
К миелинизированным нервным волокнам относятся
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
распространяется по нервным волокнам с затуханием (с декрементом ), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим .
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
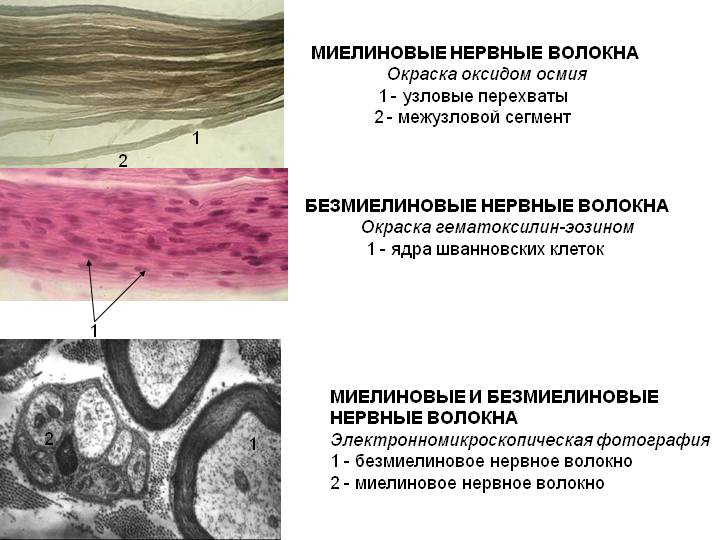
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты ( шванновские клетки ) сначала прикасаются к аксону, а затем окутывают его (рис. 1, А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис. 1, В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Рис. 1. Формирование миелиновой оболочки вокруг аксона на разных стадиях его развития (А – Г); соотношение леммоцита и безмиелиновых волокон (Д) (по Судакову, 2000)
1 – леммоцит, 2 – миелиновое волокно, 3 – миелиновая оболочка, 4 – безмиелиновое волокно
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки (рис. 1, Д).
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление.
Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 2, А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 2, Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 2, В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна.
Рис. 2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте
При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
Рис. 3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в табл. 4.1.
Таблица 4.1. Типы нервных волокон, их свойства и функциональное назначение
Тип
Диаметр (мкм)
Миелинизация
Скорость про-ведения (м/с)
Функциональное назначение
Двигательные волокна соматической НС; чувствительные волокна проприорецепторов
Чувствительные волокна кожных рецепторов
Чувствительные волокна проприорецепторов
Чувствительные волокна терморецепторов, ноцицепторов
Преганглионарные волокна симпатической НС
Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.
- Виртуальная лаборатория
- Мочевыводящая система
- Сердечно-сосудистая система
- Влияние давления и вязкости жидкости, а также радиуса и длины сосуда на движение жидкости по сосуду
- Воздействие адреналина, ацетилхолина, атропина и адреналина на основе атропина на артериальное давление
- Воздействие медикаментов и химических медиаторов на деятельность сердца
- Воздействие электрических стимулов на сердечную деятельность
- Воздействие возбуждения блуждающего нерва на сердечную деятельность
- Наложение лигатур Станниуса
- Дыхательная система
- Влияние давления в плевральной полости на вентиляцию легких
- Влияние сурфактанта на вентиляцию легких
- Механизм дыхания. Объемы и емкости легких. Влияние радиуса просвета дыхательных путей
- Внутренняя среда организма. Кровь
- Определение групп крови системы ABO с использованием стандартных сывороток
- Обмен веществ и энергии. Питание
- Физиология человека и животных
- ВНУТРЕННЯЯ СРЕДА ОРГАНИЗМА. КРОВЬ. ИММУНИТЕТ
- План. Внутренняя среда организма. Кровь. Иммунитет
- 1. Функции крови. Состав и физико-химические свойства крови. Плазма крови
- 2. Механизмы поддержания кислотно-основного равновесия. Буферные системы крови
- 3. Лимфа. Образование лимфы. Ликвор
- 4. Эритроциты: строение и функции. Гемолиз. Гемоглобин. Эритропоэз. Анемии
- 5. Группы крови. Агглютиногены (антигены) и агглютинины (антитела). Резус-фактор. Правила переливания крови
- 6. Строение и функции лейкоцитов. Лейкоцитарная формула
- 7. Регуляторная функция лейкоцитов (цитокины)
- 8. Защитная функция крови. Понятие о клеточном и гуморальном иммунитете
- 9. Тромбоциты, их строение и функции
- 10. Свертывающая и противосвертывающая системы крови. Плазменные и клеточные факторы свертывания
- ФИЗИОЛОГИЯ СЕРДЕЧНО–СОСУДИСТОЙ СИСТЕМЫ
- План. Физиология сердечно-сосудистой системы
- 1. Строение и функции сердечно-сосудистой системы
- 2. Строение сердца. Общие свойства сердечной мышцы: автоматия, проводимость, возбудимость и сократимость. Типичные (рабочие) и атипичные кардиомиоциты
- 3. Механизм автоматии миокарда. Градиент автоматии
- 4. Особенности кровоснабжения и энергетического обеспечения сердца. Функциональная роль предсердий и желудочков, клапанного аппарата
- 5. Сердечный цикл. Понятие о систолическом и минутном объемах крови
- 6. Методы изучения сердечной деятельности
- 7. Миогенная, нейрогенная и гуморальная регуляция деятельности сердца
- 8. Гемодинамика. Функциональные особенности различных отделов сосудистого русла. Линейная и объемная скорость движения крови. Факторы, обеспечивающие непрерывность кровотока
- 9. Капиллярное кровообращение и его особенности. Микроциркуляция. Представление о тонусе сосудов. Регуляция тонусов сосудов. Сосудодвигательный центр
- 10. Строение лимфатической системы и ее функции. Транспорт лимфы
- ФИЗИОЛОГИЯ ДЫХАНИЯ
- План. Физиология дыхания
- Дыхание у высших позвоночных: внешнее дыхание, газообмен в легких и тканях, транспорт газов кровью, тканевое дыхание
- Вентиляция легких. Механика и динамика дыхательных движений. Внутриплевральное давление и его значение. Роль сурфактанта. Аэрогематический барьер
- Показатели внешнего дыхания. Понятие о легочных объемах и емкостях. Состав вдыхаемого, выдыхаемого и альвеолярного воздуха
- Газообмен в легких и тканях. Парциальное давление кислорода и диоксида углерода в альвеолярном воздухе, венозной и артериальной крови. Транспорт кислорода кровью
- Механизм переноса диоксида углерода, роль карбоангидразы. Взаимосвязь между дыханием и поддержанием кислотно-щелочного равновесия крови
- Нервные механизмы регуляции дыхания. Защитные и регуляторные дыхательные рефлексы. Хеморецепторы. Дыхание при физической нагрузке, при повышенном и пониженном атмосферном давлении
- ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ. ПИТАНИЕ. ТЕРМОРЕГУЛЯЦИЯ
- План. Обмен веществ и энергии. Питание. Терморегуляция
- Обмен веществ и энергии как основное условие гомеостазиса. Анаболические и катаболические процессы. Обмен веществ как источник образования тепла
- Основной и общий обмен. Рабочая прибавка. Общие энергозатраты людей различных профессиональных групп
- Характеристика продуктов питания. Пищевые и питательные вещества. Нормы питания
- Значение воды для организма. Витамины и их роль в обмене веществ
- Принципы составления пищевых рационов
- Терморегуляция. Механизмы теплопродукции и теплоотдачи. Суточные изменения температуры тела у человека
- Лихорадка
- ФИЗИОЛОГИЯ МОЧЕВЫДЕЛЕНИЯ
- План. Физиология мочевыделения
- Строение и функции почек. Нефрон как функциональная единица почки. Особенности кровоснабжения почек
- Механизм образования мочи
- Реабсорбция в нефроне и ее механизмы. Поворотно-противоточная система. Механизмы осмотического концентрирования и разведения мочи
- Процессы секреции и экскреции в почечных канальцах. Образование конечной мочи, ее состав и свойства
- Гуморальная и гормональная регуляции почечной функции
- ФИЗИОЛОГИЯ СИСТЕМЫ ПИЩЕВАРЕНИЯ
- План. Физиология системы пищеварения
- Строение и функции пищеварительной системы. Типы пищеварения
- Пищеварение в ротовой полости. Функции слюнных желез, состав и свойства слюны. Регуляция слюноотделения
- Пищеварение в желудке. Механизмы желудочной секреции ферментов и соляной кислоты. Фазы желудочной секреции. Состав и свойства желудочного сока
- Пищеварение в тонком кишечнике. Пристеночное (мембранное) пищеварение. Всасывание в пищеварительной системе
- Роль печени и поджелудочной железы в пищеварении
- Пищеварение в толстом кишечнике
- Регуляция деятельности органов пищеварения
- Аппетит, голод, насыщение
- ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
- План. Физиология возбудимых тканей
- 1. Раздражимость и возбудимость как способность живых систем реагировать на действие факторов внешней среды. Ионные насосы и их функции (калий-натриевый, кальциевый, хлорный насос)
- 2. Ионные каналы, классификация, строение и функции. Потенциал действия и его фазы. Вклад потенциалзависимых ионных каналов в формирование потенциала действия
- 3. Причины существования относительной и абсолютной рефрактерности. Функциональная лабильность и ее проявления
- 4. Возбудимые клетки как проводники электричества. Распространение электротонического потенциала. Константа длины. Механизм проведения возбуждения по немиелинизированным и миелинизированным нервным волокнам
- 5. Классификация и свойства нервных волокон. Законы проведения возбуждения по нервным волокнам
- 6. Основы межклеточного взаимодействия в организме. Физиология синапсов
- 7. Химические синапсы. Механизм возникновения возбуждающего постсинаптического потенциала. Тормозной постсинаптический потенциал
- 8. Общая характеристика и классификация нейромедиаторов. Нейромодуляторы
- 9. Ионотропные и метаботропные рецепторы. Основные группы синаптически активных веществ (лиганды, агонисты, антагонисты)
- 10. Физиология мышц. Классификация мышц по структурным, биохимическим и функциональным критериям
- 11. Виды и режимы мышечных сокращений. Работа и сила мышц. Типы нервных волокон
- 12. Механизм мышечного сокращения. Роль АТФ, кальция и ионных насосов в мышечном сокращении. Энергетическое обеспечение мышечного сокращения
- ОБЩАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
- План. Общая физиология центральной нервной системы
- 1. Структурно-функциональная организация нейронов и глиальных клеток. Представления о функциональной роли глии
- 2. Строение и свойства нейронов
- 3. Понятие о нервном центре, нервной сети. Свойства нервных центров
- 4. Принципы координации работы нервных центров
- 5. Торможение в ЦНС и его виды. Интегративная деятельность нейрона
- 6. Рефлекторная теория. Компоненты рефлекторной дуги. Классификация рефлексов
- 7. Классификация врожденных форм поведения. Принцип обратной связи
- 8. Теория функциональных систем П.К. Анохина
- ЧАСТНАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
- План. Частная физиология центральной нервной системы
- 1. Функциональная организация спинного мозга и его роль в координации рефлекторной деятельности центральной нервной системы
- 2. Функции спинного мозга. Особенности управления движениями на уровне спинного мозга. Спинальный контроль мышечных сокращений и управление мышечным тонусом
- 3. Спинальные соматические и вегетативные рефлексы. Сегментарный и межсегментарный принцип работы спинного мозга
- 4. Структурно-функциональная характеристика основных отделов головного мозга
- 5. Функции продолговатого мозга, топография ядер, регуляция витальных функций
- 6. Роль ретикулярной формации мозгового ствола в регуляции функций. Варолиев мост
- 7. Средний мозг
- 8. Мозжечок: строение, связи и регулирующие влияния
- 9. Промежуточный мозг. Роль ядер таламуса в передаче сигналов с периферии в кору больших полушарий. Гипоталамус как интегративно координирующий вегетативный центр мозга
- 10. Подкорковые ганглии, их строение и функции. Экстрапирамидная двигательная система
- 11. Гиппокамп и лимбическая система, их взаимодействие при интеграции сигналов в мозге
- 12. Физиология вегетативной нервной системы. Современные представления о нейрогуморальной регуляции висцеральных функций организма. Вегетативная рефлекторная дуга
- 13. Функциональные особенности и взаимодействие отделов вегетативной нервной системы в управлении функциями организма
- 14. Основы физиологии коры больших полушарий. Представление о кортикализации функций. Цитоархитектоника коры больших полушарий
- 15. Понятие о полях и зонах коры больших полушарий
- 16. Электрическая активность коры головного мозга. Электроэнцефалограмма
- ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ФУНКЦИЙ
- План. Гормональная регуляция функций
- 1. Эндокринный контроль функций и его регуляторная роль. Участие желез внутренней секреции в интегративной регуляции деятельности организма
- 2. Роль эндокринной системы в регуляции процессов роста, развития, размножения, разных форм адаптации, поведения. Классификация гормонов
- 3. Современные взгляды на механизмы влияния гормонов: рецепторы и вторые посредники
- 4. Гипоталамо-нейрогипофизарная и гипоталамо-аденогипофизарная системы. Гипоталамические рилизинг – факторы (либерины и статины)
- 5. Структура и функции долей гипофиза, секретируемые тропные и эффекторные гормоны, их роль в организме
- 6. Эпифиз и роль мелатонина у животных и человека
- 7. Щитовидная железа. Тиреоидные гормоны (трииодтиронин и тироксин) и кальцитонин
- 8. Паращитовидные железы
- 9. Эндокринная функция поджелудочной железы и ее гормоны (инсулин, глюкагон, соматостатин)
- 10. Гормоны коркового и мозгового слоя надпочечников
- 11. Роль надпочечных желез в реализации адаптационно-приспособительной деятельности организма (стресс). Фазы стресса
- 12. Половые железы и их внутренняя секреция. Гормональная функция семенников. Гормональная функция яичников. Половые циклы
- 13. Эндокринная функция почек, желудочно-кишечного тракта, сердца. Гормональная функция эндотелия
- ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ
- План. Физиология сенсорных систем
- 1. Принципы строения сенсорных систем. Понятие о рецепторах
- 2. Классификация рецепторов. Механизм возбуждения рецепторов. Рецепторный и генераторный потенциалы. Адаптация рецепторов
- 3. Кодирование сенсорной информации. Проводниковые структуры сенсорных систем
- 4. Зрительный анализатор
- 4. I. Преломление света в оптических средах глаза. Построение изображения на сетчатке
- 4. II. Типы движений глаз
- 4. III. Строение сетчатки. Фоторецепторы и преобразование световой энергии в электрический сигнал
- 4. IV. Представления о механизмах цветного зрения и обработке зрительной информации в ЦНС
- 4. V. Проводниковый и корковый отделы зрительного анализатора
- 5. Слуховой анализатор
- 5. I. Строение слухового анализатора
- 5. II. Строение улитки. Орган Корти. Фонорецепторы. Звуковосприятие
- 5. III. Проводящие пути и корковое представительство слухового анализатора. Представления о механизме восприятия частоты и интенсивности звука
- 6. Вестибулярный анализатор
- 7. Вкусовой анализатор: структурные и функциональные особенности. Клеточные механизмы вкусовой рецепции
- 8. Обонятельный анализатор: структурные и функциональные особенности обонятельного анализатора. Клеточные механизмы обонятельной рецепции
- 9. Соматосенсорная система. Кожные рецепторы: тактильные, температурные, болевые. Мышечно-суставная рецепция (проприорецепция)
- 10. Болевые рецепторы. Защитные (ноцицептивные) рефлексы. Современные теории боли
- ФИЗИОЛОГИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
- План. Физиология высшей нервной деятельности
- 1. Понятие о ВНД. Современные подходы к нейробиологическим механизмам поведения
- 2. Роль эмоций и мотиваций в организации поведения
- 3. Условный рефлекс как приспособительный механизм в животном мире. Классические и инструментальные условные рефлексы. Классификация условных рефлексов
- 4. Нейрофизиологическая сущность торможения. Характеристика внешнего безусловного торможения. Запредельное торможение, его биологическое значение. Основные виды условного торможения
- 5. Нейрофизиологическая сущность сна. Электроэнцефалографическая характеристика медленноволнового сна и парадоксального сна
- 6. Память как свойство центральной нервной системы воспринимать, хранить и воспроизводить информацию
- НЕЙРОГУМОРАЛЬНАЯ РЕГУЛЯЦИЯ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
- План. Нейрогуморальная регуляция физиологических функций
- 1. Общее понятие о процессах регуляции функций. Гомеостазис. Принципы регуляции функций
- 2. Механизмы регуляции жизнедеятельности организма. Нервная регуляция как высший этап развития приспособлений организма к меняющимся условиям существования
- 3. Единство нервных, гуморальных и иммунных механизмов регуляции
- ВНУТРЕННЯЯ СРЕДА ОРГАНИЗМА. КРОВЬ. ИММУНИТЕТ
4. Возбудимые клетки как проводники электричества. Распространение электротонического потенциала. Константа длины. Механизм проведения возбуждения по немиелинизированным и миелинизированным нервным волокнам
Мембрана нейрона и его отростков (нервных волокон) обладает способностью не только генерировать потенциал действия, но и передавать го на относительно большие расстояния без уменьшения его амплитуды и расходования энергии, поэтому нервные импульсы распространяются без затухания (бездекрементно). Способность к распространению возбуждения связана с тем, что во время потенциала действия происходит изменение знака заряда в возбужденном участке мембраны (инверсия заряда). Между ним и невозбужденными соседними участками мембраны возникают локальные электрические токи, под действием которых происходит деполяризация новых соседних участков, что приводит к формированию в них потенциала действия. За счет последовательного охвата возбуждением все новых участков нервного волокна возбуждение перемещается вдоль дендритов и аксонов. Чем больше диаметр волокна, тем больше скорость распространения возбуждения.
Особенности проведения возбуждения в нервных волокнах определяются наличием или отсутствием у них оболочки. Нервные волокна делятся на миелиновые и безмиелиновые, то есть покрытые особой миелиновой оболочкой или не покрытые. Эта оболочка образуется на периферии шванновскими клетками, а в головном мозге – клетками олигодендроглии.

Механизм проведения импульса по нервным волокнам объясняется теорией “местных токов” Тасаки (рисунок 3 и рисунок 4). В безмиелиновом нервном волокне ПД генерируется каждой точкой поверхности мембраны – так называемое непрерывное проведение возбуждения. Во время пикового ПД на наружной поверхности волокна происходит перезарядка, в результате чего возникает местный ток от соседнего положительно заряженного участка к возбужденному, а внутри – наоборот. Таким образом образуется кольцевой ток.
В миелиновых волокнах слой миелина не целиком охватывает волокно, а имеет промежутки – перехваты Ранвье. Сам миелин обладает высоким сопротивлением электрическому току, поэтому в участках, покрытых миелином, потенциал действия генерироваться не может. Перехваты Ранвье располагаются примерно через каждые 0,5-1,5 мм (в зависимости от толщины оболочки), в них обнаружено очень много (около 12 тыс/мкм 2 ) натриевых каналов, поэтому в миелиновых волокнах проведение возбуждения происходит скачкообразно (сальтаторно).
Между соседними участками возникает большой по величине ток (ПД = 100-120 мВ), который в 3-5 раз превышает порог раздражения и способен возбудить соседние участки, что составляет фактор надежности.
Скорость проведения возбуждения пропорциональна корню квадратному от диаметра волокна. Самые тонкие волокна – безмиелиновые, а в миелиновых волокнах чем толще слой миелина, тем больше расстояние между перехватами Ранвье. В безмиелиновых волокнах местные кольцевые токи последовательно распространяются “маленькими шажочками” непрерывно, скорость проведения возбуждения в безмиелиновых волокнах гораздо ниже, чем в миелиновых. Такая передача импульса является медленной и неэкономичной в плане энергозатрат. Поэтому безмиелиновыми являются только самые тонкие волокна (менее 1 мкм в диаметре), проводящие возбуждение к медленно работающим органам – кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре.
Константа длины – это расстояние, на котором местный ток сохраняет пороговое значение, то есть способен вызывать образование ПД на следующем участке нервного волокна.
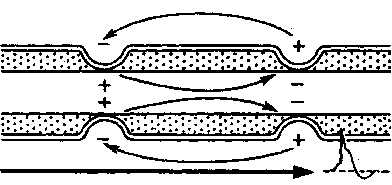
Рисунок 3 – Передача возбуждения в миелинизированном нервном волокне

Рисунок 4 – Передача возбуждения в немиелинизированном нервном волокне
Такая передача импульса является медленной и неэкономичной в плане.
Они, как и немиелинизиро- ванные, окружены глиальными клетками (они называются шванновские клетки), но мембраны этих клеток плотно прилегают к мембране нервного волокна. Сами шванновские клетки уплощаются, охватывают аксон и многократно закручиваются вокруг него наподобие изоляции электрического кабеля. Соприкасающиеся мембраны шванновской клетки смыкаются, образуя плотные пластинки — мезаксон. Смыкание и образование мезак- сона происходит за счет взаимодействия белков, примыкающих к внутренней стороне мембраны.
Белки наружной стороны мембраны тоже взаимодействуют, образуя неплотные пластинки, которые чередуются с плотными. В зависимости от диаметра аксона оболочка, образованная вокруг нервного волокна шванновской клеткой, может содержать от 10 до 200 мембранных слоев. При этом всегда сохраняется сома шванновской клетки, содержащая основные органеллы. Принципиальное строение миелинизированного нервного волокна показано на рис. 2.22. Миелиновая оболочка, таким образом, представляет собой совокупность мембран шванновских клеток. Основным компонентом мембран являются фосфолипиды (с высоким содержанием сфингомиелина), обладающие хорошими изоляционными свойствами, т.е. высоким электрическим сопротивлением.

Рис. 2.22. Схема строения миелинизированного нервного волокна
Каждая шванновская клетка, накрученная на аксон, создает по ходу аксона миелинизированный участок длиной 1—2 мм. Между последовательно расположенными шванновскими клетками всегда остается неизолированная (немиелинизированная) область волокна длиной 2—3 мкм, где ионы могут свободно переходить через мембрану из внеклеточной жидкости в аксоплазму и обратно. Эту область аксона называют перехватом Ран- вье. Таким образом, мембрана аксона состоит из регулярно чередующихся миелинизированных (межперехватных) участков длиной 1—2 мм и перехватов Ранвье длиной 2—3 мкм (см. рис. 2.22). В ЦНС миелинизированные нервные волокна выглядят так же, как в периферических нервах. Единственная особенность состоит в том, что в ЦНС одна клетка глии (оли- годендроцит) способна давать отростки к нескольким аксонам, формируя миелиповую оболочку вокруг каждого из них.

Рис. 2.23. Схема сальтаторного распространения ПД по миелинизированному нервному волокну:
ПД, возникший в одном возбужденном перехвате Ранвье, вызывает развитие локальных токов, замыкающихся лишь в следующем перехвате, где происходят деполяризация мембраны и генерация следующего ПД
Между таким возбужденным и соседними невозбужденными миелини- зированными участками мембраны возникает разность потенциалов. Эта разность порождает местные электрические токи, но они не могут выходить наружу через миелиновую оболочку ввиду ее высокого сопротивления. Поэтому нерастраченные утечкой в наружную среду локальные токи текут дальше внутри аксона по аксоплазме до соседнего невозбужденного перехвата Ранвье (см. рис. 2.23). Только там они могут проходить через мембрану, гасить ее электронегативный заряд и замыкаться.
Наряду с высоким фактором надежности сальтаторпое проведение ПД имеет еще ряд преимуществ по сравнению с непрерывным. Скачкообразная генерация ПД повышает скорость проведения возбуждения в миелинизированных волокнах в 5—50 раз. Действительно, длина межиерехватных участков составляет около 2 мм, а перехватов Ранвье — 1—2 мкм. С учетом того, что возбуждение может возникать не в следующем, а во втором или третьем перехвате, получается, что ПД распространяется по волокну скачками длиной 2—4 мм. Кроме того, сальтаторное проведение возбуждения экономит энергию для аксона. В миелинизированных волокнах деполяризуются только перехваты, что позволяет приблизительно в 100 раз снизить потерю ионов. В связи с этим снижаются траты энергии, необходимые для восстановления трансмембранной разности концентраций ионов натрия и калия после проведения серии нервных импульсов. Наконец, в крупных миелинизированных волокнах имеется еще одна особенность сальтаторного проведения: высокая изоляция миелиновой оболочкой в сочетании с 50-кратно сниженной электрической емкостью межперех- ватной мембраны позволяет осуществлять реполяризацию ПД путем перемещения очень незначительного числа ионов.
Важнейшие закономерности процесса распространения возбуждения по нервным волокнам следующие:
- 1) потенциал действия распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
- 2) генерация ПД нервными волокнами практически не вызывает их утомления;
- 3) нервные волокна обладают высокой лабильностью, т.е. могут воспроизводить потенциал действия с очень высокой частотой;
- 4) расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
- 5) распространение потенциала действия — активный процесс, в ходе которого изменяется состояние ионных каналов мембраны волокна, а также расходуется энергия гидролиза АТФ для восстановления трансмембранных ионных градиентов;
- 6) ПД распространяется по каждому нервному волокну изолированно — не переходит с одного волокна на другое. Это обусловлено значительно более низким сопротивлением межклеточной жидкости по сравнению с сопротивлением мембраны волокон. Из-за этого текущие между возбужденным и невозбужденным участками наружные местные токи проходят в основном по межклеточной жидкости, не затекая и не действуя на другие волокна;
- 7) проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность. Фактор надежности проведения сигнала у миелинизированных нервных волокон выше, чем у немиелинизированных.
Читайте также:
