
Клетки догеля симпатические ганглии
Скопление нейронов вне ЦНС – разделяются на чувствительные (сенсорные) и автономные (вегетативные)
Чувствительные ганглии подразделяются на спинальные и ганглии черепномозговых нервов (V, VII, VIII, IX, X). Первые лежат по ходу задних корешков спинного мозга, вторые по ходу черепномозговых нервов.
Источником развития являются клетки ганглиозных пластинок, которые дифференцируются на нейробласты и глиобласты, дающие основные клетки ганглиев. Сначала они биполярные, затем проксимальные участки значительно растут и сливаются с образованием псевдоуниполярной клетки.
Ганглии снаружи покрыты соединительнотканной капсулой, от которой проходят тонкие прослойки РВСТ, вместе с которыми проникают и кровеносные сосуды. Нейроны лежат по периферии узла в виде групповых скоплений, нервные волокна проходят через центральную часть узла. В ганглии выделяют две разновидности нейронов: темные, мелкие нейроны являются сенсорными клетками в вегетативных рефлекторных дугах, тогда как светлые, крупные в соматических. Периферический отросток является дендритом, идет на периферию и заканчивается чувствительным нервным окончанием или рецептором. Центральные отростки – аксоны, они входят в спинной мозг через задние корешки, затем идут в краевую зону Лиссауэра и там делятся на две ветви: короткую – нисходящую и длинную – восходящую. От этих ветвей отходят более тонкие веточки, которые на разных уровнях спинного мозга связываются с ассоциативными нейронами желатинозной субстанции. Некоторые нейроны непосредственно контактируют с мотонейронами передних рогов. Снаружи нейроны покрыты мантийными олигодендроглиоцитами или клетками сателлитами, а снаружи от них соединительнотканная капсула.
Нейроны чувствительных ганглиев передают нервные импульсы при помощи нервных медиаторов ацетилхолина, глутамата, вещества Р, срматостатина, холицистокинина; также обнаруженгастринивазоинтестинальный полипептид. При помощи вещества Р передается болевая чувствительность (pain – боль) с аксона чувствительного нейрона на нейрон спиноталамического пути. При этом боль блокирует другой нейропептид – энкефалин, который продуцируется вставочными нейронами.
ФУНКЦИЯ –рецепторная, в них не происходит переключения нервного импульса с одного нейрона на другой, узлы не являются нервными центрами.
I порядка - паравертебральные – парные образования идущие по обе стороны позвоночного столба и соединяются между собой межузловыми ветвями,
II порядка – превертебральные. Эти сплетения находятся на аорте и ее ветвях, в области шеи, в грудной, брюшной и тазовой полостях.
Ганглии I и II порядка являются симпатическими
III порядка – парасимпатические и как правило находятся в интрамуральных нервных сплетениях или параорганно
Строение симпатических ганглиев: снаружи покрыты капсулой, от которой проходят прослойки РВСТ, они состоят из мультиполярных нейронов, разных по величине, дендриты которых сильно ветвятся. Аксоны образуют постганглионарные нервные безмиелиновые волокна. Среди нейронов очень часто встречаются многоядерные и полиплоидные клетки. Каждый нейрон ганглия и его отростки окружены глиальной оболочкой, образованной мантийной олигодендроглией, снаружи от глиальной оболочки к ней примыкает соединительнотканная оболочка. Кроме постганглионарных нейронов в ганглии имеются малые, являясь тормозными ассациативными тормозными нейронами. Они блокируют передачу возбуждения с преганглионарного волокна на пресинаптические нейроны.
Парасимпатические и метасимпатические нервные ганглии к ним относятся ганглии III порядка, они находятся в интрамуральных нервных сплетениях или параорганно. В стенках полых органов они предсталяют мейснеровское сплетение (подслизистое) или ауэрбаховское сплетение (межмышечное). Основную массу нейроцитов ганглиев III порядка составляют клетки Догеля трех типов.
Клетки Догеля I типа – двигательные. Их волокна образуют постганглионарные безмиелиновые волокна, идущие к иннервируемымы структурам. Они имеют длинный аксон, поэтому наз. длинноаксональными.
Клетки Догеля II типа, равноотростчатые, по ф-ции это чувствительные нейроны, их дендриты расположены на иннервируемом органе, аксон на дендрите или теле Догеля I тем сасмым формируя местные рефлекторные дуги.
Догеля III ассоциативный нейрон. Их дендриты образуют связи несколькими клетками I и II типа, а аксоны идут в соседние ганглии, осуществляя межганглионарные связи.
Кроме этих трех основных нейронов в парасимпатических ганглиях имеются пуринергические нейроциты, а таже нейроны содержащие ВИП, соматостатин, и другие нейрогормоны, осуществляющие нейрогуморальную регуляцию органов.
Нейроны окружены майтийной олиглдендроглией, базальной мембраной и капсулой из РВСТ.
Центральная нервная система человека осуществляет контроль над деятельностью его организма и разделяется на несколько отделов. Головной мозг посылает и получает сигналы из организма и после их обработки имеет информацию о процессах. Нервная система разделяется на вегетативную и соматическую нервную системы.
Отличия вегетативной и соматической нервной системы
Соматическая нервная система регулируется сознанием человека и может управлять деятельностью скелетной мускулатуры. Все компоненты реакции человека на внешние факторы находятся под контролем полушарий мозга. Она обеспечивает сенсорные и моторные реакции человека, контролируя их возбуждение и торможение.
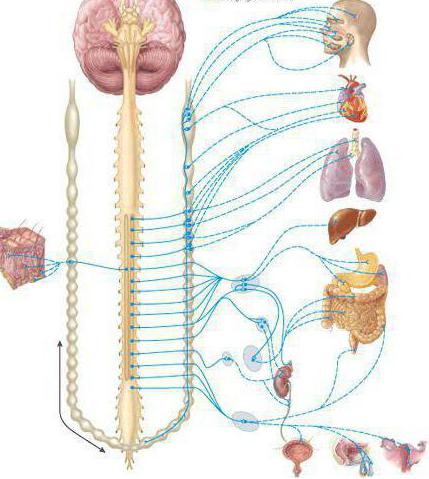
Вегетативная нервная система контролирует периферическую деятельность организма и не контролируется сознанием. Для нее характерны автономность и генерализованность воздействия на организм при полном отсутствии сознания. Эфферентная иннервация внутренних органов позволяет ей контролировать обменные процессы в организме и осуществлять обеспечение трофических процессов скелетной мускулатуры, рецепторов, кожи и внутренних органов.
Строение вегетативной системы
Работа вегетативной нервной системы контролируется гипоталамусом, который находится в центральной нервной системе. Вегетативная нервная система имеет метасегментарное строение. Ее центры находятся в головном, спинном мозге и коре головного мозга. Периферические отделы образованы стволами, ганглиями, сплетениями.
В вегетативной нервной системе различают:
- Симпатическую. Ее центр расположен в грудопоясничном отделе спинного мозга. Для нее характерны паравертебральные и предвертебральные ганглии ВНС.
- Парасимпатическую. Ее центры сосредоточены в среднем и продолговатом мозге, крестцовом отделе спинного мозга. Нервные узлы в основном интрамуральные.
- Метасимпатическую. Иннервирует желудочно-кишечный тракт, сосуды, внутренние органы организма.
В состав ее входит:
- Ядра нервных центров, расположенных в головном и спинном мозге.
- Вегетативные ганглии, которые расположены по периферии.
- Нервные волокна.
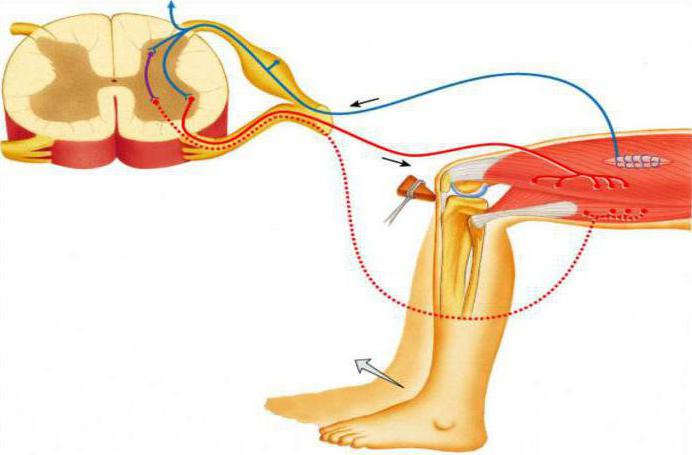
Рефлекторная дуга автономной нервной системы
Рефлекторная дуга вегетативной нервной системы состоит из трех звеньев:
- чувствительное или афферентное;
- вставочное или ассоциативное;
- эффекторное.

Чувствительное звено
Чувствительное звено расположено в спинномозговом ганглии. Этот ганглий имеет нервные клетки, сформированные группами, и их контроль осуществляется ядрами центрального головного мозга, большими полушариями и их структурами.
Чувствительное звено представлено частично униполярными клетками, которые имеют один приносящий или уносящий аксон, и они принадлежат спинальным или черепно-мозговым узлам. А также узлами блуждающих нервов, имеющих строение, похожее на спинальные клетки. В это звено входят клетки Догеля II типа, которые являются компонентами вегетативных ганглиев.
Вставочное звено
Вставочное звено в вегетативной нервной системе служит для передачи через низшие нервные центры, которыми являются вегетативные ганглии, и осуществляется это через синапсы. Расположено оно в боковых рогах спинного мозга. Нет прямой связи от афферентного звена на преганглионарные нейроны для их связи, существует кратчайший путь от афферентного нейрона до ассоциативного и от него до преганглионарного нейрона. Передача сигналов и нервных импульсов от афферентных нейронов в различных центрах осуществляется с различным количеством вставочных нейронов.
Например, в дуге спинального автономного рефлекса между чувствительным и эффекторным звеном существует три синапса, два из которых расположены в спинном мозге, а один в вегетативном узле, в котором расположен эфферентный нейрон.
Эфферентное звено
Эфферентное звено представлено эффекторными нейронами, которые расположены в вегетативных узлах. Их аксоны образуют безмиелиновые волокна, которые в составе со смешанными нервными волокнами иннервируют внутренние органы.
Вегетативные рефлекторные дуги расположены в боковых рогах.
Строение нервного узла
Ганглий – это скопление нервных клеток, которые имеют вид узелковых расширений толщиной около 10 мм. По своему строению вегетативный ганглий сверху покрыт соединительнотканной капсулой, которая образует строму из рыхлой соединительной ткани внутри органов. Мультиполярные нейроны, которые строятся из округлого ядра и крупных ядрышек, состоят из одного эфферентного нейрона и нескольких расходящихся афферентных нейронов. Эти клетки относятся по своему типу к клеткам головного мозга и являются двигательными. Их окружает неплотная оболочка – мантийная глия, которая создает постоянную среду для нервной ткани и обеспечивает полноценное функционирование нервных клеток.
Вегетативный ганглий имеет диффузное расположение нервных клеток и множество отростков, дендритов и аксонов.
Спинномозговой ганглий имеет нервные клетки, которые расположены группами, и их расположение имеет порядок обусловленный.
Вегетативные нервные ганглии разделяются на:
- Сенсорные нейроны, которые расположены близко к спинному или центральному отделу мозга. Униполярные нейроны, из которых состоит этот ганглий, представляют собой приносящий или уносящий отросток. Они служат для афферентной передачи импульсов, и их нейроны образуют бифуркацию при ветвлении отростков. Эти отростки передают информацию от периферии к центральному афферентному нейрону – это периферический отросток, центральный – от тела нейрона в мозговой центр.
- Моторные, двигательные состоят из эфферентных нейронов, и в зависимости от их положения их называют паравертебральные, предвертебральные.
Симпатические ганглии
Паравертебральные цепочки ганглиев расположены вдоль позвоночного столба в симпатических стволах, которые идут длинной вереницей от основания черепа до копчика.
Предвертебральные нервные сплетения находятся ближе к внутренним органам, и их локализация сосредоточена перед аортой. Они образуют брюшное сплетение, которое состоит из солнечного, нижне- и верхнебрыжеечного сплетений. Они представлены двигательными адренергическими и тормозящими действие холинергическими нейронами. Также связь между нейронами осуществляется преганглионарными и постганглионарными нейронами, которые используют медиаторы ацетилхолин и норадреналин.
Интрамуральные нервные узлы имеют нейроны трех типов. Их описание было сделано русским ученым Догелем А.С., который, исследуя гистологию нейронов вегетативной нервной системы, выделил такие нейроны, как длинноаксонные эфферентные клетки первого типа, равноотросчатые афферентные клетки второго типа и ассоциативные клетки третьего типа.
Рецепторы ганглиев
Афферентные нейроны выполняют узкоспециализированую функцию, и их роль заключается в восприятии раздражителей. Такими рецепторами являются механорецепторы (реакция на растяжение или давление), фоторецепторы, терморецепторы, хеморецепторы (отвечают за реакции в организме, химические связи), ноцицепторы (реакция организма на болевые раздражители – это повреждение кожи и другие).
В симпатических стволах эти рецепторы по рефлекторной дуге передают в центральную нервную систему информацию, которая служит сигналом о повреждениях или нарушениях в организме, а также нормальной его работе.
Функции ганглиев
Каждый ганглий имеет свое местоположение, кровоснабжение, и его функции определяются этими параметрами. Спинномозговой ганглий, имеющий иннервацию из ядер головного мозга, обеспечивает непосредственную связь процессов в организме через рефлекторную дугу. От этих структурных компонентов спинного мозга иннервируются железы, гладкая мускулатура мышц внутренних органов. Сигналы, поступающие по рефлекторной дуге, идут медленнее, чем в ЦНС, и они полностью регулируются вегетативной системой, также она обладает трофической, сосудодвигательной функцией.
Вегетативные ганглии представляют собой скопление многочислен-ных мультиполярных нервных клеток.
Величина вегетативных ганглиев существенно варьирует. В связи с этим различают крупные, средней величины, мелкие и очень мелкие (микроганглии) ганглии.
Необходимо отметить, что кроме анатомически обособленных ганглиев, по ходу вегетативных ветвей периферических нервов встречается большое количество нервных клеток, подобных нервным клеткам вегетативного ганглия. Эти нейроны, мигрирующие сюда в ходе эмбриогенеза, локализуются по ходу нервов поодиночке или образуют небольшие группы – микроганглии.
Вегетативный ганглий с поверхности покрыт фиброзной соединительнотканной капсулой, от которой внутрь отходят многочисленные прослойки соединительной ткани, образующей строму узла. По этим прослойкам в узел проходят кровеносные сосуды, питающие его и образующие в нем капиллярную сеть. В капсуле и строме узла часто вблизи кровеносных сосудов встречаются рецепторы – диффузные, кустиковидные или инкапсулированные.
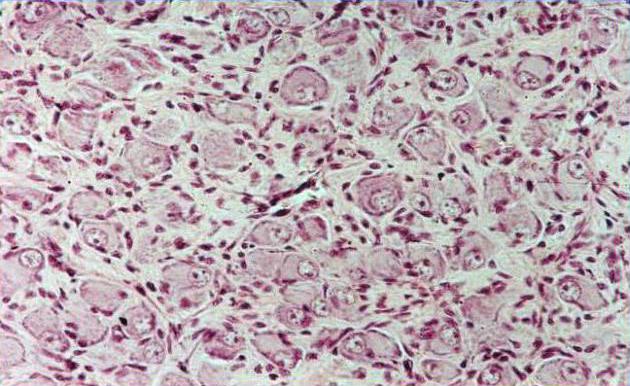
Мультиполярные нервные клетки вегетативного ганглия впервые были описаны А.С. Догелем. При этом Догель выделил 3 типа нервныхклеток вегетативного ганглия, которые получили названияклеток Догеля I, II, III типа. Морфофункциональные характеристики клеток Догеля существенно разнятся.
Клетки Догеля I типапо функциональному значению являются эффекторными (двигательными) нейронами. Это более или менее крупные нервные клетки, с несколько короткими дендритами, не выходящими за пределы данного ганглия. Аксон этих клеток более длинный выходит за пределы ганглия и направляется к рабочему аппарату – гладкомышечным клеткам, железистым клеткам, образуя на них двигательные (или соответственно секреторные) нервные окончания. Аксоны и дендриты клеток ДогеляIтипа являются безмякотными. Дендриты часто образуют пластинчатые расширения, на которых (как и на теле клетки) располагаются синаптические окончания, образующиеся разветвлениями преганглионарного нервного волокна.
Тела нейронов в вегетативном ганглии, в отличие от спиномозгового ганглия, располагаются беспорядочно по всему узлу и более рыхло (т.е. более редко). На препаратах, окрашенных гематоксилином или другими общегистологическими красителями, отростки нервных клеток остаются не выявленными, а клетки имеют такую же округлую безотросчатую форму, как в спиномозговых узлах. Тело каждой нервной клетки (как и в спинальном ганглии) окружено слоем уплощенных элементов олигодендроглии – слоем сателлитов.
К наруже от слоя сателлитов имеется еще тонкая соединительно-тканная капсула. Клетки Догеля Iтипа являются основной клеточной формой вегетативных ганглиев.
Клетки Догеля II типа– это также мультиполярные нервные клетки, с несколькими длинными дендритами и нейритом, уходящим за пределы данного ганглия в соседние ганглии. Аксон с поверхности покрыт миелином. Дендриты этих клеток начинаются рецепторными аппаратами в гладких мышцах. С функциональной точки зрения клетки ДогеляIIтипа являются чувствительными. В отличие от чувствительных псевдоуниполярных нервных клеток спиномозгового узла клетки ДогеляIIтипа, по-видимому, образуют рецепторное (афферентное) звено местных рефлекторных дуг, замыкаемых без захода нервного импульса в центральную нервную систему.
Клетки Догеля III типапредставляют собой местные ассоциативные (вставочные) элементы, соединяющие своими отростками несколько клетокIиIIтипа. Их дендриты короткие, но более длинные, чем у клетокIтипа, не выходят за пределы данного ганглия, а образуют корзинчатые разветвления, оплетающие тела других клеток данного ганглия. Нейрит клетки ДогеляIIIтипа идет в другой ганглий и там вступает в синаптическую связь с клеткамиIтипа. Следовательно, клеткиIIIтипа входят в качестве ассоциативного звена в местные рефлекторные дуги.
Нельзя не отметить, существует такая точка зрения, что клетки Догеля IIIтипа имеют рецепторную или эффекторную природу.
Соотношение численности клеток Iи II типов Догеля в различных вегетитивных ганглиях неодинаково. Парасимпатические ганглии, в отличие от симпатических ганглиев, характеризуются преобладанием клеток с короткими внутрикапсулярными дендритами, отсутствием или малым количеством пигмента в клетках. Кроме того в парасимпатических ганглиях, как правило, тела лежат значительно компактнее, чем в симпатических ганглиях. Кроме того, в симпатических ганглиях имеютсяМИФ-клетки(мелкие клетки с интенсивной флюоросценцией).
Через вегетативный ганглий проходят проводящие пути трех видов: центростремительные, центробежные и периферические (местные) рефлекторные.
Центростремительные пути образованы чувствительными отростками псевдоуниполярных клеток спинального ганглия, начинающихся рецепторами в иннервируемых тканях, а также внутри ганглия. Эти волокна проходят транзитом через вегетативные ганглии.
Центробежные пути представлены преганглионарными волокнами, которые многократно ветвятся в вегетативном узле и образуют синапсы на многих клеточных телах эффекторных нейронов. Например, в верхнем шейном узле соотношение числа преганглионарных волокон, вступивших в него, к постганглионарным равно 1:32. Это явление приводит, при возбуждении преганглионарных волокон, к резкому расширению области возбуждения (геперализация эффектора). Благодаря этому, сравнительно небольшое количество центральных вегетативных нейронов обеспечивает нервными импульсами все органы и ткани. Итак, например, при раздражении у животного преганглионарных симпатических волокон, проходящих через передние корешки IYгрудного сегмента, может наблюдаться сужение сосудов кожи головы, шеи, расширение коронарных сосудов, сужение сосудов кожных покровов передней конечности, сосудов почки и селезенки.
Продолжение этих путей составляют постганглионарные волокна, достигающие иннервируемых тканей.
Периферические (местные) рефлекторные пути начинаются в тканях разветвлениями отростков собственных чувствительных нейронов вегетативных ганглиев (т.е. клетками IIтипа Догеля). Нейриты же этих кле-ток заканчиваются на клетках ДогеляIтипа, чьи постганглионарные волокна входят в состав центробежных путей.
Морфологическим субстратом рефлекторной деятельности вегетативной нервной системы является рефлекторная дуга. Для рефлектор-ной дуги вегетативной нервной системы характерны все три звена – рецепторное (афферентное), вегетативное (ассоциативное) и эффекторное (двигательное), но локализация их иная чем в соматической.
Интересно отметить, что многие морфологи и физиологи указывают как на отличительный признак вегетативной нервной системы, отсутствие в ее составе собственного афферентного (рецепторного) звена, т.е. они считают, что чувствительная иннервация внутренних органов, сосудов и т.д. осуществляется дендритами псевдоуниполярных клеток спинального ганглия, т.е. соматической нервной системы.
Правильнее считать, что спиномозговые узлы содержат нейроны, иннервирующие скелетную мускулатуру, кожу (т.е. нейроны соматической нервной системы), так и нейроны, иннервирующие все внутренние органы, сосуды (т.е. вегетативные нейроны).
Одним словом, аффекторное звено, как и в соматической (анимальной) нервной системе, в вегетативной нервной системе представлено клеткой, лежащей в спиномозговом узле.
Тело нейрона ассоциативного звена располагается, в отличие от соматической рефлекторной нервной дуги, не в области заднего рога, а в боковых рогах серого вещества, и аксон этих клеток выходит за пределы мозга и оканчивается в одном из вегетативных ганглиев.
Наконец, наибольшие отличия между анимальной и вегетативной рефлекторными дугами наблюдаются в эфферентном звене. Так, тело эфферентного нейрона в соматической нервной системе находится в сером веществе спинального или головного ганглия и лишь его аксон идет на периферию в составе того или иного черепно или спиномозгового нерва. В вегетативной системе тела эффекторных нейронов находятся на периферии: они либо разбросаны по ходу некоторых нервов, либо образуют скопления – вегетативные ганглии.
Таким образом, для вегетативной нервной системы, в силу такой локализации эффекторных нейронов, характерно наличие, по крайней мере, одного перерыва эфферентного пути, который проходит в вегетативном ганглии, т.е. здесь нейриты вставочных нейронов, контактируют с нейронами эффекторными, образуя на их телах и дендриты синапсы. Следовательно, вегетативные ганглии представляют собой периферические нервные центры. Этим они принципиально отличаются от спинальных ганглиев, которые не являются нервными центрами, т.к. в них нет синапсов и не происходит переключения нервных импульсов.
Таким образом, спиномозговые узлы являются смешанными образованиями, анимально-вегетативными.
Особенностью рефлекторной дуги симпатической нервной системы является наличие коротких преганглионарных волокон и очень длинных постганглионарных волокон.
Особенностью рефлекторной дуги парасимпатической нервной системы является, напротив, наличие очень длинных преганглионарных и очень коротких постганглионарных волокон.
Основные функциональные различия симпатической и парасимпатической систем заключаются в следующем. Медиатором, т.е. веществом, образующимся в области синапсов и осуществляющим химическую передачу импульса, в симпатических нервных окончаниях является симпатин (вещество, тождественное гормону мозгового вещества надпочечника – ноадреналину).
Названные химические вещества – медиаторы и сами по себе, даже без раздражения вегетативных нервных волокон, вызывают в рабочих органах эффекты, аналогичные действию соответствующих вегетативных нервных волокон. Так, ноадреналин при введении в кровь ускоряет сердцебиение, но замедляет перистальтику кишечного тракта, а ацетилхолин – наоборот. Ноадреналин вызывает сужение, а ацетилхолин – расширение просвета сосудов.
Холинергичны и синапсы, образуемые волокнами соматической нервной системы.
Деятельность вегетативной нервной системы находится под контролем коры больших полушарий, а также подкорковых вегетативных центров полосатого тела и, наконец, вегетативных центров промежуточного мозга (ядро гипоталамуса).
В заключении необходимо отметить, что учение о вегетативной нервной системе большой вклад внесли и советские ученые Б.И. Лаврентьев, А.А. Заварзин, Д.И. Голуб, удостоенные государственных премий.
Жаботинский Ю.М. Нормальная и патологическая морфология вегетативных ганглиев. М.,1953
Заварзин А.А. Очерк по эволюционной гистологии нервной системы. М-Л,1941
А.Г. Кнорре, И.Д.Лев. Вегетативная нервная система. Л.,1977,с.120
Колосов Н.Г. Иннервация пищеварительного тракта человека. М-Л,1962
Колосов Н.Г. Вегетативный узел. Л.,1972
Колосов Н.Г., Хабарова А.Л. Структурная организация вегетативных ганглиев. Л.,Наука, 1978.-72с.
Кочетков А.Г., Кузнецов Б.Г., Коновалова Н.В. Вегетативная нервная система. Н-Новгород, 1993.-92с.
Мельман Е.П. Функциональная морфология иннервация органов пищеварения. М.,1970
Ярыгин Н.Е. и Ярыгин В.Н. Патологические и приспособительные изменения нейрона. М.,1973.
В вегетативной нервной системе различают центральные и периферические отделы. Центральные отделы симпатической нервной системы представлены ядрами боковых рогов тораколюмбального отдела спинного мозга. В парасимпатической нервной системе центральные отделы включают ядра среднего и продолговатого мозга, а также ядра боковых рогов сакрального отдела спинного мозга. Парасимпатические волокна краниобульбарного отдела выходят в составе III-й, VII-й, IX-й и Х-й пар черепных нервов.
Периферические отделы вегетативной нервной системы образованы нервными стволами, ганглиями и сплетениями.
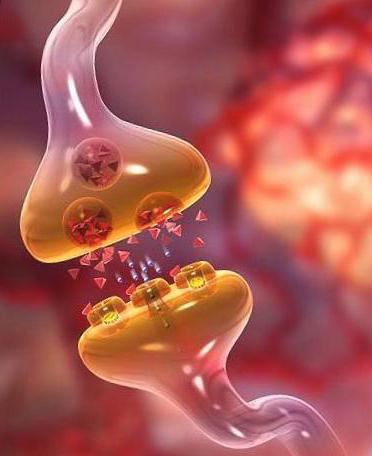
Вегетативные рефлекторные дуги начинаются чувствительным нейроном, тело которого лежит в спинномозговом узле (ганглии), как и в соматической рефлекторной дуг. Ассоциативные нейроны находятся в боковых рогах спинного мозга. Здесь нервные импульсы переключаются на промежуточные преганглионарные нейроны, отростки которых покидают центральные ядра и достигают вегетативных ганглиев, где передают импульсы на двигательный нейрон. В связи с этим различают нервные волокна преганглионарные и постганглионарные. Первые из них покидают центральную нервную систему в составе вентральных корешков спинномозговых нервов и черепных нервов. Как в симпатической, так и в парасимпатической системах преганглионарные нервные волокна принадлежат холинергическим нейронам. Аксоны нейронов, расположенных в вегетативных ганглиях, называются постганглионарными. Они не образуют прямых контактов с эффекторными клетками. Их терминальные отделы по своему ходу формируют расширения — варикозности, в составе которых находяся пузырьки медиатора. В области варикозности нет глиальной оболочки и нейромедиатор, выделяясь в окружающую среду, влияет на эффекторные клетки (например, на клетки желез, гладкие миоциты и др.).
В периферических ганглиях симпатической нервной системы, как правило, находятся адренергические эфферентные нейроны (за исключением нейронов, имеющих синаптические связи с потовыми железами, где симпатические нейроны являются холинергическими). В парасимпатических ганглиях эфферентные нейроны всегда холинергические.
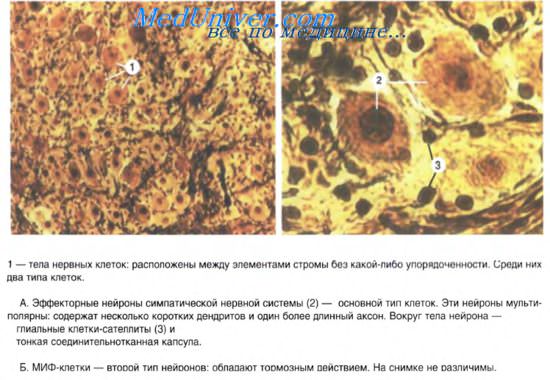
Ганглии представляют собой скопления мультиполярных нейронов (от нескольких клеток до десятков тысяч). Экстраорганные (симпатические) ганглии имеют хорошо выраженную соединительнотканную капсулу, как продолжение периневрия. Парасимпатические ганглии находятся, как правило, в интрамуральных нервных сплетениях. Ганглии интрамуральных сплетений, как и другие вегетативные узлы, содержат вегетативные нейроны местных рефлекторных дуг. Мультиполярные нейроны диаметром 20-35 мкм расположены диффузно, каждый нейрон окружен глиоцитами ганглия. Кроме того, описаны нейроэндокринные, хеморецепторные, биполярные, а у некоторых позвоночных и униполярные нейроны. В симпатических ганглиях имеются мелкие интенсивно флюоресцирующие клетки (МИФ-клетки) с короткими отростками и большим количеством гранулярных пузырьков в цитоплазме. Они выделяют катехоламины и оказывают тормозящее влияние на передачу импульсов с преганглионарных нервных волокон на эфферентный симпатический нейрон. Эти клетки называют интернейронами.
Среди крупных мультиполярных нейронов вегетативных ганглиев различают: двигательные (клетки Догеля I-го типа), чувствительные (клетки Догеля II-го типа) и ассоциативные (клетки Догеля III-го типа). Двигательные нейроны имеют короткие дендриты с пластинчатыми расширениями ("рецептивные площадки"). Аксон этих клеток очень длинный, уходит за пределы ганглия в составе постганглионарных тонких безмиелиновых нервных волокон и оканчивается на гладких миоцитах внутренних органов. Клетки I-го типа называют длинноаксонными нейронами. Нейроны II-го типа — равноотростчатые нервные клетки. От их тела отходят 2-4 отростка, среди которых различить аксон трудно. Не разветвляясь, отростки уходят далеко от тела нейрона. Их дендриты имеют чувствительные нервные окончания, а аксон оканчивается на телах двигательных нейронов в соседних ганглиях. Клетки II-го типа являются чувствительными нейронами местных вегетативных рефлекторных дуг. Клетки Догеля III-го типа по форме тела похожи на вегетативные нейроны П-го типа, но их дендриты не выходят за пределы ганглия, а нейрит направляется в другие ганглии. Многие исследователи считают эти клетки разновидностями чувствительных нейронов.
Таким образом, в периферических вегетативных ганглиях имеются местные рефлекторные дуги, состоящие из чувствительных, двигательных и, возможно, ассоциативных вегетативных нейронов.
Интрамуральные вегетативные ганглии в стенке пищеварительного тракта отличаются тем, что в их составе, кроме двигательных холинергических нейронов, имеются тормозные нейроны. Они представлены адренергическими и пуринергическими нервными клетками. В последних медиатором является пуриновый нуклеотид. В интрамуральных вегетативных ганглиях встречаются также пептидергические нейроны, выделяющие вазоинтестинальный пептид, соматостатин и ряд других пептидов, с помощью которых осуществляются нейроэндокринная регуляция и модуляция деятельности тканей и органов пищеварительной системы.
Ганглии нервной системы — скопление нейронов и глии, находящиеся за пределами головного и спинного мозга.
Подобные образования в центральной нервной системе называют ядрами. Они выступают связующими звеньями структур нервной системы, осуществляют первичную обработку импульсов, отвечают за функции висцеральных органов.
Человеческий организм осуществляет два типа функций – соматические и вегетативные. Соматические подразумевают восприятие внешних раздражителей и реакцию на них при помощи скелетной мускулатуры. Эти реакции могут контролироваться сознанием человека, а за их выполнение отвечает центральная нервная система.
Вегетативные функции – пищеварение, метаболизм, кроветворение, кровообращение, дыхание, потоотделение и другое, контролирует вегетативная система, которая не зависит от человеческого сознания. Кроме регуляции работы висцеральных органов, вегетативная система обеспечивает трофику мускулатуры и ЦНС.
Ганглии, отвечающие за соматические функции, представляют спинномозговые узлы и узлы черепных нервов. Вегетативные, в зависимости от расположения центров в ЦНС, делят на: парасимпатические и симпатические.
Первые находятся в стенках органа, а симпатические дистанционно – в структуре, называемой пограничным стволом.
Строение ганглия
В зависимости от морфологических особенностей, размер ганглиев колеблется от нескольких микрометров до нескольких сантиметров. По сути это скопление нервных и глиальных клеток, покрытое соединительной оболочкой.
Соединительнотканный элемент пронизан лимфатическими и кровеносными сосудами. Каждый нейроцит (или группа нейроцитов) окружен капсульной оболочкой, выстланной изнутри эндотелием, а снаружи волокнами соединительной ткани. Внутри капсулы располагается нервная клетка и глиальные структуры, которые обеспечивают жизнедеятельность нейрона.
От нейрона отходит один аксон, покрытый миелиновой оболочкой, который разветвляется на две части. Одна из них входит в состав периферического нерва и образует рецептор, а вторая направляется в ЦНС.
Вегетативные центры находятся в стволе головного мозга и спинном мозге. Парасимпатические центры локализуются в краниальном и сакральном отделах, а симпатические – в тораколюмбальном.
Ганглии вегетативной нервной системы
Симпатическая система включает два типа узлов: вертебральные и превертебральные.
Вертебральные располагаются с двух сторон от позвоночного столба, образуя пограничные стволы. Они связаны со спинным мозгом с помощью нервных волокон, которые дают начало белым и серым соединительным веткам. Выходящие из узла нервные волокна направляются к висцеральным органам.
Превертебральные расположены на большем расстоянии от позвоночника, при этом также дистанционно они находятся от органов, за которые они ответственны. Примером превертебральных узлов служат шейные, брыжеечные скопления нейронов, солнечное сплетение.
Парасимпаческий отдел образован ганглиями, расположенными на органах, или в непосредственной к ним близости.
Внутриорганные нервные сплетения размещены на органе или в его стенке. Крупные внутриорганные сплетения расположены в сердечной мышце, в мышечном слое стенки кишечника, в паренхиме железистых органов.
Ганглии вегетативной и центральной нервной системы имеют свойства:
- проведение сигнала в одну сторону;
- входящие в узел волокна перекрывают зоны влияния друг друга;
- пространственная суммация (сумма импульсов способна породить потенциал в нейроците);
- окклюзия (стимуляция нервов вызывает малый ответ, чем стимуляция каждого в отдельности).
Синоптическая задержка в вегетативных ганглиях больше, чем в аналогичных структурах ЦНС, а постсинаптический потенциал продолжительный. Волну возбуждения в ганглионарных нейроцитах сменяет депрессия. Эти факторы приводят к относительно низкому ритму импульсов, сравнительно с ЦНС.
Какие функции выполняют ганглии
Основным назначением вегетативных узлов является распределение и передача нервных импульсов, а также генерация локальных рефлексов. Каждый ганглий, зависимости от расположения и особенностей трофики, отвечает за функции определенного участка тела.
Ганглиям присуща автономность от ЦНС, что позволяет им регулировать деятельность органов без участия головного и спинного мозга.
В структуре внутристеночных узлов содержатся клетки – водители ритма, способные задавать периодичность сокращений гладкой мускулатуры кишечника.
Особенность связана с прерыванием, направляющимся к внутренним органам, волокон ЦНС на периферических узлах вегетативной системы, где они образуют синапсы. При этом непосредственное влияние на внутренний орган оказывают аксоны, выходящие из ганглия.
Каждое нервное волокно, приходящее в симпатический ганглий, осуществляет иннервацию до тридцати постганглионарных нейроцитов. Это дает возможность умножать сигнал и распространять импульс возбуждения, выходящий из нервного узла.
В парасимпатические узлах одно волокно осуществляет иннервацию не более четырех нейроцитов, а передача импульса осуществляется локально.
Ганглии – рефлекторные центры
Ганглии нервной системы принимают участие в рефлекторной дуге, что позволяет корректировать деятельность органов и тканей без участия мозга. В конце девятнадцатого века русский гистолог Догель в результате опытов по изучению нервных сплетений в желудочно-кишечном тракте, выявил три типа нейронов – моторные, вставочные и рецепторные, а также синапсы между ними.
Наличие рецепторных нервных клеток подтверждает и возможность трансплантации сердечной мышцы от донора реципиенту. Если бы регуляция сердечного ритма осуществлялась через ЦНС, после пересадки сердца нервные клетки подверглись дегенерации. Нейроны и синапсы в пересаженном органе продолжают функционировать, что свидетельствует об их автономности.
В конце двадцатого века экспериментальным путем установлены механизмы периферических рефлексов, которые делают превертебральные и интрамуральные вегетативные узлы. Возможность создавать рефлекторную дугу свойственна некоторым узлам.
Местные рефлексы позволяют разгрузить центральную нервную систему, делают регуляцию важных функций более надежность, способны продолжать автономную работу внутренних органов в случае прерывания связи с ЦНС.
Вегетативные узлы получают и обрабатывают информацию о работе органов, после чего направляют ее в мозг. Это вызывает рефлекторную дугу как в вегетативной, так и в соматической системе, что запускает не только рефлексы, но и сознательные поведенческие реакции.
Читайте также:
