
Как образованы миелинизированные нервные волокна
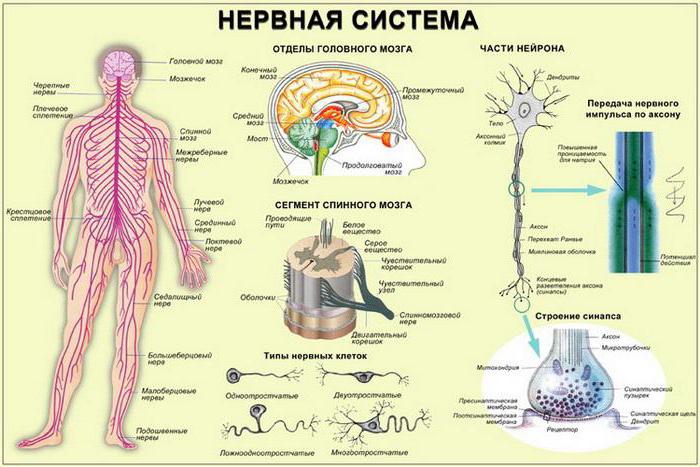
Нервная система человека и позвоночных животных имеет единый план строения и представлена центральной частью – головным и спинным мозгом, а также периферическим отделом – отходящими от центральных органов нервами, представляющими собой отростки нервных клеток – нейронов.
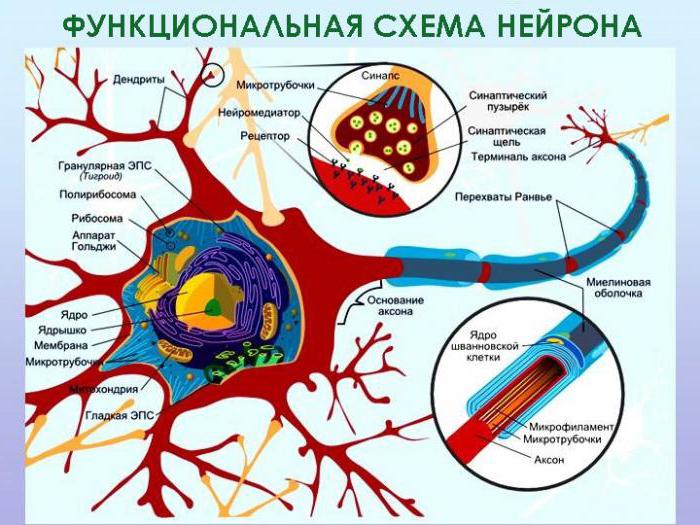
Их совокупность образует нервную ткань, главными функциями которой является возбудимость и проводимость. Эти её свойства объясняются прежде всего особенностями строения оболочек нейронов и их отростков, состоящих из вещества, названного миелином. В данной статье мы рассмотрим строение и функции этого соединения, а также выясним возможные способы его восстановления.
Почему нейроциты и их отростки покрыты миелином
Совсем не случайно дендриты и аксоны имеют защитный слой, состоящий из белково-липидных комплексов. Дело в том, что возбуждение является биофизическим процессом, в основе которого лежат слабые электрические импульсы. Если электрический ток идёт по проводу, то последний должен быть покрыт изоляционным материалом, чтобы уменьшить рассеивание электрических импульсов и не допустить снижение силы тока. Такие же функции в нервном волокне выполняет миелиновая оболочка. Кроме того, она является опорой, а также обеспечивает питание волокна.
Химический состав миелина
Как и большинство клеточных мембран, он имеет липопротеидную природу. Причём содержание жиров здесь очень высокое – до 75%, а белков – до 25%. Миелин в незначительном количестве содержит также гликолипиды и гликопротеиды. Химический состав его различается в спинномозговых и в черепно-мозговых нервах.
В первых наблюдается высокое содержание фосфолипидов – до 45%, а остальная часть приходится на холестерин и цереброзиды. Демиелинизация (то есть замена миелина на другие вещества в нервных отростках) приводит к таким тяжёлым аутоиммунным заболеваниям, как, например, рассеянный склероз.
С химической точки зрения, этот процесс будет выглядеть так: миелиновая оболочка нервных волокон меняет свою структуру, что проявляется прежде всего в уменьшении процентного содержания липидов по отношению к белкам. Далее снижается количество холестерина и возрастает содержание воды. А всё это приводит к постепенной замене миелина, содержащего олигодендроциты или шванновские клетки на макрофаги, астроциты и межклеточную жидкость.
Результатом таких биохимических изменений будет резкое снижение способности аксонов проводить возбуждение вплоть до полной блокировки прохождения нервных импульсов.
Особенности нейроглиальных клеток
Как мы уже говорили, миелиновая оболочка дендритов и аксонов образована специальными структурами, характеризующимися низкой степенью проницаемости для ионов натрия и кальция, а потому имеющих только потенциалы покоя (они не могут проводить нервные импульсы и выполняют электроизоляционные функции).
Данные структуры называются глиальными клетками. К ним относятся:
- олигодендроциты;
- волокнистые астроциты;
- клетки эпендимы;
- плазматические астроциты.
Все они формируются из наружного слоя зародыша – эктодермы и имеют общее название – макроглия. Глия симпатических, парасимпатических и соматических нервов представлена шванновскими клетками (нейролеммоцитами).
Строение и функции олигодендроцитов
Они входят в состав центральной нервной системы и являются клетками макроглии. Так как миелин – это белково-липидная структура, она способствует увеличению скорости проведения возбуждения. Сами клетки образуют электроизолирующий слой нервных окончаний в головном и спинном мозге, формируясь уже в период внутриутробного развития. Их отростки обворачивают в складки своей наружной плазмалеммы нейроны, а также дендриты и аксоны. Получается, что миелин – это основной электроизолирующий материал, разграничивающий нервные отростки смешанных нервов.
Шванновские клетки и их особенности
Миелиновая оболочка нервов периферической системы образована нейролеммоцитами (шванновскими клетками). Их отличительная особенность состоит в том, что они способны образовывать защитную оболочку только одного аксона, и не могут формировать отростки, как это присуще олигодендроцитам.
Между шванновскими клетками на расстоянии 1-2 мм располагаются участки, лишённые миелина, так называемые перехваты Ранвье. По ним скачкообразно происходит проведение электрических импульсов в пределах аксона.
Леммоциты способны к репарации нервных волокон, а также выполняют трофическую функцию. В результате генетических аббераций клетки оболочки леммоцитов начинают неконтролируемое митотическое деление и рост, вследствие чего в различных отделах нервной системы развиваются опухоли – шванномы (невриномы).
Роль микроглии в разрушении миелиновой структуры
Микроглия представляет собой макрофаги, способные к фагоцитозу и умеющие распознавать различные патогенные частицы – антигены. Благодаря мембранным рецепторам эти глиальные клетки вырабатывают ферменты – протеазы, а также цитокины, например, интерлейкин 1. Он является медиатором воспалительного процесса и иммунитета.
Более того, цитокины, активируя рецепторы, провоцируют избыточный транспорт ионов кальция в тело нейрона. Протеазы и фосфолипазы начинают расщеплять органеллы и отростки нервных клеток, что приводит к апоптозу – гибели данной структуры.
Она разрушается, распадаясь на частицы, которые и пожирают макрофаги. Это явление называется эксайтотоксичностью. Оно вызывает дегенерацию нейронов и их окончаний, приводя к таким заболеваниям, как болезнь Альцгеймера и болезнь Паркинсона.
Мякотные нервные волокна
Если отростки нейронов – дендриты и аксоны, покрывает миелиновая оболочка, то они называются мякотными и иннервируют скелетную мускулатуру, входя в соматический отдел периферической нервной системы. Немиелинизированные волокна образуют вегетативную нервную систему и иннервируют внутренние органы.
Мякотные отростки имеют больший диаметр, чем безмякотные, и формируются следующим образом: аксоны прогибают плазматическую мембрану клеток глии и формируют линейные мезаксоны. Затем они удлиняются и шванновские клетки многократно обворачиваются вокруг аксона, образуя концентрические слои. Цитоплазма и ядро леммоцита перемещаются в область наружного слоя, который называется неврилеммой или шванновской оболочкой.
Внутренний слой леммоцита состоит из слоистого мезоксона и называется миелиновой оболочкой. Толщина её в различных участках нерва неодинакова.
Как восстановить миелиновую оболочку
Рассматривая роль микроглии в процессе демиелинизации нервов, мы установили, что под действием макрофагов и нейромедиаторов (например, интерлейкинов) происходит разрушение миелина, что в свою очередь приводит к ухудшению питания нейронов и нарушению передачи нервных импульсов по аксонам.
Данная патология провоцирует возникновение нейродегенеративных явлений: ухудшение когнитивных процессов, прежде всего памяти и мышления, появление нарушения координации движений тела и тонкой моторики.
В итоге возможна полная инвалидизация больного, которая возникает в результате аутоиммунных заболеваний. Поэтому вопрос о том, как восстановить миелин, в настоящее время стоит особенно остро. К таким способам относится прежде всего сбалансированная белково-липидная диета, правильный образ жизни, отсутствие вредных привычек. В тяжелых случаях заболеваний применяют медикаментозное лечение, восстанавливающее количество зрелых глиальных клеток – олигодендроцитов.
Состоят из отростка нервной клетки, покрытого оболочкой, которая формируется олигодендроцитами. Отросток нервной клетки (аксон или дендрит) в составе нервного волокна называется осевым цилиндром.
- безмиелиновое (безмякотное) нервное волокно,
- миелиновое (мякотное) нервное волокно.
Безмиелиновые нервные волокна
Находятся преимущественно в составе вегетативной нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи, в которых на определенном расстоянии друг от друга видны овальные ядра. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько (10—20) осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в смежное. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. При электронной микроскопии безмиелиновых нервных волокон видно, что по мере погружения осевых цилиндров в тяж неиролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне
29. Миелиновые нервные волокна
1) внутренний, более толстый, — миелиновый слой,
2) наружный, тонкий, состоящий из цитоплазмы, ядер нейролеммоцитов и нейролеммы.
Миелиновый слой содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии — насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы видны участки волокна, лишенные миелинового слоя, — узловатые перехваты, или перехваты Ранвье, т.е. границы между соседними леммоцитами.
Отрезок волокна между смежными перехватами называется межузловым сегментом.
В процессе развития аксон погружается в желобок на поверхности нейролеммоцита. Края желобка смыкаются. При этом образуется двойная складка плазмолеммы нейролеммоцита — мезаксон. Мезаксон удлиняется, концентрически наслаивается на осевой цилиндр и образует вокруг него плотную слоистую зону — миелиновый слой. Цитоплазма с ядрами отодвигается на периферию – образуется наружная оболочка или светлая Шванновская оболочка (при окраске осмиевой кислотой).
Осевой цилиндр состоит из нейроплазмы, продольных параллельных нейрофиламентов, митохондрий. С поверхности покрыт мембраной – аксолеммой, обеспечивающей проведение нервного импульса. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Нервный импульс в миелиновом нервном волокне проводится как волна деполяризации цитолеммы осевого цилиндра, "прыгающая" (сальтирующая) от перехвата к следующему перехвату со скоростью до 120 м/сек.
В случае повреждения только отростка нейроцита регенерация возможна и протекает успешно при наличии определенных для этого условий. При этом, дистальнее места повреждения осевой цилиндр нервного волокна подвергается деструкции и рассасывается, но леммоциты при этом остаются жизнеспособными. Свободный конец осевого цилиндра выше места повреждения утолщается - образуется "колба роста", и начинает расти со скоростью 1 мм/день вдоль оставшихся в живых леммоцитов поврежденного нервного волокна, т.е. эти леммоциты играют роль "проводника" для растущего осевого цилиндра. При благоприятных условиях растущий осевой цилиндр достигает бывшего рецепторного или эффекторного концевого аппарата и формирует новый концевой аппарат.
30. шванновские клетки (леммоциты) — вспомогательные клетки нервной ткани, которые формируются вдоль аксонов периферических нервных волокон. Создают, а иногда и разрушают, электроизолирующую миелиновую оболочку нейронов. Выполняют опорную (поддерживают аксон) и трофическую (питают тело нейрона) функции. Описаны немецким физиологом Теодором Шванном в 1838 году и названы в его честь.
Каждое периферическое нервное волокно одето тонким цитоплазматическим слоем — невролеммой или шванновской оболочкой. Волокно является миелинизированным, если между ним и цитоплазмой шванновской клетки имеется значительный слой миелина. Если волокна лишены миелина, то они называются немиелинизированными безмякотными. Шванновские клетки могут осуществлять волнообразные движения, что, вероятно, способствует транспортировке различных веществ по отросткам нервных клеток.
С нарушением работы шванновских клеток связаны такие нервные заболевания, как синдром Гийена — Барре, болезнь Шарко-Мари, шванноматозис и хроническая воспалительная демиелинизирующая полинейропатия. Демиелинизация в основном происходит из-за ослабления двигательных функций шванновских клеток, в результате чего они оказываются не способны образовывать миелиновую оболочку.
Особенности распространения возбуждения по безмиелиновым волокнам:
1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением.
2. Возбуждение распространяется с небольшой скоростью.
3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна).
По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров.
Однако низкая скорость распространения возбуждения и его затухание не всегда выгодно организму. Поэтому природой был выработан еще один дополнительный механизм распространения возбуждения.
32. Механизмы проведения возбуждения в миелиновых волокнах. Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. В миелинизированномволокне токи проводятся только в зонах, не покрытых миелином (перехватах Ранвье). В этих участках генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением.
При действии раздражителя пороговой силы на мембрану миелинового волокна в области перехвата Ранвье изменяется проницаемость для ионов Nа + , которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны, что ведет к возникновению круговых токов. Этот ток идет через межтканевую жидкость к соседнему перехвату, где происходит смена заряда. Таким образом, возбуждение перепрыгивает с одного участка на другой. Обратное движение возбуждения невозможно так как участок, через который оно прошло, находится в фазе абсолютной рефрактерности.
Особенности распространения возбуждения по миелиновым волокнам:
2. Возбуждение распространяется с большой скоростью.
3. Возбуждение распространяется без декримента.
По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции.
Сальтаторное проведение
(лат. saltatorius, от salto — скачу, прыгаю)
Сальтаторное распространение возбуждения в миелинизированном нервном волокне от перехвата к перехвату [стрелками показано направление тока, возникающего между возбуждённым (А) и соседним покоящимся (Б) перехватом].
34. Проведение нервного импульса, передача сигнала в виде волны возбуждения в пределах одного нейрона и от одной клетки к другой. П. н. и. по нервным проводникам происходит с помощью электротонических потенциалов и потенциалов действия, которые распространяются вдоль волокна в обоих направлениях, не переходя на соседние волокна (см. Биоэлектрические потенциалы, Импульс нервный). Передача межклеточных сигналов осуществляется через синапсы чаще всего с помощью медиаторов, вызывающих появление потенциалов постсинаптических. Нервные проводники можно рассматривать как кабели, обладающие относительно низким осевым сопротивлением (сопротивление аксоплазмы — ri) и более высоким сопротивлением оболочки (сопротивление мембраны — rm). Нервный импульс распространяетсявдоль нервного проводника посредством прохождения тока между покоящимися и активными участками нерва (локальные токи). В проводнике по мере увеличения расстояния от места возникновения возбуждения происходит постепенное, а в случае однородной структуры проводника экспоненциальное затухание импульса, который в 2,7 раза уменьшается на расстоянии l = (константа длины). Так как rm и riнаходятся в обратном отношении к диаметру проводника, то затухание нервного импульса в тонких волокнах происходит раньше, чем в толстых. Несовершенство кабельных свойств нервных проводников восполняется тем, что они обладают возбудимостью. Основное условие возбуждения — наличие у нервов потенциала покоя. Если локальный ток через покоящийся участок вызовет деполяризацию мембраны, достигающую критического уровня (порога), это приведёт к возникновению распространяющегося потенциала действия (ПД). Соотношение уровня пороговой деполяризации и амплитуды ПД, обычно составляющее не менее 1: 5, обеспечивает высокую надёжность проведения: участки проводника, обладающие способностью генерировать ПД, могут отстоять друг от друга на таком расстоянии, преодолевая которое нервный импульс снижает свою амплитуду почти в 5 раз. Этот ослабленный сигнал будет снова усилен до стандартного уровня (амплитуда ПД) и сможет продолжить свой путь по нерву.
Строение безмиелиновых нервных волокон. Примеры их локализации.
Структурные компоненты нервных волокон
Отростки нейроцитов почти всегда покрыты оболочками. Исключение составляют свободные окончания некоторых отростков. Отросток нейрона вместе с оболочкой называется нервным волокном. Сам же отросток нейрона, находящийся в составе волокна, называется осевым цилиндром. Оболочки в нервном волокне образованы олигодендроцитами, которые в случае п.н.с. называются шванновскими клетками (или леммоцитами). По своему строению нервные волокна подразделяются на 2 типа - безмиелиновые (безмякотные) и миелиновые (мякотные).
| Безмиелиновые волокна | Миелиновые волокна |
| 1. Несколько осевых цилиндров, располагающихся по периферии волокна. | 1. Один осевой цилиндр находится в центре волокна. |
| 2. Осевые цилиндры - это, как правило, аксоны эфферентных нейронов вегетативной нервной системы. | 2. Осевой цилиндр может быть как аксоном, так и дендритом нейроцита. |
| 3. Ядра олигодендроцитов находятся в центре волокон. | 3. Ядра и цитоплазма леммоцитов оттеснены к периферии волокна. |
| 4. Мезаксоны осевых цилиндров - короткие. | 4. Мезаксон многократно закручивается вокруг осевого цилиндра, образуя миелиновый слой. |
| 5. Na + -каналы располагаются по всей длине осевого цилиндра. | 5. Na + -каналы - только в перехвате Ранвье |
Безмиелиновые волокна находятся: преимущественно - в составе вегетативной нервной системы, где содержат, главным образом, аксоны эффекторных нейронов этой системы; в меньшей степени - в ЦНС.
На поперечном сечении при электронной микроскопии:
· В центре располагается ядро олигодендроцита (леммоцита).
· По периферии в цитоплазму погружено обычно несколько (10-20) осевых цилиндров.
· При погружении осевого цилиндра в цитоплазму глиоцита плазмолемма сближается над цилиндром, образуя "брыжейку" последнего – мезаксон
· С поверхности нервное волокно покрыто базальной мембраной .
По длине волокна олигодендроциты (леммоциты) соединяются друг с другом конец в конец, образуя непрерывный тяж.
В ЦНС и в соматических отделах ПНС. Они могут содержать как аксоны, так и дендриты нервных клеток.
На поперечном сечении:
· Всего один и располагается в центре.
· 2 слоя: внутренний - миелиновый слой и наружный – нейролемма.
o Миелиновый слой
§ Несколько слоев мембраны олигодендроцита, концентрически закрученных вокруг осевого цилиндра.
§ это удлинённый мезаксон, образующийся при погружении осевого цилиндра в цитоплазму глиоцита и последующем вращении цилиндра вокруг своей оси.
o Нейролемма - это оттеснённые к периферии (т.е. кнаружи от миелинового слоя) цитоплазма и ядро глиоцита.
· Снаружи волокно в периферическом нерве покрыто базальной мембраной.
· В ЦНС миелиновые волокна имеют ряд особенностей:
o один олигодендроцит с помощью несколько отростков участвует в образовании оболочки сразу нескольких соседних волокон;
o у миелина - специфический липопротеидный состав, вокруг волокна нет базальной мембраны.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Они, как и немиелинизиро- ванные, окружены глиальными клетками (они называются шванновские клетки), но мембраны этих клеток плотно прилегают к мембране нервного волокна. Сами шванновские клетки уплощаются, охватывают аксон и многократно закручиваются вокруг него наподобие изоляции электрического кабеля. Соприкасающиеся мембраны шванновской клетки смыкаются, образуя плотные пластинки — мезаксон. Смыкание и образование мезак- сона происходит за счет взаимодействия белков, примыкающих к внутренней стороне мембраны.
Белки наружной стороны мембраны тоже взаимодействуют, образуя неплотные пластинки, которые чередуются с плотными. В зависимости от диаметра аксона оболочка, образованная вокруг нервного волокна шванновской клеткой, может содержать от 10 до 200 мембранных слоев. При этом всегда сохраняется сома шванновской клетки, содержащая основные органеллы. Принципиальное строение миелинизированного нервного волокна показано на рис. 2.22. Миелиновая оболочка, таким образом, представляет собой совокупность мембран шванновских клеток. Основным компонентом мембран являются фосфолипиды (с высоким содержанием сфингомиелина), обладающие хорошими изоляционными свойствами, т.е. высоким электрическим сопротивлением.

Рис. 2.22. Схема строения миелинизированного нервного волокна
Каждая шванновская клетка, накрученная на аксон, создает по ходу аксона миелинизированный участок длиной 1—2 мм. Между последовательно расположенными шванновскими клетками всегда остается неизолированная (немиелинизированная) область волокна длиной 2—3 мкм, где ионы могут свободно переходить через мембрану из внеклеточной жидкости в аксоплазму и обратно. Эту область аксона называют перехватом Ран- вье. Таким образом, мембрана аксона состоит из регулярно чередующихся миелинизированных (межперехватных) участков длиной 1—2 мм и перехватов Ранвье длиной 2—3 мкм (см. рис. 2.22). В ЦНС миелинизированные нервные волокна выглядят так же, как в периферических нервах. Единственная особенность состоит в том, что в ЦНС одна клетка глии (оли- годендроцит) способна давать отростки к нескольким аксонам, формируя миелиповую оболочку вокруг каждого из них.

Рис. 2.23. Схема сальтаторного распространения ПД по миелинизированному нервному волокну:
ПД, возникший в одном возбужденном перехвате Ранвье, вызывает развитие локальных токов, замыкающихся лишь в следующем перехвате, где происходят деполяризация мембраны и генерация следующего ПД
Между таким возбужденным и соседними невозбужденными миелини- зированными участками мембраны возникает разность потенциалов. Эта разность порождает местные электрические токи, но они не могут выходить наружу через миелиновую оболочку ввиду ее высокого сопротивления. Поэтому нерастраченные утечкой в наружную среду локальные токи текут дальше внутри аксона по аксоплазме до соседнего невозбужденного перехвата Ранвье (см. рис. 2.23). Только там они могут проходить через мембрану, гасить ее электронегативный заряд и замыкаться.
Наряду с высоким фактором надежности сальтаторпое проведение ПД имеет еще ряд преимуществ по сравнению с непрерывным. Скачкообразная генерация ПД повышает скорость проведения возбуждения в миелинизированных волокнах в 5—50 раз. Действительно, длина межиерехватных участков составляет около 2 мм, а перехватов Ранвье — 1—2 мкм. С учетом того, что возбуждение может возникать не в следующем, а во втором или третьем перехвате, получается, что ПД распространяется по волокну скачками длиной 2—4 мм. Кроме того, сальтаторное проведение возбуждения экономит энергию для аксона. В миелинизированных волокнах деполяризуются только перехваты, что позволяет приблизительно в 100 раз снизить потерю ионов. В связи с этим снижаются траты энергии, необходимые для восстановления трансмембранной разности концентраций ионов натрия и калия после проведения серии нервных импульсов. Наконец, в крупных миелинизированных волокнах имеется еще одна особенность сальтаторного проведения: высокая изоляция миелиновой оболочкой в сочетании с 50-кратно сниженной электрической емкостью межперех- ватной мембраны позволяет осуществлять реполяризацию ПД путем перемещения очень незначительного числа ионов.
Важнейшие закономерности процесса распространения возбуждения по нервным волокнам следующие:
- 1) потенциал действия распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
- 2) генерация ПД нервными волокнами практически не вызывает их утомления;
- 3) нервные волокна обладают высокой лабильностью, т.е. могут воспроизводить потенциал действия с очень высокой частотой;
- 4) расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
- 5) распространение потенциала действия — активный процесс, в ходе которого изменяется состояние ионных каналов мембраны волокна, а также расходуется энергия гидролиза АТФ для восстановления трансмембранных ионных градиентов;
- 6) ПД распространяется по каждому нервному волокну изолированно — не переходит с одного волокна на другое. Это обусловлено значительно более низким сопротивлением межклеточной жидкости по сравнению с сопротивлением мембраны волокон. Из-за этого текущие между возбужденным и невозбужденным участками наружные местные токи проходят в основном по межклеточной жидкости, не затекая и не действуя на другие волокна;
- 7) проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность. Фактор надежности проведения сигнала у миелинизированных нервных волокон выше, чем у немиелинизированных.
Гистологический препарат № 11
Миелиновые нервные волокна. Узловой перехват. Насечки.
Импрегнация серебром. Большое увеличение. Найти:
- миелиновое нервное волокно и в нем:
- осевой цилиндр,
- миелиновый слой оболочки,
- насечки миелина,
- наружный слой оболочки,
- ядра леммоцитов,
- узловые перехваты.
Гистологический препарат. Безмиелиновые нервные волокна.
Окраска гематоксилином и эозином. Увеличение большое. Найти:
- безмиелиновое нервное волокно и в нем:
- осевой цилиндр,
- ядра лиммоцитов.
Нервные волокна (neurofibrae) бывают двух видов: миелиновые и безмиелиновые. Оба типа нервных волокон имеют единый план строения и представляют собой отростки нервных клеток (осевые цилиндры), окруженные оболочкой из олигодендроглии-леммоцитов (шванновских клеток). С поверхности к каждому волокну примыкает базальная мембрана с прилегающими к ней коллагеновыми волокнами.
Миелиновые волокна (neurofibrae myelinafae) имеют относительно больший диаметр, сложно устроенную оболочку из леммоцитов и большую скорость проведения нервного импульса (15 — 120 м/сек). В оболочке миелинового волокна выделяют два слоя; внутренний, миелиновый (stratum myelini), более толстый, содержащий много липидов и окрашивающийся осмием в черный цвет. Он состоит из плотноупакованных по спирали вокруг осевого цилиндра слоев-пластин плазматической мембраны леммоцита. Наружный, более тонкий и светлый слой оболочки миелинового волокна, представлен цитоплазмой леммоцита с его ядром. Этот слой называют неврилеммой или шванновской оболочкой. По ходу миелинового слоя имеются косо идущие светлые насечки миелина (incisurae myelini). Это места, где между пластинами миелина проникают прослойки цитоплазмы леммоцита. Сужения нервного волокна, где отсутствует миелиновый слой, называют узловыми перехватами (nodi neurofibrae). Они соответствуют границе двух смежных леммоцитов.
Безмиелиновые нервные волокна (neurofibrae nonmyelinatae) более тонкие, чем миелиновые. В их оболочке, образованной тоже леммоцитами отсутствует миелиновый слой, насечки и перехваты. Такое строение безмиелиновых нервных волокон обусловлено тем, что хотя леммоциты и охватывают осевой цилнидр, но они не закручиваются вокруг него. В один леммоцит при этом может быть погружено несколько осевых цилиндров. Это волокна кабельного типа, Безмиелиновые нервные волокна входят преимущественно в состав вегетативной нервной системы. Нервные импульсы в них распространяются медленнее (1 — 2 м/сек), чем в миелиновых, и имеют тенденцию к рассеиванию и затуханию.
Нервные окончания
Нервные волокна заканчиваются концевыми нервными аппаратами, называемыми нервными окончаниями (terminationes neruorum). Различают три вида нервных окончаний: эффекторы (эффекторные), рецепторы (чувствительные) и межнейронные связи — синапсы.
Эффекторы (effectores) бывают двигательными и секреторными. Двигательные окончания представляют собой концевые аппараты аксонов моторных клеток (преимущественно передних рогов спинного мозга) соматической или вегетативной нервной системы. Двигательные окончания в поперечно-полосатой мышечной ткани называют нервно-мышечными окончаниями (синапсами) или моторными бляшками. Моторные нервные окончания в гладкой мышечной ткани имеют вид пуговчатых утолщений или четкообразных расширений. Секреторные окончания выявлены на железистых клетках.
Рецепторы (receptores) представляют собой концевые аппараты дендритов чувствительных нейронов. Одни из них воспринимают раздражение из внешней среды—это экстерорецепторы. Другие получают сигналы от внутренних органов — это интерорецепторы, Среди чувствительных нервных окончаний по их функциональным проявлениям различают: механорецепторы, барорецепторы, терморецепторы и хеморецепторы.
По строению рецепторы подразделяют на свободные — это рецепторы в виде усиков, кустиков, клубочков. Они состоят только из ветвлений самого осевого цилиндра и не сопровождаются нейроглией. Другой вид рецепторов— это несвободные. Они представлены терминалами осевого цилиндра, сопровождаемыми нейроглиальными клетками. Среди несвободных нервных окончаний выделяют инкапсулированные, покрытые соединительнотканными капсулами. Это осязательные тельца Мейснера, пластинчатые тельца Фатер-Пачини и др. Второй разновидностью несвободных нервных окончаний являются неинкапсулированные нервные окончания. К ним относят осязательные мениски или осязательные диски Меркеля, залегающие в эпителии кожи и др.
Межнейрональные синапсы (synapses Interneuronales)— это места контактов двух нейронов. По локализации различают следующие виды синапсов: аксодендритические, аксосоматические и аксоаксональные (тормозные). В световом микроскопе синапсы имеют вид колечек, пуговок, булав (концевые синапсы) или тонких нитей, стелющихся по телу или отросткам другого нейрона. Это так называемые касательные синапсы. На дендритах выявляются синапсы, получившие название дендритических шипиков (шипиковый аппарат). Под электронным микроскопом в синапсах различают так называемый пресинаптический полюс с пресинаптической мембраной одного нейрона и постсинаптический полюс с постсинаптической мембраной (другого нейрона). Между этими двумя полюсами располагается синаптическая щель. На полюсах синапса часто сосредоточено большое количество митохондрий, а в области пресинаптического полюса и синаптической щели — синаптических пузырьков (в химических синапсах).
По способу передачи нервного импульса синапсы различают химические, электрические. В химических синапсах в синаптических пузырьках содержатся медиаторы — норадреналин в адренэргических синапсах (темные синапсы) и ацетилхолин в холинэргических синапсах (светлые синапсы). Нервный импульс в химических синапсах передается с помощью этих медиаторов. В электрических (беспузырьковых) синапсах не имеется синаптических пузырьков с медиаторами. Однако в них наблюдается тесный контакт прс- и постсинаптических мембран. В этом случае нервный импульс передается с помощью электрических потенциалов.
По производимому эффекту различают возбуждающие и тормозные синапсы. В тормозных синапсах медиатором может быть гаммааминомаслянная кислота. В химических синапсах всегда имеет место только одностороннее проведение нервного импульса.
Нейроны посредством синапсов объединяются в нейронные цепи. Цепь нейронов. обеспечивающая проведецие нервного импульса от рецептора чувствительного нейрона до двигательного нервного окончания, называется рефлекторной дугой. Существуют простые и сложные рефлекторные дуги.
Самая простая рефлекторная дуга образована всего двумя нейронами: первый — чувствительный и второй — двигательный. В большинстве случаев в рефлекторных дугах между этими нейронами включены еще ассоциативные, вставочные нейроны. Различают также соматические и вегетативные рефлекторные дуги. Соматические рефлекторные дуги регулируют работу скелетной мускулатуры, а вегетативные — обеспечивают непроизвольное сокращение мускулатуры внутренних органов.
Гистологический препарат. Пластинчатое тельце (тельце Фатер-Пачини).
Окраска гематоксилином и эозином. Малое увеличение. Найти:
- наружную капсулу (наружную луковицу) тельца,
- внутреннюю колбу (внутреннюю луковицу),
- терминали осевого цилиндра,
- ядра глиальных клеток.
Гистологический препарат. Осязательное тельце (тельце Мейснера).
Импрегнация серебром. Большое увеличение. Наити:
- капсулу тельца,
- терминальные ветвления осевого цилиндра,
- осязательные клетки.
Гистологический препарат. Двигательное нервное окончание в поперечнополосатой мышечной ткани. Моторная бляшка.
Импрегнация серебром. Большое увеличение. Найти:
- нервное волокно,
- его концевые веточки,
- ядра глиальных клеток,
- поперечнополосатое мышечное волокно,
- сарколемму,
- моторную бляшку.
Методичка МГМСУ в формате PDF — скачать и читать со страницы 56 (Тема 5. Нервная ткань. Читать весь раздел.)
Методичка МГМСУ. Общая гистология.
Читайте также:
