
Как называется узел из нервных клеток у червей и моллюсков
3. РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ В ФИЛОГЕНЕЗЕ
Для беспозвоночных животных характерно наличие нескольких источников происхождения нервных клеток. У одного и того же типа животных нервные клетки могут одновременно и независимо происходить из трех разных зародышевых листков. Полигенез нервных клеток беспозвоночных является основой разнообразия медиаторных механизмов их нервной системы.
Нервная система впервые появляется у кишечнополостных животных. Кишечнополостные — это двухслойные животные. Их тело представляет собой полый мешок, внутренняя полость которого является пищеварительной полостью. Нервная система кишечнополостных принадлежит к диффузному типу. Каждая нервная клетка в ней длинными отростками соединена с несколькими соседними, образуя нервную сеть. Нервные клетки кишечнополостных не имеют специализированных поляризованных отростков. Их отростки проводят возбуждение в любую сторону и не образуют длинных проводящих путей. Контакты между нервными клетками диффузной нервной системы бывают нескольких типов. Существуют плазматические контакты, обеспечивающие непрерывность сети (анастомозы). Появляются и щелевидные контакты между отростками нервных клеток, подобные синапсам. Причем среди них существуют контакты, в которых синаптические пузырьки располагаются по обе стороны контакта — так называемые симметричные синапсы, а есть и несимметричные синапсы: в них везикулы располагаются только с одной стороны щели.
Нервные клетки типичного кишечнополостного животного гидры равномерно распределены по поверхности тела, образуя некоторые скопления в районе ротового отверстия и подошвы (рис. 8). Диффузная нервная сеть проводит возбуждение во всех направлениях. При этом волну распространяющегося возбуждения сопровождает волна мышечного сокращения.
Рис. 8. Схема строения диффузной нервной системы кишечнополостного животного:
1 — ротовое отверстие; 2 — щупальце; 3 — подошва
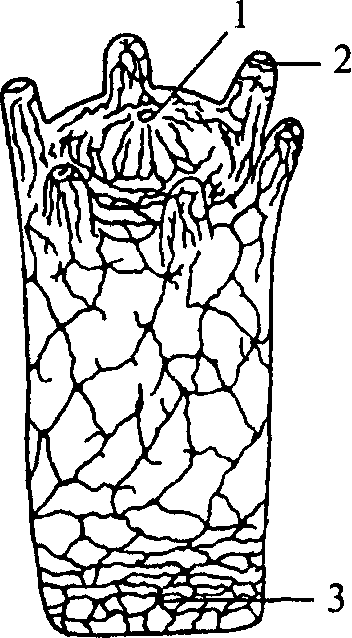
Рис. 9. Схема строения диффузностволовой нервной системы турбеллярии:
1 — нервный узел; 2 — глотка; 3 — брюшной продольный ствол; 4 — боковой нервный ствол
Следующим этапом развития беспозвоночных является появление трехслойных животных — плоских червей. Подобно кишечнополостным они имеют кишечную полость, сообщающуюся с внешней средой ротовым отверстием. Однако у них появляется третий зародышевый слой — мезодерма и двусторонний тип симметрии. Нервная система низших плоских червей принадлежит диффузному типу. Однако из диффузной сети уже обособляются несколько нервных стволов (рис. 9, 3, 4).
У свободно живущих плоских червей нервный аппарат приобретает черты централизации. Нервные элементы собираются в несколько продольных стволов (рис. 10, 4, 5) (для самых высокоорганизованных животных характерно наличие двух стволов), которые соединяются между собой поперечными волокнами (комиссурами) (рис. 10, 6). Упорядоченная таким образом нервная система называется ортогоном. Стволы ортогона представляют собой совокупность нервных клеток и их отростков (рис. 10).
1 — щупальцевидный вырост; 2 — нерв, иннервирующий вырост; 3 — мозговой ганглий; 4 — боковой продольный нервный ствол; 5 — брюшной продольный нервный ствол; 6 — комиссура
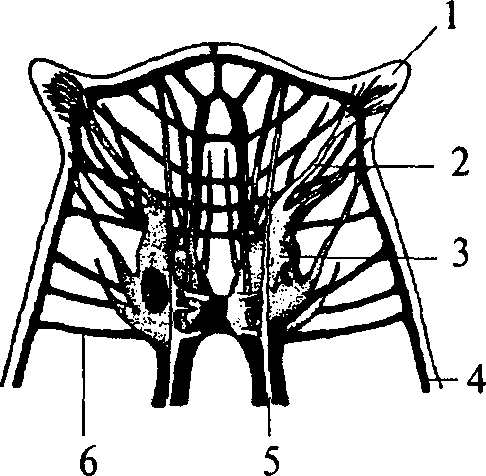
ганглия появляются длинные отростки, идущие в продольные стволы ортогона (рис. 10, 4, 5).
Таким образом, ортогон представляет собой первый шаг к централизации нервного аппарата и его цефализации (появлению мозга). Централизация и цефализация являются результатом развития сенсорных (чувствительных) структур.
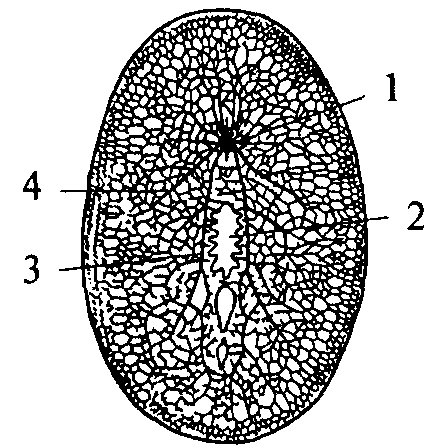
Следующим этапом развития беспозвоночных животных является появление сегментированных животных — кольчатых червей. Их тело метамерно, т.е. состоит из сегментов. Структурной основой нервной системы кольчатых червей является ганглий — парное скопление нервных клеток, расположенных по одному в каждом сегменте. Нервные клетки в ганглии размещаются по периферии. Центральную его часть занимает нейропиль — переплетение отростков нервных клеток и глиальные клетки. Ганглий расположен на брюшной стороне сегмента под кишечной трубкой. Он посылает свои чувствительные и двигательные волокна в свой сегмент и в два соседних. Таким образом, каждый ганглий имеет три пары боковых нервов, каждый из которых является смешанным и иннервирует свой сегмент. Приходящие с периферии чувствительные волокна попадают в ганглий через вентральные корешки нервов. Двигательные волокна выходят из ганглия по дорсальным корешкам нервов. Соответственно этому чувствительные нейроны расположены в вентральной части ганглия, а двигательные — в дорсальной. Кроме того, в ганглии есть мелкие клетки, иннервирующие внутренние органы (вегетативные элементы), они расположены латерально — между чувствительными и двигательными нейронами. Среди нейронов чувствительной, двигательной или ассоциативной зон ганглиев кольчатых червей не обнаружено группирования элементов, нейроны распределены диффузно, т.е. не образуют центров.
Ганглии кольчатых червей соединены между собой в цепочку. Каждый последующий ганглий связан с предыдущим при помощи
1 — надглоточный нервный ганглий;
2 — подглоточный нервный ганглий;
3 — сложный слившийся ганглий грудного сегмента; 4 — брюшной ганглий; 5 — периферический нерв; 6 — коннектива
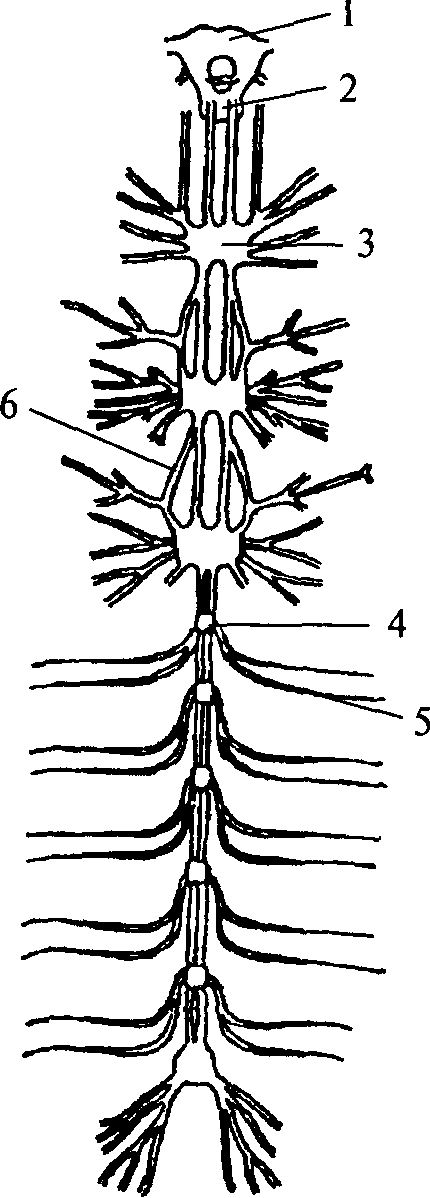
нервных стволов, которые называются коннективами. На переднем конце тела кольчатых червей два слившихся ганглия образуют крупный подглоточный нервный узел. Коннективы от подглоточного нервного узла, огибая глотку, вливаются в надглоточный нервный узел, который является самой ростральной (передней) частью нервной системы. В состав надглоточного нервного ганглия входят только чувствительные и ассоциативные нейроны. Двигательных элементов там не обнаружено. Таким образом, надглоточный ганглий кольчатых червей является высшим ассоциативным центром, он осуществляет контроль над подглоточным ганглием. Подглоточный ганглий контролирует нижележащие узлы, он имеет связи с двумя-тремя последующими ганглиями, тогда как остальные ганглии брюшной нервной цепочки не образуют связей длинней, чем до соседнего ганглия.
В филогенетическом ряду кольчатых червей есть группы с хорошо развитыми органами чувств (многощетинковые черви). У этих животных в надглоточном ганглии обособляются три отдела. Передний отдел иннервирует щупальца, средняя часть иннервирует глаза и антенны. И наконец, задняя часть развивается в связи с совершенствованием химических органов чувств.
Сходную структуру имеет нервная система членистоногих, т.е. построена по типу брюшной нервной цепочки, однако может достигать высокого уровня развития (рис. 11). Она включает в себя значительно развитый надглоточный ганглий, выполняющий функ-
1 — грибовидное тело; 2 — протоцеребрум; 3 — зрительная лопасть; 4 — дейтоцеребрум; 5 — тритоцеребрум
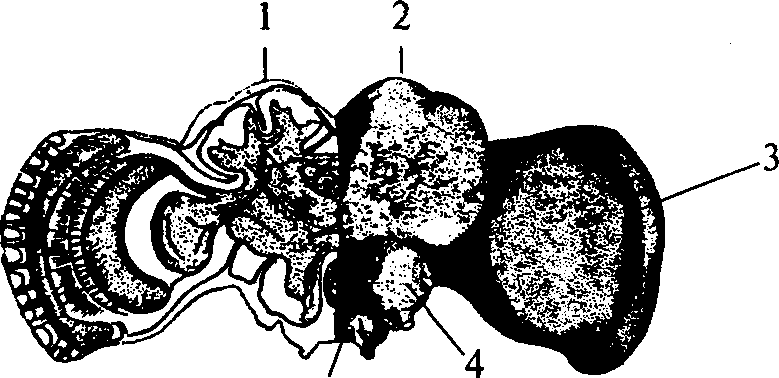
цию мозга, подглоточный ганглий, управляющий органами ротового аппарата, и сегментарные ганглии брюшной нервной цепочки. Ганглии брюшной нервной цепочки могут сливаться между собой, образуя сложные ганглиозные массы.
Головной мозг членистоногих состоит из трех отделов: переднего — протоцеребрума, среднего — дейтоцеребрума и заднего — тритоцеребрума. Сложным строением отличается мозг насекомых. Особенно важными ассоциативными центрами насекомых являются грибовидные тела, располагающиеся на поверхности протоцеребрума, причем чем более сложным поведением характеризуется вид, тем более развиты у него грибовидные тела. Поэтому наибольшего развития грибовидные тела достигают у общественных насекомых (рис. 12).
Практически во всех отделах нервной системы членистоногих существуют нейросекреторные клетки. Нейросекреты играют важную регулирующую роль в гормональных процессах членистоногих.
В процессе эволюции первоначально диффузно расположенные биполярные нейросекреторные клетки воспринимали сигналы либо отростками, либо всей поверхностью клетки, затем сформировались нейросекреторные центры, нейросекреторные тракты и нейросекреторные контактные области. В последующем произошла специализация нервных центров, увеличилась степень надежности во взаимоотношениях двух основных регуляторных систем (нервной и гуморальной) и сформировался принципиально новый этап регуляции — подчинение нейросекреторным центрам периферических эндокринных желез.
1 — церебральная комиссура; 2 — церебральные ганглии; 3 — педальные ганглии; 4 — коннектива; 5 — висцеральные ганглии
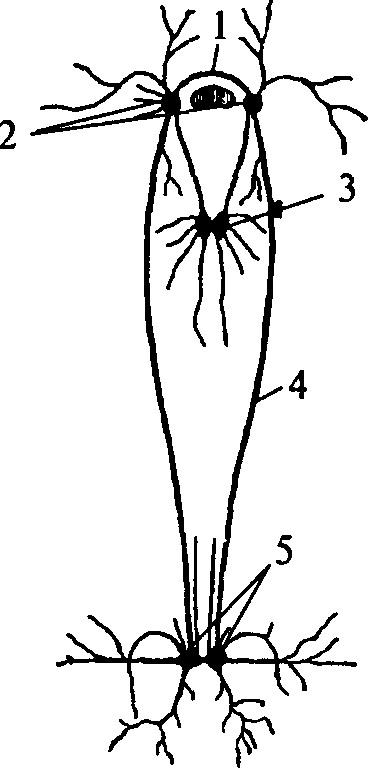
Нервная система моллюсков также имеет ганглионарное строение (рис. 13). У простейших представителей типа она состоит из нескольких пар ганглиев. Каждая пара ганглиев управляет определенной группой органов: ногой, висцеральными органами, легкими и т.д. — и расположена рядом с иннервируемыми органами или внутри их. Одноименные ганглии попарно соединены между собой комиссурами. Кроме того, каждый ганглий связан длинными коннективами с церебральным комплексом ганглиев.
У более высокоорганизованных моллюсков (головоногие) нервная система преобразуется (рис. 14). Ганглии ее сливаются и образуют общую окологлоточную массу — головной мозг. От заднего отдела головного мозга отходят два крупных мантийных нерва и образуют два больших звездчатых ганглия. Таким образом, у головоногих наблюдается высокая степень цефализации.
Группа нервных тканей объединяет ткани эктодермального происхождения, которые в совокупности образуют нервную систему и создают условия для реализации ее многочисленных функций. Обладают двумя основными свойствами: возбудимостью и проводимостью.
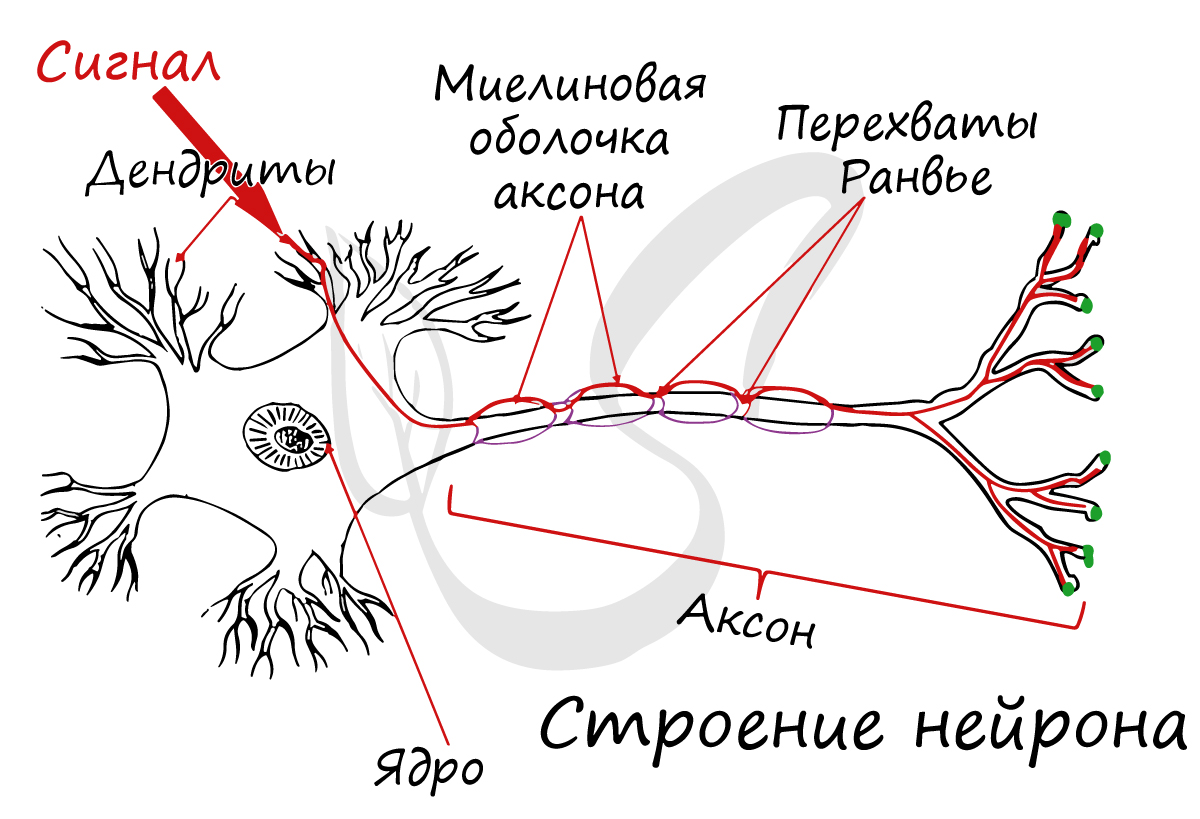
Структурно-функциональной единицей нервной ткани является нейрон (от др.-греч. νεῦρον — волокно, нерв) - клетка с одним длинным отростком - аксоном, и одним/несколькими короткими - дендритами.
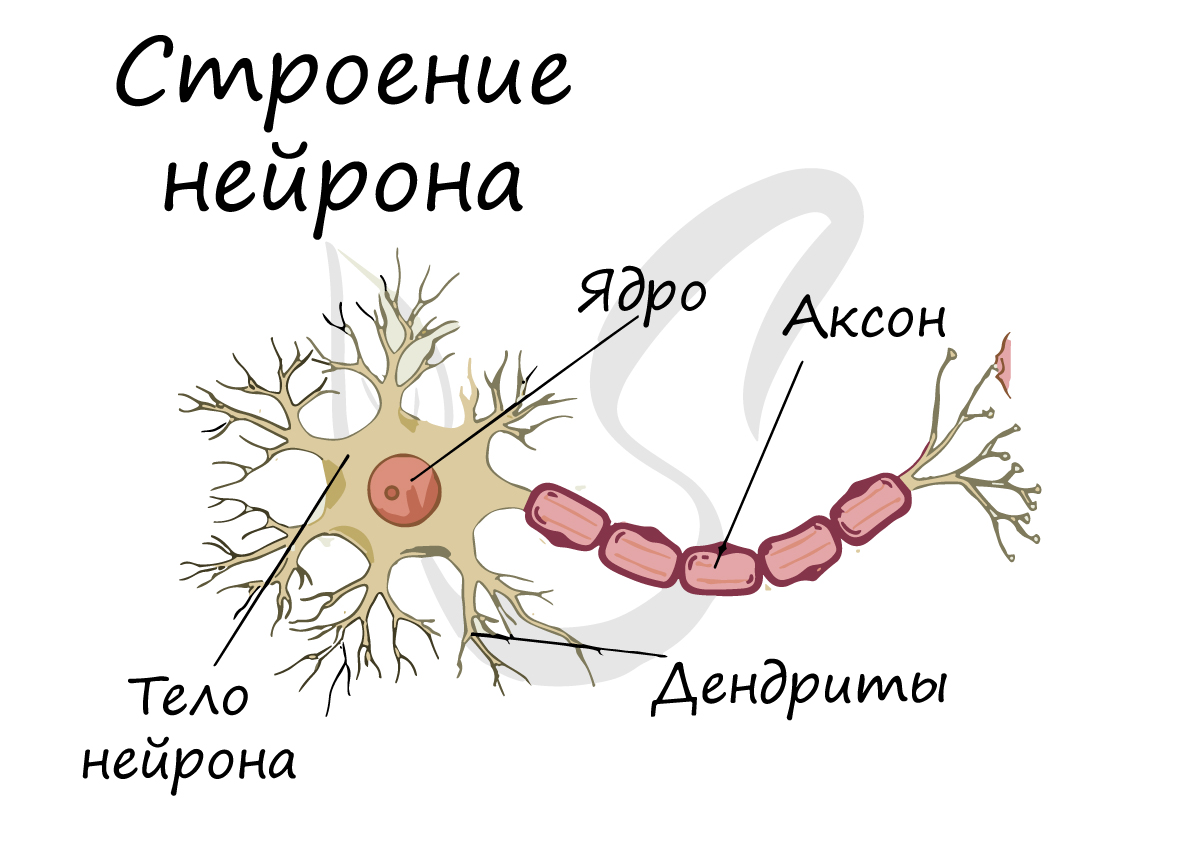
Спешу сообщить, что представление, будто короткий отросток нейрона - дендрит, а длинный - аксон, в корне неверно. С точки зрения физиологии правильнее дать следующие определения: дендрит - отросток нейрона, по которому нервный импульс перемещается к телу нейрона, аксон - отросток нейрона, по которому импульс перемещается от тела нейрона.
Отростки нейронов проводят сгенерированные нервные импульсы и передают их другим нейронам, эффекторам (мышцы, железы), благодаря чему мышцы сокращаются или расслабляются, а секреция желез усиливается или уменьшается.
Отростки нейронов покрыты жироподобным веществом - миелиновой оболочкой, которая обеспечивает изолированное проведение нервного импульса по нерву. Если бы не было миелиновой оболочки (вообразите!) нервные импульсы распространялись бы хаотично, и, когда мы хотели сделать движение рукой, двигалась бы нога.
Существует болезнь, при которой собственные антитела уничтожают миелиновую оболочку (случаются и такие сбои в работе организма.) Эта болезнь - рассеянный склероз, по мере прогрессирования приводит к разрушению не только миелиновой оболочки, но и нервов - а значит, происходит атрофия мышц и человек постепенно становится обездвиженным.

Вы уже убедились, насколько значимы нейроны, их высокая специализация приводит к возникновению особого окружения - нейроглии. Нейроглия - вспомогательная часть нервной системы, которая выполняет ряд важных функций:
- Опорная - поддерживает нейроны в определенном положении
- Изолирующая - ограничивает нейроны от соприкосновения с внутренней средой организма
- Регенераторная - в случае повреждения нервных структур нейроглия способствует регенерации
- Трофическая - с помощью нейроглии осуществляется питание нейронов: напрямую с кровью нейроны не контактируют
В состав нейроглии входят разные клетки, их в десятки раз больше чем самих нейронов. В периферическом отделе нервной системы миелиновая оболочка, изученная нами, образуется именно из нейроглии - шванновских клеток. Между ними хорошо заметны перехваты Ранвье - участки, лишенные миелиновой оболочки, между двумя смежными шванновскими клетками.
Нейроны функционально подразделяются на чувствительные, двигательные и вставочные.
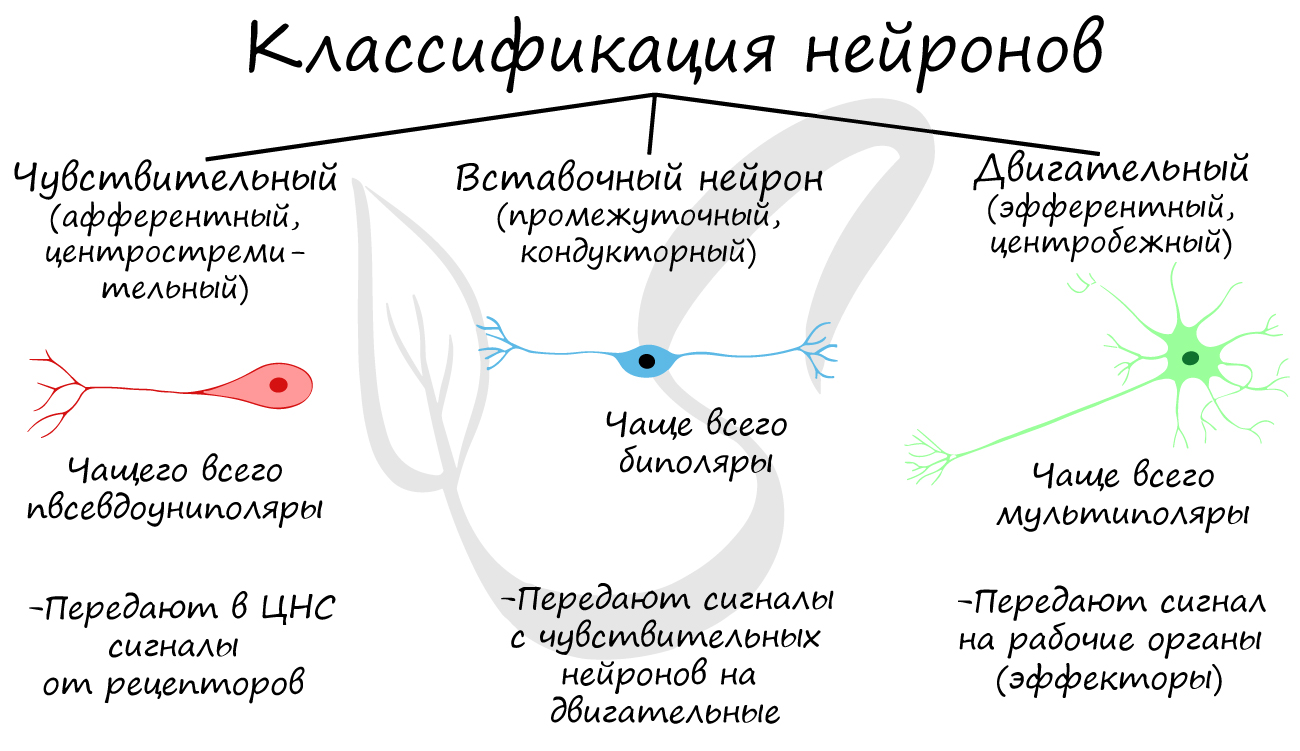
Чувствительные нейроны также называются афферентные, центростремительные, сенсорные, воспринимающие - они передают возбуждение (нервный импульс) от рецепторов в ЦНС. Рецептором называют концевое окончание чувствительных нервных волокон, воспринимающих раздражитель.
Вставочные нейроны также называются промежуточные, ассоциативные - они обеспечивают связь между чувствительными и двигательными нейронами, передают возбуждение в различные отделы ЦНС.
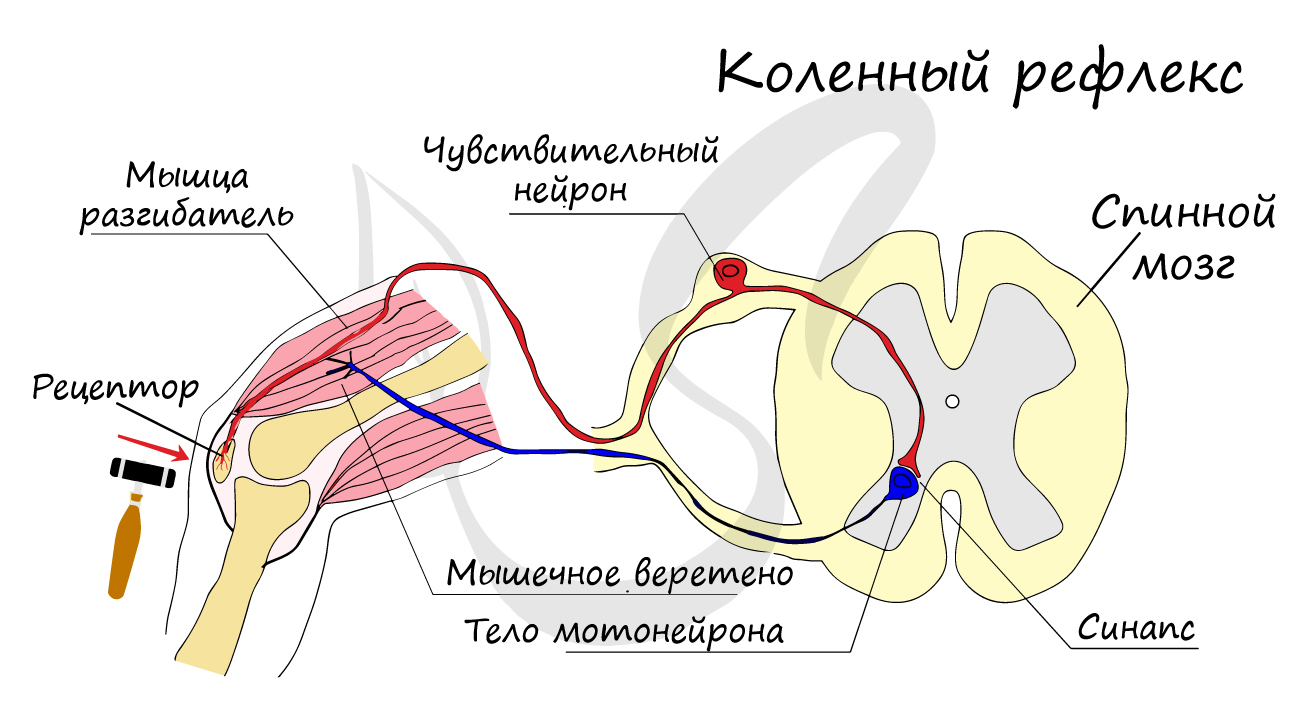
Двигательные нейроны по-другому называются эфферентные, центробежные, мотонейроны - они передают нервный импульс (возбуждение) из ЦНС на эффектор (рабочий орган). Наиболее простой пример взаимодействия нейронов - коленный рефлекс (однако вставочного нейрона на данной схеме нет). Более подробно рефлекторные дуги и их виды мы изучим в разделе, посвященном нервной системе.
На схеме выше вы наверняка заметили новый термин - синапс. Синапсом называют место контакта между двумя нейронами или между нейроном и эффектором (органом-мишенью). В синапсе нервный импульс "преобразуется" в химический: происходит выброс особых веществ - нейромедиаторов (наиболее известный - ацетилхолин) в синаптическую щель.
Разберем строение синапса на схеме. Его составляют пресинаптическая мембрана аксона, рядом с которой расположены везикулы (лат. vesicula — пузырек) с нейромедиатором внутри (ацетилхолином). Если нервный импульс достигает терминали (окончания) аксона, то везикулы начинают сливаться с пресинаптической мембраной: ацетилхолин поступает наружу, в синаптическую щель.
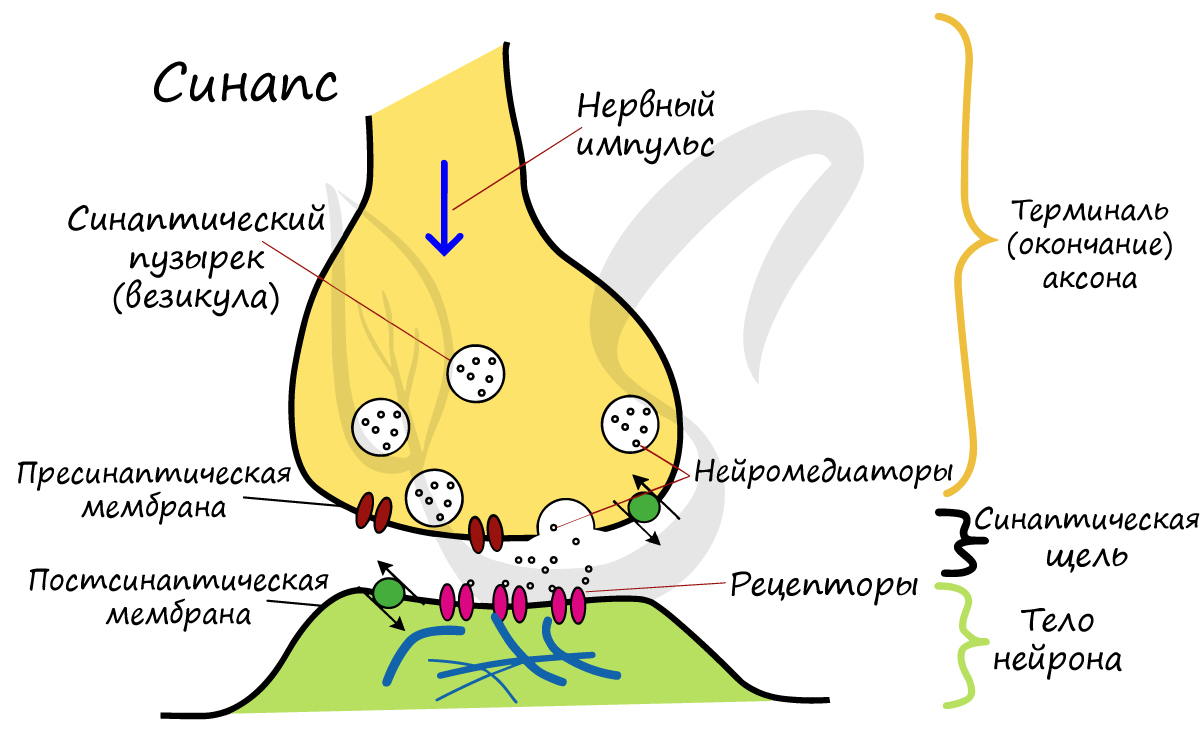
Попав в синаптическую щель, ацетилхолин связывается с рецепторами на постсинаптической мембране, таким образом, возбуждение передается другому нейрону, и он генерирует нервный импульс. Так устроена нервная система: электрический путь передачи сменяется химическим (в синапсе).
Гораздо интереснее изучать любой предмет на примерах, поэтому я постараюсь как можно чаще радовать вас ими ;) Не могу утаить историю о яде кураре, который используют индейцы для охоты с древних времен.
Этот яд блокирует ацетилхолиновые рецепторы на постсинаптической мембране, и, как следствие, химическая передача возбуждения с одного нейрона на другой становится невозможна. Это приводит к тому, что нервные импульсы перестают поступать к мышцам организма, в том числе к дыхательным мышцам (межреберным, диафрагме), вследствие чего дыхание останавливается и наступает смерть животного.

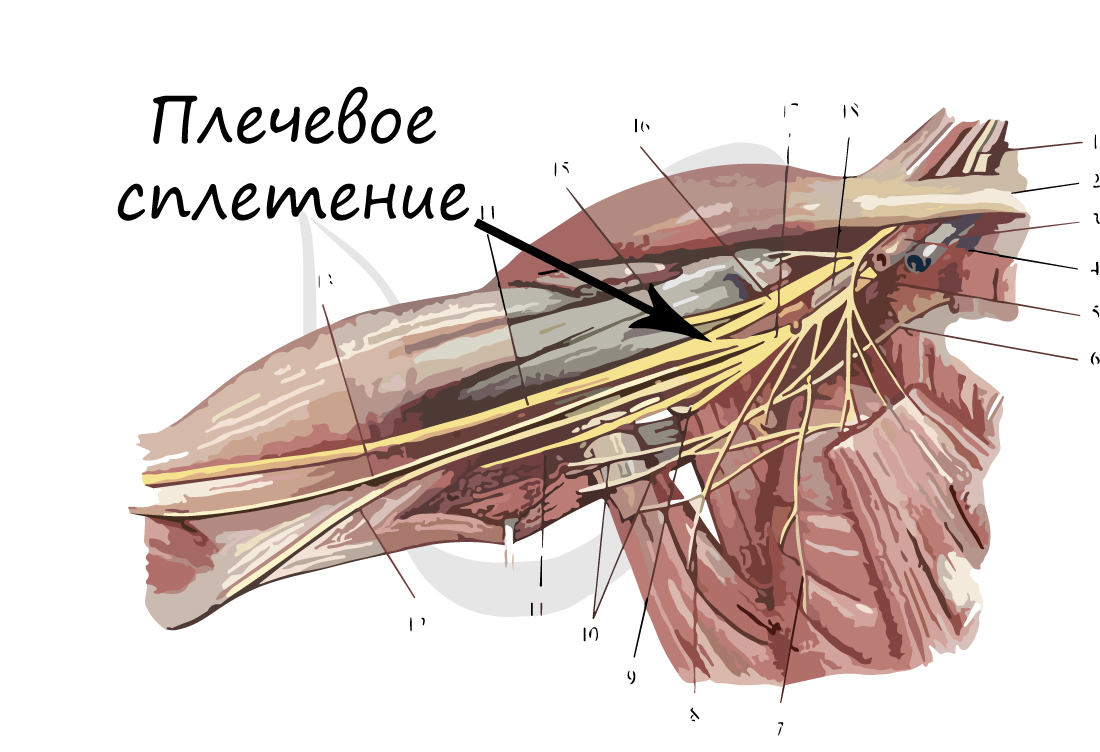
Собираясь вместе, аксоны образуют нервные пучки. Нервные пучки объединяются в нервы, покрытые соединительнотканной оболочкой. В случае, если тела нервных клеток концентрируются в одном месте за пределами центральной нервной системы, их скопления называют нервные узлы - или ганглии (от др.-греч. γάγγλιον — узел).
В случае сложных соединений между нервными волокнами говорят о нервных сплетениях. Одно из наиболее известных - плечевое сплетение.
Неврологические болезни могут развиваться в любой точке нервной системы: от этого будет зависеть клиническая картина. В случае повреждения чувствительного пути пациент перестает чувствовать боль, холод, тепло и другие раздражители в зоне иннервации пораженного нерва, при этом движения сохранены в полном объеме.
Если повреждено двигательное звено, движение в пораженной конечности будет невозможно: возникает паралич, но чувствительность может сохраняться.

Постепенно любые движения мышцами становятся для пациента все труднее, становится тяжело долго говорить, повышается утомляемость. Наблюдается характерный симптом - опущение верхнего века. Болезнь может привести к слабости диафрагмы и дыхательных мышц, вследствие чего дыхание становится невозможным.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Сохранились животные, которые не имеют нервной системы. Это – простейшие одноклеточные организмы и примитивные многоклеточные. У одноклеточных животных функции восприятия выполняют особые участки мембраны клетки (хемочувствительные, фоточувствительные), а моторные функции – специальные органы движения (мембранеллы) типа ресничек, жгутиков и т. п.
У таких сложно организованных одноклеточных организмов, как инфузории, обнаружены внутриклеточные органеллы, выполняющие функции анализа поступающей информации (сенсориум) и организации согласованных движений эффекторов (моториум), а также специальные фибриллы (кинотодесмы), по которым передаются команды от моториума к эффекторам. Т.е. у одноклеточных организмов отсутствует необходимость в существовании специализированных клеток, поскольку сам организм является одноклеточным.
У многоклеточных организмов произошла дифференциация клеток тела по их расположению в теле (наружные, промежуточные и внутренние, выстилающие внутреннюю полость), так и по функции.
Часть поверхностных клеток специализировалась на восприятии внешних воздействий и превратилась в чувствительные (рецепторные). Другие клетки приобрели способность сокращаться и, объединившись, создали мышечные слои. И, наконец, появились клетки, специализированные на передаче возбуждения от рецепторов к мышечным клеткам. Это – нервные клетки. Поскольку мышечные клетки образовали мышечные слои, способные выполнять согласованные сокращения, нервные клетки тоже должны были объединиться в систему для их координации. Так развивается нервная система. Чем более совершенны движения животных, тем лучше развита их нервная система. Неподвижные животные (губки) имеют на поверхности вокруг пор отдельные эпителиально-мышечные клетки, которые способны и воспринимать раздражения, и реагировать на них сокращением.
Тем не менее, диффузная сеть с окологлоточным нервным кольцом была относительно медленно действующим устройством. Измеренная проводимость по нервной сети кишечнополостных составляет не более 5-20 см/с. Этого явно не хватает животным размером более 5 см, поэтому уже у актиний выделились участки нервной сети с высокой скоростью проведения (см. рис. 3, в). В некоторых случаях она достигает 150 см/с, что делает актиний изощрёнными охотниками за значительно более эволюционно продвинутыми позвоночными. Окологлоточное нервное кольцо было явным достижением, но оно не могло дифференциально управлять всем телом или обеспечить контроль за свободным плаванием.
У свободно живущих многоклеточных (кишечнополостные) существует сеть нервных клеток, сгущающаяся в тех местах организма, которые выполняют функцию перемещения тела или захвата и заглатывания пищи. Таким образом, у наиболее примитивного представителя кишечнополостных (пресноводной губки) сеть короткоотростчатых нервных клеток сгущается вблизи подошвы, вокруг щупалец и ротового отверстия. У более сложно организованного кишечнополостного (медузы) сеть уплотняется в тяжи, образующие два кольца вокруг колокола (зонтика). Согласованные сокращения мышц колокола медузы позволяют ей перемещаться в воде по принципу реактивного двигателя.
В нервной сети кишечнополостных нейроны соединяются между собой отростками как протоплазматическим (непрерывным), так и эфаптическим (непосредственный контакт между отростками) способами. Обнаружены и настоящие синаптические контакты. Такая организация нервной системы называется сетевидной, или диффузной.
Так, уиглокожих в центре вокруг пищевода нервные клетки, концентрируясь, образуют три нервных кольца, от которых в каждый луч (у морской звезды) отходят соответственно по три нервных ствола. В стволах имеются и тела нервных клеток, связанных с периферической нервной сетью. Справедливо было бы выделить диффузно-узловой тип организации нервной системы, промежуточный между примитивным диффузным и сложным централизованным. Его развитие связано прежде всего с совершенствованием анализаторной функции.
Уже умедуз можно обнаружить скопления нервных клеток в виде узлов (краевые тельца), выполняющих функцию анализа информации, получаемой от органов чувств (глазков, органа равновесия). В связи с концентрацией органов чувств на переднем конце тела у животных с билатеральным строением тела там же скапливаются нервные клетки в виде узла – "головного мозга". Например, уплоских червей от головного узла к переднему концу тела отходят головные нервы к органам чувств (глазам, органам осязания и т. д.), а к заднему – несколько продольных нервных тяжей ("стволов"), соединяющихся между собой поперечными перемычками. Тяжи состоят из длинных отростков нервных клеток, а также из самих нервных клеток с короткими отростками. Подобную конструкцию называют ортагоном. Такое же строение нервной системы и у круглых червей, но у них тяжи начинаются от окологлоточного кольца.
Централизованная узловая (ганглионарная) нервная система дождевого червя представлена окологлоточным нервным кольцом и брюшной нервной цепочкой. В окологлоточном кольце сильно развитые надглоточные ганглии соединены перемычками с менее развитыми подглоточными узлами; которые, в свою очередь, связаны с нервной цепочкой Цепочка состоит из парных ганглиев, располагающихся в каждом сегменте тела червя Головные надглоточные ганглии переплетаются, в основном, с дистантными рецепторами. Однако дальнейшее развитие церебральных (головных) надглоточных ганглиев зависит от развития интегративной (ассоциативной) функции.
У моллюсков имеется несколько пар ганглиев в "стратегически важных" частях тела:
ü церебральные ганглии, связанные преимущественно с рецепторами головы, расположены в головной части тела;
ü педальные (моторные) ганглии – у начала ноги;
ü висцеральные ганглии – вблизи внутренних органов;
ü плевральные ганглии – между головой и ногой. Ганглии соединены между собой нервными стволами, а с периферией – нервами. У двустворчатых моллюсков нервная система менее развита и состоит лишь из трех пар узлов.
Ø у головоногих моллюсков (кальмары, осьминоги, каракатицы) ЦНС является наиболее сложной:
ü головнойганглий у них стал единственным, а все остальные слились с ним, образовав мощную массу нервной ткани (головной мозг). Головной мозг защищен хрящевой капсулой, имеет множество (до 14) долей. Последние осуществляют связь с хорошо развитыми органами чувств и выполняют анализаторную (сенсорную) функцию:
· подглоточные доли, а также базальные доли надглоточного отдела замыкаются на различных мышцах и определяют моторную функцию,
· задним долям мозга свойственны интегративные (ассоциативные) функции.
Функции головного мозга головоногих моллюсков и позвоночных животных совпадают. Задние доли по своим функциям и свойствам сходны с ассоциативной корой млекопитающих.
ü нижнего (окологлоточного) – тритоцеребрума,
ü среднего – дейтероцеребрума;
У ракообразных ЦНС имеет головные (надглоточные и подглоточные) ганглии и цепочку их в головогруди. У некоторых раков цепочка ганглиев сливается в один крупный узел, у паукообразных вместе с надглоточным ганглием она образует в головогруди сложноорганизованный мозг с большим количеством нервных клеток.
У насекомых нервная система кроме головного мозга имеет цепочку парных ганглиев, которые объединяются в крупные узлы:
ü брюшные узлы выполняют преимущественно висцеральные функции,
ü грудные – моторные функции (организация движения ног и крыльев).
Сенсорные же (анализаторные) и интегративные (ассоциативные) функции контролируются у насекомых головным мозгом.
Впервые в эволюции НС появляется у типа кишечнополостные. Это диффузная НС. Кишечнополостные обладают радиальной (лучевой) симметрией и имеют две жизненные формы — прикрепленные полипы и свободноплавающие медузы. Тело у них состоит из двух слоев клеток — эктодермы и энтодермы. Основу каждого слоя составляют эпителиальномускульные клетки. Таким образом, моторика кишечнополостных бесске- летная, носящая перистальтический характер. Нейроны расположены как в экто-, так и в эктодермальном слоях. Сенсорные функции выполняют специализированные эктодермальные нервные клетки, чувствительные волоски которых выходят на поверхность тела и воспринимают раздражения (см. рис. 16.2, б). Отдельные части тела животных с диффузной НС способны к автономным движениям за счет того, что в каждом участке есть чувствительная клетка, двигательная клетка и нервное волокно. Но даже у самых примитивных кишечнополостных можно найти элементы концентрации — нейроны около рта и на подошве располагаются гуще, чем на остальном теле.
Нервные сети кишечнополостных могут иметь синцитиальное строение, т.е. отростки нервных клеток могут сливаться между собой, проводя возбуждение в разные стороны. Но в большинстве случаев возбуждение передается с помощью типичных синапсов, большинство которых поляризовано, т.е. проводят возбуждение только в одну сторону.
У медуз, которые способны к более активному передвижению, НС несколько усложняется: но краю зонтика у них наблюдаются скопления нервных клеток, которые вместе со своими отростками образуют сплошное нервное кольцо. Таким образом, концентрация нервных элементов у медуз больше, чем у полипов. Более того, у медуз появляются примитивные органы чувств двух типов — статоцисты и глазки — поверхностные скопления светочувствительных клеток. Статоцисты — это органы равновесия, которые можно встретить почти у всех беспозвоночных животных. В большинстве случаев они представляют собой замкнутые пузырьки, на внутренних стенках которых лежат чувствительные клетки. Полость пузырька заполнена жидкостью, в которой находится сгатолит — кристаллик углекислой извести. При изменении положения тела в пространстве чувствительные клетки испытывают давление и возбуждаются.
Животные, следующие в эволюционном развитии за кишечнополостными, — это разные типы червей. Все животные, расположенные на эволюционном древе выше кишечнополостных, — животные трехслойные, проходящие в процессе онтогенеза стадию трех зародышевых листков, которые в своем дальнейшем развитии дают ткани. Таким образом, даже у самых примитивных червей — плоских — уже есть гладкая мышечная ткань, но движения сохраняют перистальтический характер. Концентрация нейронов, только намечавшаяся у медуз, приобретает у червей вполне выраженный характер — появляются нервные ганглии. Но в отличие от кишечнополостных, у которых сокращение мускульных волоконец охватывает все тело, черви способны последовательно сокращать отдельные участки тела, что возможно благодаря иннервации разных участков нейронами, расположенными в разных ганглиях.
Принципиальное отличие червей, как и всех остальных беспозвоночных (за исключением гребневиков и иглокожих) от кишечнополостных, — это наличие билатеральной (двусторонней) симметрии. В связи с этим у них появляется передний конец тела, в сторону которого обычно осуществляется движение. Вследствие этого на нем концентрируются важнейшие органы чувств и, соответственно, передние ганглии укрупняются.
В типе плоские черви, самом примитивном типе червей (например, пла- нария, бычий цепень), централизация нервных клеток еще очень невелика. Их НС состоит из парного головного ганглия и нескольких продольных стволов, соединенных кольцевыми перемычками. Начиная с круглых червей (например, аскарида) у беспозвоночных появляется окологлоточное нервное кольцо, окружающее переднюю часть пищеварительной трубки. Оно обычно состоит из парных надглоточных ганглиев, которые часто сливаются, образуя так называемый головной мозг, и непарного подглоточного ганглия, соединенных нервными волокнами.
В наиболее выраженном виде узловая НС наблюдается в типах кольчатые черви (самые высокоорганизованные черви, например, дождевой червь или нереида), членистоногие и моллюски.
Тело кольчатых червей и членистоногих построено но метамерному принципу и состоит из хорошо выраженных сегментов. От подглоточного узла начинается брюшная нервная цепочка. Исходно (у кольчатых червей) она состоит из парных ганглиев, соединенных комиссурами. Части нервных стволов, соединяющие ганглии каждой стороны соседних сегментов, называются коннективами (см. рис. 16.3, а). Вся цепь имеет характерный вид лестницы (лестничная НС). У разных видов кольчецов наблюдается постепенное сближение и слияние парных ганглиев, при этом коннективы остаются парными. В дальнейшем объединяются и коннективы, и в своем классическом виде брюшная нервная цепочка выглядит как на рис. 16.3, б.
Расположение ганглиев на брюшной стороне связано, по-видимому, с расположением основных рецептивных областей. Черви передвигаются путем ползания и получают многие сенсорные импульсы от вентральной поверхности. В передней же части тела наибольшее значение имеет рецепция, получаемая от рострального конца, в связи с чем головной ганглий поднимается вверх.
НС членистоногих (ракообразные, паукообразные, насекомые) устроена значительно совершеннее, чем у кольчатых червей. Это связано как с более сложным поведением, так и с особенностями организации. Для этих животных характерна гетерономная сегментация, т.е. сегменты тела не одинаковы, как у кольчатых червей, а отличаются друг от друга как по строению, так и по функциям. Конечности состоят из нескольких члеников, подвижно соединенных друг с другом и с телом с помощью суставов. Следовательно, принципиально меняется характер движения — он становится рычаговым. Помимо этого, большая часть гладкой мускулатуры заменяется поперечнополосатой (впервые в эволюции), что способствует гораздо более быстрым движениям. Что касается НС, то концентрация и централизация нейронов (если говорить о беспозвоночных) достигает своего максимума. Нередко наблюдается слияние нервных узлов брюшной нервной цепочки с образованием более крупных ганглиев, т.е. наблюдается и продольная концентрация элементов НС.
Но особенно сильно НС членистоногих отличается от НС червей в строении ее головного конца. У большинства этих животных ГМ (надглоточный ганглий) состоит из трех отделов: переднего — протоцеребрума, среднего — дейтоцеребрума и заднего — тритоцеребрума. Каждый отдел головного мозга иннервирует разные органы чувств и состоит из нескольких ганглиев, отделенных друг от друга белым веществом. В протоцере- бруме расположены особенно важные ассоциативные центры — грибовидные тела (рис. 16.5).
Наибольшее развитие головного мозга наблюдается у общественных насекомых (муравьев, пчел, термитов), т.е. у животных с особенно сложным поведением. Причем даже в пределах одного вида величина мозга может значительно варьироваться в зависимости от сложности поведения. Так, например, у рабочих муравьев грибовидные тела развиты значительно сильнее, чем у цариц и самцов.
Что касается моллюсков, то это резко отграниченный тип беспозвоночных, ведущий начало от древних кольчатых червей, но имеющий несегмен- тированное тело. От окологлоточного кольца у моллюсков отходят четыре продольных нервных ствола. У высших форм моллюсков на стволах формируются парные ганглии, соединенные коннективами, некоторые ганглии соединяются и с помощью комиссур (рис. 16.6). Нервные клетки имеются не только в ганглиях, но и рассеяны в периферических нервах. Такой тип нервной системы называют разбросанно-узловым.

Рис. 16.5. Нервная система насекомого

Рис. 16.6. Нервная система брюхоногого моллюска
У высших моллюсков крупные ганглии расположены, как правило, рядом с иннервируемым органом или непосредственно в его стенках. Надглоточные ганглии иннервируют глаза и статоцисты, глотку и щупальца; ганглии, иннервирующие мускулатуру ноги, расположены в самой ноге; рядом с ними лежат ганглии, иннервирующие мантию; следующая пара ганглиев связана с жабрами и органами химического чувства; еще одна пара управляет внутренними органами.
Особенно высокой сложности строения НС достигает у головоногих моллюсков (рис. 16.7).

Рис. 16.7. Головной мозг головоногого моллюска
Большинство этих животных — свободноплавающие и очень подвижные хищники. Для них характерно настолько сложное поведение, что их иногда называют приматами моря. Ганглии у них очень велики и образуют сплошную окологлоточную нервную массу (ГМ). В этой общей массе выделяют отдельные ганглии, выполняющие различные функции. В мозге головоногих даже найдены участки (в зрительной доле), имеющие слоистое строение. Для защиты такой НС головоногие имеют подкожную хрящевую капсулу, окружающую мозг аналогично черепу позвоночных.
Читайте также:
