усилении стимула определяются ростом числа активных волокон, подключением к низкопороговым и быстрым А(альфа)-волокнам более высокопороговых медленных бета-, гамма-, дельта-волокон группы А, затем В- и, наконец, С-группы.
| Группы волокон (по Эрлангеру и Гассеру) | Диаметр, мкм | Скорость проведения, м/с |
| Aa | 13 - 22 | 70 - 120 |
| Ab | 8 -13 | 40 - 70 |
| Ag | 4 - 8 | 15 – 40 |
| Ad | 1 – 4 | 5 – 15 |
| B | 1 - 3 | 3 – 14 |
| C | 0,5 – 1,0 | 0,5 - 2 |

Таблица . Классификация нервных волокон по Дж.Эрлангеру и Х.Гассеру
| Группы волокон (по Эрлангеру и Гассеру) | Диаметр, мкм | Пороги электрического раздраженния (относительно Aa) | Длительность пика ПД 1 | Отрицательный следовой потенциал (ОСП) | Положительный следовой потенциал | Скорость проведения, м/с | |
| Длительность, мс | Амплитуда СП, % к амплитуде ПД | Длительность, мс | Амплитуда СП, % к амплитуде ПД | ||||
| Aa | 13 - 22 | 1,0 | 0,4 | 15 – 20 | 40 - 60 | 0,2 | 70 - 120 |
| Ab | 8 -13 | 40 - 70 | |||||
| Ag | 4 - 8 | 15 – 40 | |||||
| Ad | 1 – 4 | 5 – 15 | |||||
| B | 1 - 3 | 11,7 | 1,2 | ОСП нет | 100 – 300 | 3 – 14 | |
| C | 0,5 – 1,0 | 100,0 | 2,0 | 50 – 60 | 300 - 1000 | 0,5 - 2 |
1 Приблизительно ту же величину имеют и абсолютные рефрактерные фазы
Дата добавления: 2016-09-06 ; просмотров: 3712 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
ПД – волна возбуждения, распространяющаяся по мембранам нервных и мышечных клеток. ПД обладает способностью к самораспространению. ПД – обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам. Синоним ПД – нервный импульс, спайк.
По мембранам мышечных клеток и в безмякотных нервных волокнах возбуждение распространяется непрерывно вдоль всей мембраны.
В волокнах, покрытых миелиновой оболочкой, образованной шванновскими клетками, ПД может распространяться только скачкообразно (сальтаторно), перепрыгивая через участки волокна, покрытые миелином, с одного перехвата Ранвье на другой.
Причины сальтаторного проведения:
1) В перехватах Ранвье, свободных от миелина, сопротивление электрическому току
2) Порог раздражения в перехватах Ранвье минимальный.
3) Велика плотность натриевых каналов на мембране перехвата (12000 на 1 мкм2 – это
значительно больше, чем в любом другом участке волокна).
4) Возбуждение, возникающее в одном перехвате Ранвье, вызывает смещение ионов во внешней и внутренней средах данного волокна и этого смещения достаточно, чтобы вызвать возбуждение в соседнем участке.
Особенности декрементного и бездекрементного распространения волны возбуждения.Декремент – постепенное ослабление.
1) Наблюдается в безмиелиновых волокнах.
2) Свойственно волокнам, которые иннервируют внутренние органы, обладающие низкой
3) Скорость распространения возбуждения невелика и определяется диаметром волокна.
1) ПД проходит весь путь от места раздражения до места реализации без затухания.
2) Характерно для миелиновых волокон, которые передают сигналы к органам,
обладающим высокой реактивностью.
3) Время проведения возбуждения обратно пропорционально длине между перехватами
Скорость проведения возбуждения по нервному волокну зависит от диаметра волокон и наличия перехватов Ранвье. Диаметр волокна определяет характеристики электротонического (зависимого от полюсов) распространения мембранных токов. При увеличении диаметра волокна продольное сопротивление его внутренней среды, определяемое площадью поперечного сечения, снижается относительно сопротивления мембраны. В результате электротонические токи распространяются на большее расстояние и возрастает скорость проведения. Длина участков между перехватами Ранвье различна и также зависит от толщины нервного волокна: чем оно толще, тем длиннее расстояние между перехватами.
| Классификация нервных волокон по Эрлангеру и Гассеру (1937)Группа волокон | Диаметр волокна, мкм | Скорость проведения, м/с |
| A A A A | 12-20 5 – 12 4 – 8 2– 5 | 70 – 120 30 – 70 15 – 40 5 – 15 |
| B | 1 – 3 | 3 – 4 |
| C | 0,3-1,3 | 0,5 – 2,3 |
Аа- миелиновые афферентные волокна от механорецепторов кожи,мышечных и сухожильных рецепторов.Эфферентные волокна к скелетным мышцам.
Ав- миелиновые афферентные волокна от кожных рецепторов прикосновения и давления и от части мышечных рецепторов.
Аd-миелиновые афферетные волокна от части тактильных,температурных и болевых рецепторов кожи, а также суставных рецепторов
Волокна С-немиелиновые постганглионарные волокна вегетативной нервной системы, волокна обонятельных нервов, афферентные волокна от некоторых болевых,тепловых, висцеральных и обонятельных рецепторов.
Физиологические свойства нервных и глиальных клеток, их взаимосвязанная деятельность. Трофическая функция нервных и глиальных клеток.
Нейроглия.
Структурной и функциональной единицей ЦНС является нервная клетка (нейрон), которая окружена клетками нейроглии.
Нейроны.
Нейроны – специализированные клетки, способные принимать, обрабатывать, кодировать, передавать и хранить информацию; способные устанавливать контакты с другими нейронами, клетками органов; способные генерировать электрические разряды и передавать информацию с помощью специализированных окончаний – синапсов.
Серое вещество мозга – тела нейронов. Белое вещество различных отделов мозга – отростки нейронов.
Тела нейронов выполняют трофическую функцию по отношению к их отросткам (гибель тела клетки ведет к дегенерации ее отростков).
Классификация по характеру отростков-
псевдоуниполярные -нейроны спинальых ганглиев
биполярные- обонятельные рецепторы,биполярные нейроны сетчатки, нейроны спирального ганглия
+ инфа из вопроса 3.
Нейроглия(глиоциты) – совокупность всех клеточных элементов нервной ткани кроме нейронов. В мозге взрослого человека 1150 – 200 млрд. глиальных клеток, что в 10 раз больше нервных. Нейроглия делится на микроглию (глиальные макрофаги) и макроглию (астроциты, олигодендроциты, эпендимоциты).
Астроцитысоставляют 45 – 60% серого вещества мозга. Покрывают 85% поверхности капилляров мозга (сосудистые ножки астроцитов), крупные отростки астроцитов контактируют с телами нейронов. Основная функция – трофическая.
Олигодендроцитыобразуют миелин в нервной системе и поддерживают его целостность.
Эпендимоциты –клетки, выстилающие стенки спинномозгового канала и всех желудочков головного мозга. Это граница между спинномозговой жидкостью (ликвор) и тканью мозга.
Нервные волокна (neurofibrae) – это отростки нервных клеток, окруженные оболочками из нейроглиальных клеток.
Существуют различные классификации нервных волокон.
По наличию (или отсутствию) миелиновой оболочки нервные волокна делят на миелиновые и безмиелиновые.
Классификация нервных волокон по Эрлангеру – Гассеру основана на исследовании составного потенциала действия смешанного периферического нерва (например, седалищного). После электрической стимуляции такого нерва регистрируется целый спектр потенциалов действия, отражающий наличие в нерве волокон с разными скоростями проведения импульса. Согласно этой классификации выделяют три основных типа волокон – А, В, С, соответствующие трем волнам составного потенциала действия. Волокна типа А подразделяются на 4 подтипа – α, β, γ, δ. Данная классификация учитывает как чувствительные, так и двигательные волокна.
Классификация нервных волокон по Ллойду – Ханту учитывает диаметр волокна и скорость проведения импульса, но применяется только для чувствительных волокон. Согласно этой классификации выделяют четыре класса волокон – I,II,III,IV. ВолокнаIкласса подразделяют на 2 группы – а иb. Эти классификации не противоречат друг другу и представлены в сводной таблице.
Функция, тип обслуживаемого рецептора
Чувствительные нервные волокна
Первичные афферентные волокна мышечных веретен и от сухожильных органов Гольджи.
Вторичные афферентнфе волокна мышечных веретен, кожные механорецепторы
Кожные механорецепторы, терморецепторы, ноцицепторы
Кожные механорецепторы, терморецепторы, ноцицепторы (немиелинизированные волокна)
Двигательные нервные волокна
Экстрафузальные волокна скелетных мышц
Интрафузальные мышечные волокна
Преганглионарные вегетативные волокна
Постганглионарные вегетативгые (немиелинизированные волокна)
Существует прямая зависимость между диаметром аксона, толщиной миелиновой оболочки, расстоянием между узловыми перехватами Ранвье и скоростью проведениянервного импульса.
Функциональная анатомия спинного мозга Внешнее строение
Спинной мозг (medulla spinalis) располагается в позвоночном канале от уровня большого затылочного отверстия до 1(♂) –2 (♀) поясничных позвонков. Состоит из 31 сегмента (8 шейных, 12 грудных, 5 поясничных, 5 крестцовых, 1 копчиковый).
Сегмент – это участок спинного мозга, соответствующий одной паре спинномозговых нервов. Каждый спинномозговой нерв образуется в результате слияния заднего (чувствительного) и переднего (двигательного) корешков. На заднем корешке имеется чувствительный спинальный ганглий, в котором расположены тела чувствительных (псевдоуниполярных) нейронов, их центральные отростки в составе задних корешков входят в спинной мозг, периферические идут в составе спинномозговых нервов к рецепторам, расположенным в коже, мышцах, внутренних органах. Передние корешки образованы аксонами двигательных нейронов, расположенных в передних рогах спинного мозга. Передний и задний корешки соединяются в области межпозвоночного отверстия. Спинной мозг имеет шейное (от С4до Т1) и пояснично-крестцовое (L1-S2) утолщения, которые иннервируют верхние и нижние конечности соответственно.
Спинной мозг двусторонне симметричен. Вдоль его передней поверхности проходит передняя срединная щель (fissura mediana anterior), вдоль задней поверхности – задняя срединная борозда (sulcus medianus posterior). Латеральнее передней срединной щели проходит передне-латеральная борозда (sulcus anterolateralis), из которой выходят передние корешки спинного мозга. Латеральнее задней срединной борозды проходит задне-латеральная (sulcus posterolateralis) борозда, в области которой в спинной мозг входят задние корешки спинного мозга. Внутри спинного мозга находится узкий центральный канал (canalis centralis) (полость спинного мозга), заполненный спинно-мозговой жидкостью.
Спинной мозг покрыт тремя оболочками: твердой мозговой оболочкой, паутинной оболочкой и сосудистой оболочкой. Между паутинной и мягкой мозговыми оболочками находится подпаутинное пространство, заполненное спинно-мозговой жидкостью.
Гуманитарный факультет
Кафедра психологии
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Бишкек - 2005
Составила: к.м.н., доцент кафедры психологии КРСУ Обидина О.К.
Введение
Основными функциями центральной нервной системы являются объединение деятельности различных систем организма и осуществление взаимодействия целостного организма с внешней средой. Центральная нервная система осуществляет свои функции по механизму рефлекторного акта (рефлекса) при тесном и согласованном взаимодействии всех отделов мозга. И.П. Павлов экспериментально доказал, что рефлекторный акт является физиологическим механизмом и для психической деятельности, осуществляемой корой больших полушарий головного мозга. Однако для изучения сложной нервной деятельности приходится рассматривать работу каждого отдела, искусственно выделяя его из всей системы. При таком аналитическом изучении функций различных отделов центральной нервной системы не следует забывать об их взаимодействии в естественных условиях жизнедеятельности.
Тема 1. Общая физиология возбудимых структур.
Вопросы для самоподготовки.
I. Биомембраны. Транспорт веществ через мембраны .
1. Биологические мембраны, определение, функции, строение.
2. Свойства клеточных мембран (асимметричность, селективная проницаемость, изменчивость, электрогенез).
3. Транспорт веществ через мембраны. Виды транспорта веществ (активный, пассивный), их характеристика.
II. Биоэлектрические явления в живых тканях.
1. Понятия о раздражимости, возбудимости и возбуждении. Сравнительная характеристика возбудимости и раздражимости. Меры измерения возбудимости.
2. Понятие о биоэлектрических потенциалах. История их открытия. Методы регистрации биопотенциалов.
3. Основные положения мембранной теории биотоков (селективная проницаемость клеточной мембраны, ионная асимметрия).
4. Виды потенциалов:
- потенциал покоя (определение, механизм образования);
- потенциал действия (определение, компоненты ПД, механизм образования).
5. Изменение возбудимости при возбуждении. Соотношение фаз изменения возбудимости с потенциалом действия (ПД).
6. Местное возбуждение и волна возбуждения (условия их возникновения, особенности).
III. Физиология нейрона. Физиология синапса.
1. Нейрон (особенности строения, функции его составных частей).
2. Нервные волокна (строение, функции, классификация).
3. Механизм проведения возбуждения по нервным волокнам.
4. Законы проведения возбуждения по нерву (их характеристика).
5. Синапс (понятие, строение, классификация).
6. Механизм передачи возбуждения через синапс.
7. Постсинаптический потенциал (особенности, виды).
8. Основные свойства синапсов.
Основополагающий материал.
Живые клетки ограничены от окружающей их среды плазматической мембраной. Основной матрикс мембраны состоит из липидов, главным образом фосфолипидов. Они представлены головной гидрофильной группой, к которой присоединены длинные гидрофобные углеродные цепи. Клеточные мембраны представляют собой липидные бислои, где гидрофильные группы обращены к водной среде, а гидрофобные – друг к другу, образуя безводную липидную фазу.
Главными функциональными элементами, погруженными в сравнительно инертный липидный матрикс мембраны, являются белки. Белок по массе составляет от 25 до 75% в различных мембранах. Одни белки пронизывают мембрану от ее наружной до внутренней поверхности, другие же закреплены в каком-то одном слое. При этом гидрофобные группы белковых молекул погружены в липидную мембрану, а гидрофильные группы на поверхности мембраны погружены в водную среду. Углеводы занимают незначительную часть структуры мембраны. Порядок расположения белков, жиров и углеводов в мембранах различных тканей разнообразен. В мембране имеются ионные каналы. Они образованы молекулами белков и обладают специфичностью (проницаемы только для определенных веществ, ионов).
Биомембраны выполняют следующие функции:
1) барьерную, которая включает в себя ограничение клетки от окружающей среды, ограничение органоидов клетки и избирательную проницаемость веществ для поддержания гомеостаза;
2) транспортную – организация обмена веществ между клеткой и окружающей средой;
3) участие в межклеточном взаимодействии, состоящее в способности воспринимать сигналы и участвовать в иммунных реакциях;
4) рецепторную;
5) генерацию биопотенциалов.
Толщина мембраны колеблется от 4 до 10 мк.
Основные свойства клеточных мембран:
1) асимметричность (структурная, ионная и электрическая);
2) избирательная проницаемость для веществ;
Клетку нельзя считать статичным образованием, так как здесь происходит постоянный обмен веществами между внутриклеточными элементами и окружающей средой. При этом структура мембраны находится в динамическом состоянии.
Транспорт веществ через мембрану осуществляется двумя путями: активным и пассивным.
Пассивный транспорт веществ идет по градиенту (разница параметра между двумя точками пространства) и без затрат энергии АТФ.
Пассивный транспорт характеризуется следующими механизмами:
1) диффузия – передвижение веществ по концентрационному градиенту (растворимые вещества – ионы, О2, СО2, жирорастворимые вещества);
2) осмос – диффузия воды как растворителя по осмотическому градиенту;
3) фильтрация – механическое разделение раствора через мембранные поры (по гидростатическому градиенту).
Активный транспорт - перенос веществ через клеточную мембрану с затратой биологической энергии молекул АТФ, протекающий против электрохимического градиента.
Активный транспорт реализуется с помощью нескольких типов молекулярных машин – насосов (Nа + , К + , Са ++ ), переносчиков, а также эндо- и экзоцитоз.
Общим свойством всех живых организмов является раздражимость. Раздражимостью называется способность организма отвечать на изменения внешней и внутренней среды изменением интенсивности обмена веществ. Воздействующие на организм факторы называются раздражителями, которые можно разделить на три большие группы: физические (световые, звуковые, механические, термические, электрические и т.д.), химические (щелочи, кислоты, соли, яды и т.д.) и раздражители биологического происхождения. Раздражители могут различаться также по силе, скорости нарастания в единицу времени и длительности воздействия на организм.
Однако ответная реакция некоторых тканей на раздражение характеризуется важными качественными особенностями. Наиболее анатомически и функционально дифференцированные ткани (нервная, мышечная, железистая) реагируют на раздражение строго определенным, специализированным процессом – возбуждением. Способность высокодифференцированных тканей отвечать на раздражение специфической ответной реакцией (сокращением, образованием и выделением секрета, генерацией и проведением нервного импульса) называется возбудимостью.
Возбудимость величина непостоянная и зависит от вида ткани (у нервной ткани выше, чем у мышечной и железистой) и ее состояния (при утомлении ткани снижается). Возбудимость тканей измеряется порогом раздражения и полезным временем.
Порог раздражения – это та минимальная сила раздражителя, которая способна вызвать минимальную регистрируемую ответную реакцию. Раздражители, сила которых меньше пороговой, называются допороговыми; больше пороговой – сверхпороговыми.
Полезное время – это минимальное время, в течение которого раздражитель должен действовать на возбудимую структуру, чтобы вызвать ответную реакцию.
Мышечная и железистая ткани могут находиться в состояниях покоя, возбуждения, а нервная еще и в состоянии торможения. Электрически такие состояния проявляются в виде мембранного потенциала (потенциала покоя, ПП) и потенциала действия (ПД).
Предположения о наличии электричества в живом организме были высказаны еще в конце 18 века Л. Гальвани. Далее его опыты были продолжены такими учеными, как А. Вольта, К. Маттеучи, Г. Гельмгольц и многие другие. В 1896 году русский ученый В.Ю.Чаговец предложил гипотезу об ионном механизме биопотенциалов. Он предположил и обосновал экспериментально, что наличие электрических потенциалов в клетках обусловлено неравенством концентрации ионов Na + , K + , Ca ++ , Cl - внутри и вне клетки, и различной проницаемостью для них поверхностной мембраны.
В 1902 году Ю.Бернштейн развил мембранно-ионную теорию, которую модифицировали А.Ходжкин, А.Хаксли и Катц (1949-1952). Эта теория и в настоящее время пользуется всеобщим признанием.
Мембранный потенциал (потенциал покоя – ПП) – это разность потенциалов между наружной и внутренней поверхностями мембраны клетки в состоянии физиологического покоя. У разных видов тканей этот потенциал варьирует от -60 до -100мВ, за исключением гладко–мышечных клеток (-30мВ).
В основе механизма образования ПП лежат следующие свойства мембраны:
ионная асимметрия и избирательная проницаемость. Известно, что концентрация некоторых ионов (Na + , K + , Cl - ) неодинакова внутри и вне клетки. Так, например, ионов K + внутри клетки в 50-100 раз больше, чем во внеклеточной жидкости. Ионов Na + , наоборот, вне клетки в 8-15 раз больше, чем внутри. Следует также учитывать и разность концентраций анионов (Cl - ), кислотных остатков и радикалов белка. Трансмембранный градиент концентраций основных ионов (Na + и K + ) создается и поддерживается калий–натриевым насосом.
Избирательная (селективная) проницаемость мембраны. Известно, что мембрана клетки, находясь, как в состоянии покоя, так и в состоянии возбуждения, пропускает не все, а только определенные ионы. В покое клетка более проницаема для ионов К + , чем для других ионов. Ионы К + пассивно, по концентрационному градиенту выходят во внеклеточную жидкость, и создают вместе с другими катионами (Na + ) избыток положительных зарядов на внешней поверхности мембраны. Поскольку отрицательные ионы не могут проходить через каналы клеточной мембраны, они остаются внутри клетки и локализуются, в основном, на внутренней поверхности мембраны, тем самым увеличивая внутренний отрицательный заряд. Транспорт других ионов (Na + , Cl - ) через мембрану покоящейся клетки очень незначителен и, поэтому, не учитывается в механизме формирования ПП.
Величина ПП в различных тканях обусловлена степенью избирательной проницаемости.
Рис. 1. Изменение мембранного потенциала клетки (А) при действии электрического тока различной силы (Б).
ЭП - электротонический потенциал, ЛО – локальный ответ, ПД – потенциал действия.
Потенциал действия, возникнув в возбужденной области мембраны, приводит к нарушению ионного равновесия в соседних областях. Происходит распространение возбуждения. Но возбуждение приобретает распространяющийся характер при определенных условиях. Обязательным условием является достаточная возбудимость ткани. Кроме того, сила раздражающего стимула должна быть равна или превышать порог возбуждения. Если сила раздражающего стимула меньше пороговой величины, возникает местное возбуждение (локальный ответ) в области действия раздражителя. Местное возбуждение характеризуется развитием небольшой начальной деполяризации (уменьшением исходного мембранного потенциала). Местное возбуждение самопроизвольно затухает, если действие слабого раздражителя прекратилось. Амплитуда локального ответа прямо пропорциональна величине стимула. Если величина раздражающего стимула превышает порог, местное возбуждение достигает критического уровня и переходит в распространяющееся возбуждение.
Еще одним важным свойством возбудимости является то, что в период генерации ПД происходит развитие рефрактерности мембраны. В период деполяризации возбудимость ее падает до нуля вследствие инактивации натриевых каналов. Возбуждение на мембране не возникает даже при действии на нее раздражителя сверхпороговой силы. Такое состояние полной невозбудимости называется абсолютным рефрактерным периодом. За ним следует относительный рефрактерный период, когда действием раздражителя сверхпороговой силы все же можно вызвать ПД, хотя его амплитуда и будет снижена по сравнению с нормой. Этот период соответствует фазе реполяризации. Возвращение к нормальной возбудимости проходит еще период супернормальной возбудимости, соответствующий отрицательному следовому потенциалу, и период субнормальной возбудимости, соответствующий положительному следовому потенциалу (рис.2).
Рис. 2. Изменение мембранного потенциала (А), интенсивности калиевого и натриевого трансмембранного тока (Б) и возбудимости клетки (В) в разные фазы потенциала действия.
Д – фаза деполяризации,
РБ – фаза быстрой реполяризации,
РМ – фаза медленной реполяризации,
Г – фаза гиперполризации;
Н – период нормальной возбудимости,
РА – период абсолютной рефрактерности,
РО – период относительной рефрактерности,
Н+ – период супернормальной возбудимости,
Н– – период субнормальной возбудимости.
Большое значение в фазных колебаниях возбудимости имеет период абсолютной рефрактерности, который ограничивает максимальную частоту генерирования потенциалов действия. Т.е., если абсолютный рефрактерный период завершается через 2 мс после начала ПД, то клетка может возбуждаться с частотой максимум 500/с. Существуют клетки и с более коротким рефрактерным периодом - 1000/с.
Нервные волокна, составляющие нервные стволы, разделяются на две группы: мякотные (миелиновые) и безмякотные. Мякотные нервные волокна состоят из осевого цилиндра, который покрыт миелиновой оболочкой разной толщины (рис.3). Миелиновая оболочка является электрическим изолятором и выполняет трофическую функцию. Через определенные промежутки миелиновая оболочка прерывается, обнажая осевой цилиндр. Эти открытые участки (до 1 мк) называются перехватами Ранвье и играют большую роль в скорости проведения возбуждения.

Рис. 3. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А–Г); соотношение леммоцита и безмиелиновых волокон (Д)
(по Судаков, 2000).
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно.
Безмякотные нервные волокна покрыты только шванновской оболочкой. Внутри осевого цилиндра – в аксоплазме – проходит множество нейрофибрилл диаметром от 100 до 400 Å (ангстрем). По ним транспортируются различные вещества (медиаторы, нейропептиды, гормоны, питательные вещества и др.) от тела нейрона к нервному окончанию и наоборот.
Плазматическая мембрана осевого цилиндра нервного волокна имеет очень сложное строение и приспособлена к генерации и проведению нервных импульсов.
Возбужденный участок нервного волокна вызывает ионный сдвиг с последующей генерацией ПД, который, в свою очередь, является сверхпороговым стимулом для соседних участков. Вследствие чего, в них также возникает ионный сдвиг и возбуждение. При этом, волна возбуждения в безмякотных волокнах распространяется вдоль мембраны непрерывно (рис.4.1.) В миелиновых волокнах возбуждение (ПД) распространяется от одного перехвата Ранвье к другому или даже через несколько перехватов. Таким образом, по мякотным волокнам передача возбуждения передается скачкообразно (сальтоторно) и, тем самым определяет высокую скорость проведения возбуждения (рис.4.2.).
Рис. 4.1. Механизм распространения возбуждения по безмиелиновому нервному волокну.
Рис. 4.2. Механизм распространения возбуждения по миелиновому нервному волокну.
Гассер и Эрлангер показали, что не только наличие миелиновой оболочки, но и толщина нервного волокна влияют на скорость проведения возбуждения.
Классификация нервных волокон по Эрлангеру – Гассеру
Читайте также:
