
Колебание потенциала в нервных клетках
В конце XVIII века [10] Луиджи Гальвани открыл электрическую природу нервного импульса, он установил, что мышечное сокращение конечностей препарированной лягушки могут вызывать раздражения электрическим током, а сама живая ткань является источником электрического импульса. В начале XIX века утвердилось представление о том, что электрический ток распространяется по нервным волокнам как по проводам. Однако Гельмгольцем во второй половине XIX века было показано, что скорость распространения нервного импульса в 3 млн. раз меньше, чем скорость распространения электрического импульса по проводам. Поэтому к концу XIX века гипотеза электрической природы нервного импульса была отвергнута. Только к середине XX века в работах лаборатории английского физиолога А. Ходжкина было найдено экспериментальное подтверждение того, что нервный импульс представляет собой импульс электрического тока. За это открытие в 1963 году Ходжкин, Хаксли и Иклс получили Нобелевскую премию по медицине.
Опыт по исследованию нервного импульса ученые проводили на гигантском аксоне кальмара методом микроэлектродов, которые вводились в аксон нейрона. Было установлено, что введя внутрь клетки микроэлектрод, соединенный с регистрирующей установкой [11] мгновенно (скачком) возникает некоторый постоянный электроотрицательный потенциал по отношению к электроду, расположенному в окружающей клетку жидкости. Величина разности этих потенциалов у нервных клеток и волокон, например гигантских нервных волокон кальмара, составляет около -70 мВ. Эту величину называют мембранным потенциалом покоя (МПП). Во всех точках аксоплазмы этот потенциал практически одинаков.

Рисунок 1 – Внутриклеточное раздражение и отведение потенциалов гигантского аксона кальмара при введении электродов
Кроме регистрирующего электрода в аксон ввели дополнительный раздражающий электрод, через который в нервное волокно подавались толчки электрического тока. При подаче слабого электрического тока внутриклеточный электрод регистрировал кратковременное падение мембранного потенциала (МП), т.е. возникал так называемый электротонический потенциал (ЭП) При подаче несколько более сильного тока происходило дальнейшее падение мембранного потенциала, возникал так называемый подпороговый локальный ответ (ЛО), который в естественных условиях не распространялся. При усилении тока до критического уровня происходило резкое изменение заряда мембраны с отрицательного на положительный заряд. Это явление получило название явление деполяризации, а быстрое колебание заряда на мембране нервной клетки получило название потенциал действия.
Определение_2
Потенциал действия нервной клетки – это электрический процесс, выражающийся в быстром колебании мембранного потенциала и распространяющийся без затухания (без декремента)
В естественных условиях раздражающим фактором, вызывающим потенциал действия в нервной клетке является изменение проницаемости клеточной мембраны для ионов натрия, при сохранении высокой проницаемости ионов калия. Количество натриевых каналов примерно в 10 раз больше числа калиевых каналов, но в покоящемся состоянии они остаются закрытыми. Изменение состояния мембраны (например, действие медиатора, или электрического тока) вызывает открытие ионных каналов натрия, и эти ионы устремляются внутрь нервной клетки. Положительно заряженные ионы вначале нейтрализуют внутренний отрицательный заряд мембраны, а затем изменяют его на положительный заряд.
Величину потенциала действия определяют ионы натрия
Величину мембранного потенциала действия можно оценить по уравнению Нернста для ионов натрия (2):
φм ПД = RT/FZ · ln ([Na + ]н / [Na + ]в) (2)
где [Na + ]н и [Na + ]в – молярные концентрации ионов по обе стороны мембраны,
R – универсальная газовая постоянная (8,31 Дж/(моль · К)),
Т – температура, градусы Кельвина (T=273+t),
F – постоянная Фарадея (96500 Кл/моль),
Величина ПД нейрона колеблется в пределах 80-110 мВ.
ПД либо совсем не возникает на раздражение клетки, если оно мало, либо он максимальной величины, если раздражение является пороговым или сверхпороговым.
В составе ПД различают три фазы (рассмотрите рисунок 2):
1 фаза - деполяризация, т.е. исчезновение заряда клетки - уменьшение мембранного потенциала до нуля;
2 фаза - инверсия, изменение заряда клетки на противоположный, когда внутренняя сторона мембраны клетки заряжается положительно, а внешняя - отрицательно (от лат. inversio - переворачивание);
3 фаза - реполяризация, восстановление исходного заряда клетки, когда внутренняя поверхность клеточной мембраны снова заряжается отрицательно, а наружная - положительно.

Рисунок 2 – Схема, отражающая процесс возбуждения: 1 - деполяризация, 2 - инверсия, 3 - реполяризация, 4 – следовая гиперполяризация
Вся нисходящая часть пика потенциала действия обусловлена выходом иона калия из клетки, она сопровождается восстановлением потенциала покоя мембраны и поэтому называется фазой реполяризации. Фаза реполяризации длится от 0,5 до 1,5 мс.
Возникновение потенциала действия возможно при достижении необходимого критического порога (КУД – критический уровень деполярицаии). Достигнув некоторого положительного значения, назовем его потенциалом реверсии (φм рев ), мембранный потенциал возвращается к значению потенциала покоя (φм ПП ). Таким образом, амплитуду потенциала действия можно вычислить по выражению (3):
В случае достижения КУД, возникает локальный потенциал, т.е. местное не распространяющееся подпороговое возбуждение, существующее в пределах от потенциала покоя (-70 мВ) до критического уровня деполяризации (-50 мВ).
В основе возбуждения мембраны нервных клеток лежит повышение проницаемости мембраны для ионов натрия и открытие натриевых каналов. Внешнее раздражение вызывает перемещение заряженных частиц внутри клетки, что уменьшает исходную разность потенциалов по обе стороны мембраны. Небольшие величины деполяризации приводят к открытию новых ионных каналов натрия и запускают процесс деполяризации всей мембраны. Изменение проницаемости мембраны в локальном (ограниченном месте) называют подпороговым возбуждением или локальным током.
При увеличении раздражения деполяризация мембраны достигает порога возбудимости или критического уровня деполяризации (КУД). В результате открывается практически все натриевые каналы, и ионы натрия лавинообразно устремляются в клетку. Лавинообразное поступление ионов натрия внутрь клетки, вызывающее резкое смену заряда мембраны, что и регистрируется в виде потенциала действия. Внутренняя сторона мембраны в месте возбуждения оказывается заряженной положительно, а внешняя – отрицательно.
Возникновение возбуждения (потенциала действия) возможно лишь при сохранении достаточного количества ионов натрия в окружающей клетку среде. Большие потери натрия организмом (например, с потом при длительной мышечной работе в условиях высокой температуры воздуха) могут нарушить нормальную деятельность нервных клеток, снизить работоспособность человека. В условиях кислородного голодания тканей (например, при наличии большого кислородного долга во время мышечной работы) процесс возбуждения также нарушается из-за поражения механизма вхождения в клетку ионов натрия, и клетка становится невозбудимой.
Вопрос_16
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.
Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим.


Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.
Электрические потенциалы, возникающие в тканях и отдельных клетках человека, животных и растений, важнейшие компоненты процессов возбуждения (См. Возбуждение) и торможения (См. Торможение). Исследование Б. п. имеет большое значение для понимания физико-химических и физиологических процессов в живых системах и применяется в клинике с диагностической целью (Электрокардиография, Электроэнцефалография, Электромиография и др.).
Различают следующие основные виды Б. п. нервных и мышечных клеток: потенциал покоя, потенциал действия, возбуждающие и тормозные постсинаптические потенциалы, генераторные потенциалы.
Потенциал покоя (ПП, мембранный потенциал покоя). У живых клеток в покое между внутренним содержимым клетки и наружным раствором существует разность потенциалов (ПП) порядка 60—90мв, которая локализована на поверхностной мембране. Внутренняя сторона мембраны заряжена электроотрицательно по отношению к наружной (рис. 1). ПП обусловлен избирательной проницаемостью покоящейся мембраны для ионов К + (Ю. Бернштейн, 1902, 1912; А. Ходжкин и Б. Катц, 1947). Концентрация К + в протоплазме примерно в 50 раз выше, чем во внеклеточной жидкости, поэтому, диффундируя из клетки, ионы выносят на наружную сторону мембраны положительные заряды, при этом внутренняя сторона мембраны, практически не проницаемой для крупных органических анионов, приобретает отрицательный потенциал. Поскольку проницаемость мембраны в покое для Na + примерно в 100 раз ниже, чем для К + , диффузия натрия из внеклеточной жидкости (где он является основным катионом) в протоплазму мала и лишь незначительно снижает ПП, обусловленный ионами К + . В скелетных мышечных волокнах в возникновении потенциала покоя важную роль играют также ионы Cl - , диффундирующие внутрь клетки. Следствием ПП является ток покоя, регистрируемый между поврежденным и интактным участками нерва или мышцы при приложении отводящих электродов. Мембраны нервных и мышечных клеток (волокон) способны изменять ионную проницаемость в ответ на сдвиги мембранного потенциала. При увеличении ПП (гиперполяризация мембраны) проницаемость поверхностных клеточных мембран для Na + и К + падает, а при уменьшении ПП (деполяризация) она возрастает, причём скорость изменений проницаемости для Na + значительно превышает скорость увеличения проницаемости мембраны для К + .
Постсинаптические потенциалы (ПСП) возникают в участках мембраны нервных или мышечных клеток, непосредственно граничащих с синаптическими окончаниями. Они имеют амплитуду порядка нескольких мв и длительность 10—15 мсек. ПСП подразделяются на возбуждающие (ВПСП) и тормозные (ТПСП). ВПСП представляют собой местную деполяризацию постсинаптической мембраны, обусловленную действием соответствующего медиатора (например, ацетилхолина в нервно-мышечном соединении). При достижении ВПСП некоторого порогового (критического) значения в клетке возникает распространяющийся ПД (рис. 5, а, б). ТПСП выражается местной гиперполяризацией мембраны, обусловленной действием тормозного медиатора (рис. 5, в). В отличие от ПД, амплитуда ПСП постепенно увеличивается с увеличением количества выделившегося из нервного окончания медиатора. ВПСП и ТПСП суммируются друг с другом при одновременном или последовательном поступлении нервных импульсов к окончаниям, расположенным на мембране одной и той же клетки.
Генераторные потенциалы возникают в мембране чувствительных нервных окончаний — рецепторов. Они внешне сходны с ВПСП — их амплитуда порядка нескольких мв и зависит от силы приложенного к рецептору раздражения (рис. 6). Когда генераторный потенциал достигает порогового (критического) значения, в соседнем участке мембраны нервного волокна возникает распространяющийся ПД. Ионный механизм генераторных потенциалов ещё недостаточно изучен.
Наряду с перечисленными относительно быстро развивающимися Б. п., в нервных клетках, волокнах гладких мышц и некоторых растительных клетках регистрируются также очень медленные колебания мембранного потенциала неизвестной природы, причём на гребне волны деполяризации мембраны часто возникают разряды импульсов.
Все Б. п. могут быть зарегистрированы и точно измерены только с помощью внутриклеточных микроэлектродов, позволяющих отводить разности потенциалов между внутренней и наружной сторонами мембраны клетки. При отведении колебаний Б. п. от целых нервов, мышц или мозга с помощью поверхностных электродов регистрируется лишь суммарно потенциал множества синхронно или, чаще, асинхронно работающих клеток. Так, электромиограмма представляет собой результат сложения (интерференции) ПД множества скелетных мышечных волокон; электрокардиограмма — результирующая колебаний электрических потенциалов мышечных волокон различных отделов сердца; электроэнцефалограмма — результат суммации главным образом ВПСП и ТПСП множества клеток различных слоев коры больших полушарий. Регистрация таких интерференционных электрограмм, хотя и не позволяет анализировать колебания Б. п. отдельных клеток, имеет важное значение для суждения о состоянии исследуемого органа в целом. В клинической практике электромиограмму, электрокардиограмму и электроэнцефалограмму регистрируют с помощью электродов, расположенных на коже соответствующих частей тела. Оценка данных, полученных этими методами, основана на сопоставлении изменений характера соответствующей кривой с результатами клинических, физиологических и патологоанатомических исследований.
Лит.; Физиология человека, М., 1966; Гальвани Л. и Вольта А., Избр. работы о животном электричестве, М. — Л., 1937; Ходжкин А., Нервный импульс, пер. с англ., М., 1965; Экклс Дж., Физиология нервных клеток, пер. с англ., М., 1959; его же, Физиология синапсов, М., 1966; Катц Б., Нерв, мышца и синапс, пер. с англ., М., 1968; Ходоров Б. И., Проблема возбудимости, Л., 1969.
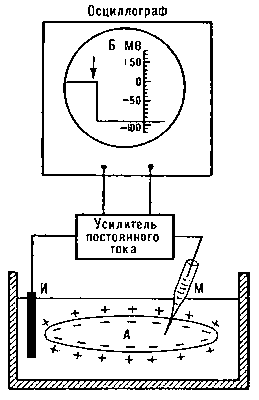
Рис. 1. Схема измерений мембранного потенциала покоя с помощью внутриклеточного стеклянного микроэлектрода (М). Второй электрод (И) помещен в омывающую клетку жидкость.
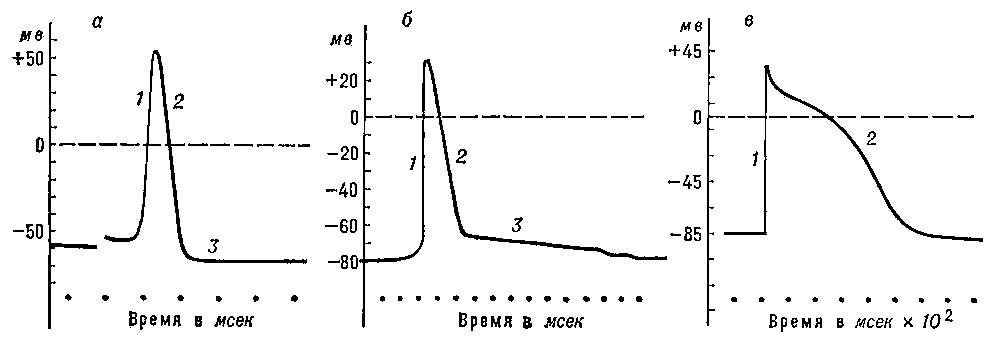
Рис. 2. Потенциалы действия, зарегистрированные с помощью внутриклеточных микроэлектродов: а — гигантского аксона кальмара; б — скелетного мышечного волокна; в — волокна мышцы сердца собаки; 1 — восходящая фаза ПД; 2 — нисходящая фаза; 3 — следовая гиперполяризация (а) и следовая деполяризация (б).
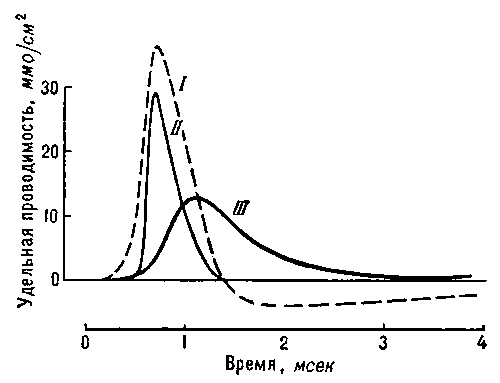
Рис. 3. Изменения натриевой и калиевой проводимости мембраны нервного волокна во время генерации потенциала действия (I). Изменения проводимости пропорциональны изменениям проницаемости для Na + (II) и К + (III).
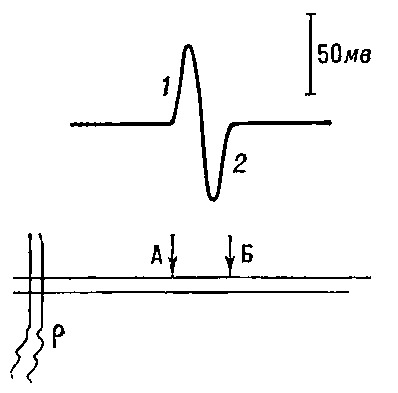
Рис. 4. Регистрация распространения потенциала действия вдоль нервного волокна. А, Б — внеклеточные электроды; р — раздражающие электроды. Вверху — отклонение луча осциллографа под влиянием волны возбуждения; 1 — волна возбуждения под электродом А; 2 — под электродом Б.
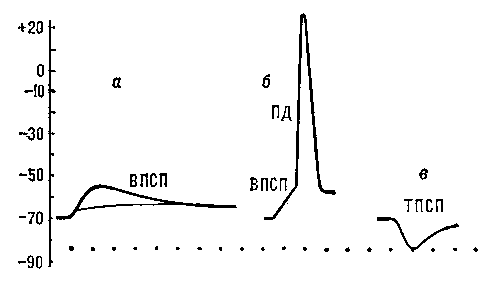
Рис. 5. Постсинаптические потенциалы: а — подпороговые ВПСП, возникающие в нервной клетке в ответ на раздражения соответствующих нервных волокон; б — ВПСП, достигший порогового значения, достаточного для возникновения ПД; в — ТПСП, вызванный раздражением тормозных нервных волокон.
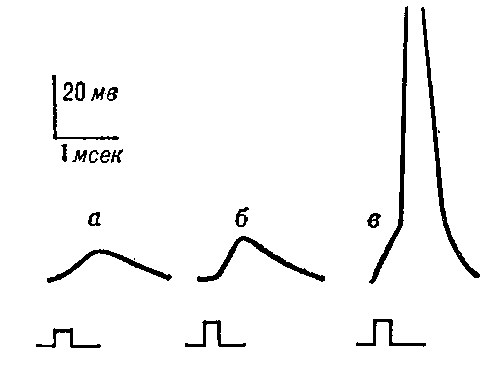
Рис. 6. Генераторные потенциалы: увеличение амплитуды при усилении раздражения рецептора (а — в). При достижении порогового значения (в) генераторный потенциал вызвал в чувствительном нервном волокне потенциал действия.
Потенциал действия нервной клетки
- Потенциал действия нервной клетки
- Ионные механизмы потенциала действия
- Проведение возбуждения по нервным волокнам
- Законы проведения возбуждения
- Типы нервных волокон и их функции
- Все страницы
Проведение ПД по мембране можно сравнить с поджиганием пороховой дорожки: вспыхнувший порох немедленно воспламеняет впереди лежащие частицы, и пламя движется вперёд до конца дорожки.
Продолжительность потенциала действия нервной клетки измеряется единицами миллисекунд (мс).
Потенциалы действия, зарегистрированные двумя электродами, один из которых находится внутри клетки, а другой — в окружающем растворе, представлены на рис. 5-3 и 5-7.
Рис. 5–3. Изменения мембранного потенциала и потенциал действия. Вертикальная стрелка в нижней части рисунка — момент появления раздражающего стимула, на отметке –80 мВ — исходный уровень МП.
Между моментом нанесения раздражения и первым проявлением ПД имеется задержка — латентный период. Латентный период соответствует времени, когда ПД движется по мембране нервной клетки от места раздражения до отводящего электрода. Под действием раздражающего стимула происходит нарастающая деполяризация мембраны — локальный ответ. При достижении критического уровня деполяризации, который в среднем составляет —55 мВ, начинается фаза деполяризации. В эту фазу уровень МП падает до нуля и даже приобретает положительное значение (овершут), а затем возвращается к исходному уровню (фазареполяризации). Фазы деполяризации, овершута и реполяризации образуют спайк (пик) ПД. Длительность спайка составляет 1—2 мс. После спайка наблюдается замедление скорости спада потенциала — (раза следовой деполяризации. После достижения исходного уровня покоя нередко наблюдается фаза следовой гиперполяризации. Эти следовые потенциалы могут длиться десятки и сотни миллисекунд.
В основе изменений мембранного потенциала (МП), происходящих в течение потенциала действия (ПД), лежат ионные механизмы. На рис. 5—7 представлены суммарные ионные токи, протекающие через мембрану нервной клетки в ходе потенциала действия.
Рис. 5-7. Потенциал действия и ионные токи нервной клетки [5|. По оси ординат отложены значения МП (мВ), по оси абсцисс — время (мс)
Локальный потенциал. Раздражение клетки приводит к открытию части Na+-каналов и появлению локального (нераспространяющегося) потенциала.
Фаза деполяризации. При достижении критического уровня деполяризации мембраны (соответствует порогу активации потен-циалозависимых Na+-каналов) начинается лавинообразный процесс открытия большого количества Na+-каналов. В фазу деполяризации происходит массивный вход в клетку ионов Na+ по концентрационному и электрохимическому градиентам.
Овершут. Деполяризация мембраны приводит к реверсии МП (МП становится положительным). В фазу овершута Na+-ток начинает стремительно спадать, что связано с инактивацией потенциало-зависимых Na+-каналов (время открытого состояния –– доли миллисекунды) и исчезновением электрохимического градиента Na+.
Фаза реполяризации. Помимо инактивации Na+-каналов, развитию реполяризации способствует открытие потенциалозависимых K+-каналов. Этот процесс происходит медленнее, чем открытие Na+-каналов, но K+-каналы остаются открытыми более продолжительное время. Выход К+ наружу способствует полному завершению фазы реполяризации.
Следовые потенциалы связаны с длительными изменениями кинетических свойств К+-каналов. Восстановление исходного уровня МП приводит Na+- и K+-каналы в состояние покоя.
В ходе развития ПД происходят изменения возбудимости мембраны нервной клетки.
Абсолютно рефрактерный период. Во время фазы деполяризации и большей части фазы реполяризации ПД клетка находится в абсолютно рефрактерном периоде, в течение которого даже сверхпороговое раздражение не способно вызвать ПД. Этот феномен связан с инактивацией большинства Na+-каналов.
Относительно рефрактерный период. В конце фазы реполяризации, а также во время следовой гиперполяризации клетка способна генерировать ПД только в ответ на сверхпороговые раздражители. Это связано со значительным реполяризующим действием выходящих калиевых токов.
Наличие рефрактерности ограничивает частоту генерации ПД. Физиологическое значение рефрактерности заключается в создании условий для своевременного и полного осуществления восстановительных процессов в нервной клетке. Феномен рефрактерности лежит в основе понятия о функциональной подвижности, или лабильности (Н.Е. Введенский [1] ).
Лабильность — максимально возможная частота генерации ПД для данного типа возбудимой клетки. Лабильность большинства нейронов составляет приблизительно 400 ПД/с, а у интернейронов спинного мозга доходит до 1000 ПД/с.
В нормальных условиях рефрактерность предохраняет нервные клетки от излишне частой генерации ПД. Мутации, гипоксия, механические травмы и другие патологические воздействия приводят к значительным изменениям возбудимости нейронов. Такие нейроны или группы нейронов являются потенциальными источниками возникновения пароксизмальных состояний ЦНС, эпилептических припадков и других неврологических расстройств.
Нервные волокна — аксоны нервных клеток, окружённые оболочкой из олигодендроглиоцитов в ЦНС и шванновских [2] клеток в периферических нервах. Нервные волокна подразделяют на 2 типа — безмиелиновые и миелиновые. Основная функция нервных волокон — проведение ПД. Скорость проведения в миелиновых и безмиелиновых волокнах различна (рис. 5–8) и существенно зависит от диаметра нервных волокон.
Рис. 5–8. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокнах разного диаметра [4]. Скорость проведения пропорциональна диаметру нервного волокна и в миелиновых волокнах выше, чем в безмиелиновых.
Безмиелиновые нервные волокна (рис. 5–9А). В покое мембрана аксона (осевого цилиндра) поляризована — положительно заряжена снаружи и отрицательно внутри. При ПД полярность изменяется, и наружная поверхность мембраны приобретает отрицательный заряд. Из-за разности потенциалов между возбуждённым и невозбуждёнными сегментами возникают локальные токи, деполяризующие соседний участок мембраны. Теперь этот участок становится возбуждённым и деполяризует следующий участок мембраны.
Рис. 5–9. Проведение возбуждения в нервных волокнах [7]. А — безмиелиновое волокно (электротоническое проведение), Б — миелиновое волокно (скачкообразное проведение). Миелин, полностью окружая аксон в межузловых промежутках, выступает в роли электрического изолятора, а межклеточная жидкость в перехватах Ранвье [3] — проводник.
Появление так называемых рефрактерных каналов (рефрактерное состояние мембраны после прохождения ПД) предупреждает распространение возбуждения в обратном направлении.
Скорость проведения возбуждения по безмиелиновому нервному волокну в основном составляет 0,5–2 м/с и зависит от диаметра волокна: чем больше диаметр, тем выше скорость проведения ПД (см. рис. 5–8).
Миелиновое нервное волокно (рис. 5–9Б) состоит из осевого цилиндра (аксона), вокруг которого шванновские клетки образуют миелин за счёт концентрического наслаивания собственной плазматической мембраны. Миелин прерывается через регулярные промежутки (от 0,2 до 2 мм) концентрической щелью шириной около 1 мкм, это узлы, или перехваты Ранвье. Таким образом, межузловые сегменты аксона, расположенные между соседними перехватами Ранвье, содержат миелин — электрический изолятор, не позволяющий проходить через него локальным токам, поэтому ПД возникают только в перехватах Ранвье. Другими словами, ПД перемещается вдоль нервного волокна скачками, от одного перехвата Ранвье к другому перехвату (скачкообразное проведение).
Плотность потенциалозависимых Na+?каналов аксолеммы в перехватах Ранвье — до 2000 на 1 мкм2 (в перикарионе — 50–70, в начальном сегменте аксона — 2000, в межузловых сегментах Na+?каналы практически отсутствуют). В силу высокой плотности Na+?каналов перехваты Ранвье характеризуются высокой возбудимостью, а локальные токи достаточно велики для возбуждения соседнего перехвата.
Локальные токи текут от перехвата к перехвату (через внеклеточную жидкость кнаружи от миелина и через аксоплазму внутри аксона) с минимальными потерями.
Энергозатраты нервного волокна на проведение ПД относительно невелики, поскольку возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% общей поверхности мембраны аксона. Поэтому даже после длительных ритмических пачек ПД трансмембранный градиент концентраций ионов практически не изменяется.
В физиологических условиях ПД движутся в одном направлении от места раздражения (ортодромное проведение). ПД, проходящий по нервному волокну, возбуждает следующий, но не предыдущий участок мембраны. Это связано с рефрактерностью предыдущего участка после возбуждения. Проведение в противоположном направлении (антидромное проведение) возможно при травматическом поражении нервных волокон и в редких случаях (аксон–рефлекс).
Нарушение миелинизации нервных волокон приводит к нарушениям проводимости (демиелинизирующие заболевания). При разрушении миелиновой оболочки происходит резкое снижение скорости и надёжности проведения возбуждения по нервам. Наиболее распространённым среди демиелинизирующих заболеваний является множественный склероз, проявляющийся различными параличами и потерей чувствительности.
Бездекрементное проведение возбуждения. Амплитуда ПД в различных участках нерва одинакова, то есть проведение возбуждения по нервному волокну осуществляется без затухания (бездекрементно). Таким образом, кодирование информации осуществляется не за счёт изменения амплитуды ПД, а путём изменения их частоты и распределения во времени.
Изолированное проведение возбуждения. Нервные стволы обычно образованы большим количеством нервных волокон, однако ПД, идущие по каждому из них, не передаются на соседние. Эта особенность нервных волокон обусловлена:
- наличием оболочек, окружающих отдельные нервные волокна и их пучки (в результате образуется барьер, предупреждающий переход возбуждения с волокна на волокно);
- сопротивлением межклеточной жидкости (жидкость, находящаяся между волокнами, имеет гораздо меньшее сопротивление току, чем мембрана аксонов; поэтому ток шунтируется по межволоконным пространствам и не доходит до соседних волокон).
Физиологическая и анатомическая целостность. Необходимым условием проведения возбуждения является не только его анатомическая целостность, но и нормальное функционирование мембраны нервного волокна (физиологическая целостность). В клинике широко применяют различные ЛС, нарушающие физиологическую целостность нервных волокон. Так, эффекты местных анестетиков (новокаин, лидокаин, и др.) основаны на блокаде потенциалозависимых Na+?каналов. Нарушение физиологической целостности чувствительных нервных волокон вызывает анестезию (потерю чувствительности).
При регистрации электрической активности нервного ствола Джозеф Эрлангер и Герберт Гассер в 1937 г. обнаружили составной характер тока действия нервного ствола. На основании полученных данных (диаметр, скорость проведения, функция) разработана классификация (табл. 5—1), в соответствии с которой нервные волокна подразделены на группы А, В и С с дальнейшими градациями ( ? , ? , и т.д.).
Орлов Р.С., Ноздрачёв А.Д. Нормальная физиология : Учебник. - М.: ГЭОТАР-Медиа, 2009. 688 с. - Глава 5. Физиология нейронов. - Потенциал действия. С. 73-78.
[2] Шванн Теодор (Schwann T.), немецкий гистолог и физиолог (1810–1882); вместе с М. Шульце создал клеточную теорию (1839 г.); в 1836 г. открыл пепсин, в 1838 г. опубликовал первую работу по строению миелиновой оболочки.
[3] Ранвье Луи (Ranvier L.), французский патолог (1835–1922). Занимался изучением нервной ткани с применением азотнокислого серебра и хлорного золота. Его именем названы безмиелиновые участки (узловые перехваты) миелинового нервного волокна.
Нервный импульс
Строго говоря, движущееся по нервам возбуждение представляет собой нервные импульсы, а не потенциалы действия.
Можно сказать короче:
Но в физиологической литературе в качестве синонима для нервного импульса принято использовать также и термин "потенциал действия". Хотя потенциал действия - это только электрический компонент нервного импульса.
Потенциал действия – это резкое скачкообразное изменение мембранного потенциала с отрицательного на положительный и обратно.
Потенциал действия - это электрическая характеристика (электрическая составляющая) нервного импульса.
Нервный импульс - это сложный структурно-электро-химический процесс, распространяющийся по мембране нейрона в виде бегущей волны изменений.
Потенциал действия - это только электрический компонент нервного импульса, характеризующий изменения электрического заряда (потенциала) на локальном участке мембраны во время прохождения через него нервного импульса (от -70 до +30 мВ и обратно). (Кликните на изображение слева, чтобы увидеть анимацию.)
Сравните два приведённых выше рисунка (покликайте по ним) и, как говорится, почувствуйте разницу!
Где рождаются нервные импульсы?
Как ни странно, не все студенты, изучившие физиологию возбуждения, могут ответить на этот вопрос. ((
Хотя ответ не сложен. Нервные импульсы рождаются на нейронах всего в нескольких местах:
1) аксонный холмик (это переход тела нейрона в аксон),
2) рецепторное окончание дендрита,
3) первый перехват Ранвье на дендрите (триггерная зона дендрита),
4) постсинаптическая мембрана возбуждающего синапса.
Места возникновения нервных импульсов:
1. Аксонный холмик - главный породитель нервных импульсов.
Аксонный холмик - это самое начало аксона, там где он начинается на теле нейрона. Именно аксонный холмик является главным породителем (генератором) нервных импульсов на нейроне. Во всех остальных местах вероятность рождения нервного импульса намного меньше. Дело в том, что у мембраны аксонного холмика повышена чувствительность к возбуждению и понижен критический уровень деполяризации (КУД) по сравнению с остальными участками мембраны. Поэтому, когда на мембране нейрона начинают суммироваться многочисленные возбуждающие постсинаптические потенциалы (ВПСП), которые возникают в самых разных местах на постсинаптических мембранах всех его синаптических контактов, то раньше всего КУД достигается именно на аксонном холмике. Там-то эта сверхпороговая для холмика деполяризация и открывает потенциал-чувствительные натриевые каналы, в которые входит поток ионов натрия, порождающий потенциал действия и нервный импульс.
Итак, аксонный холмик является интегративной зоной на мембране, он интегрирует все возникающие на нейроне локальные потенциалы (возбуждающие и тормозные) - и первый срабатывает на достижение КУД, порождая нервный импульс.
Важно также учесть следующий факт. От аксонного холмика нервный импульс разбегается по всей мембране своего нейрона: как по аксону к пресинаптическоим окончаниям, так и по дендритам к постсинаптическим "начинаниям". Все локальные потенциалы при этом снимаются с мембраны нейрона и со всех его синапсов, т.к. они "перебиваются" потенциалом действия от пробегающего по всей мембране нервного импульса.
2. Рецепторное окончание чувствительного (афферентного) нейрона.
Если нейрон имеет рецепторное окончание, то на него может воздействовать адекватный раздражитель и порождать на этом окончании сначала генераторный потенциал, а затем и нервный импульс. Когда генераторный потенциал достигает КУД, то на этом окончании открываются потенциал-зависимые натриевые ионные каналы и рождается потенциал действия и нервный импульс. Нервный импульс бежит по дендриту к телу нейрона, а затем по его аксону к пресинаптическим окончаниям для передачи возбуждения на следующий нейрон. Так работают, к примеру, болевые рецепторы (ноцицепторы), являющиеся дендритными окончаниями болевых нейронов. Нервные импульсы в болевых нейронах вознимают именно на рецепторных окончаниях дендритов.
3. Первый перехват Ранвье на дендрите (триггерная зона дендрита).
Локальные возбуждающие постсинаптические потенциалы (ВПСП) на окончаниях дендрита, которые формируются в ответ на возбуждения, приходящие к дендриту через синапсы, суммируются на первом перехвате Ранвье этого дендрита, если он, конечно, миелинизирован. Там находится участок мембраны с повышенной чувствительностью к возбуждению (пониженным порогом), поэтому именно в этом участке легче всего преодолевается критический уровень деполяризации (КУД), после чего открываются потенциал-управляемые ионные каналы для натрия - и возникает потенциал действия (нервный импульс).
4. Постсинаптическая мембрана возбуждающего синапса.
В редких случаях ВПСП на возбуждающем синапсе может быть настолько силён, что прямо там же достигает КУД и порождает нервный импульс. Но чаще это бывает возможно только в результате суммации нескольких ВПСП: или с нескольких соседних синапсов, сработавших одновременно (пространственная суммация), или за счёт того, что на данный синапс пришло несколько импульсов подряд (временная суммация).
Видео: Проведение нервного импульса по нервному волокну
Потенциал действия как нервный импульс
Ниже размещён материал, взятый из учебно-методического пособия автора данного сайта, на который вполне можно ссылаться в своём списке литературы:
Сазонов В.Ф. Понятие и виды торможения в физиологии центральной нервной системы: Учебно-методическое пособие. Ч. 1. Рязань: РГПУ, 2004. 80 с.
Все процессы мембранных изменений, происходящих в ходе распространяющегося возбуждения, достаточно хорошо изучены и описаны в научной и учебной литературе. Но не всегда это описание легко понять, поскольку в данном процессе задействовано слишком много компонентов (с точки зрения обычного студента, а не вундеркинда, конечно).
Для облегчения понимания мы предлагаем рассматривать единый электрохимический процесс распространяющегося динамичного возбуждения с трех сторон, на трех уровнях:
Электрические явления – развитие потенциала действия.
Химические явления – движение ионных потоков.
Структурные явления – поведение ионных каналов.
Три стороны процесса распространяющегося возбуждения
1. Потенциал действия (ПД)
Потенциал действия – это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно.
Обычно мембранный потенциал в нейронах ЦНС изменяется от –70 мВ до +30 мВ, а затем вновь возвращается к исходному состоянию, т.е. к –70 мВ. Как видим, понятие потенциала действия характеризуется через электрические явления на мембране.
На электрическом уровне изменения начинаются как смена поляризованного состояния мембраны на деполяризацию. Сначала деполяризация идет в виде локального возбуждающего потенциала. Вплоть до критического уровня деполяризации (примерно –50 мВ) это относительно простое линейное уменьшение электроотрицательности, пропорциональное силе воздействующего раздражителя. А вот потом начинается более крутая самоусиливающаяся деполяризация, она развивается не с постоянной скоростью, а с ускорением . Говоря образно, деполяризация так разгоняется, что перескакивает через нулевую отметку, не заметив этого, и даже переходит в положительную поляризацию. После достижения пика (обычно +30 мВ) начинается обратный процесс – реполяризация , т.е. восстановление отрицательной поляризации мембраны.
Кратко опишем электрические явления во время течения потенциала действия:
Восходящая ветвь графика:
потенциал покоя – исходное обычное поляризованное электроотрицательное состояние мембраны (–70 мВ);
нарастающий локальный потенциал – пропорциональная раздражителю деполяризация;
критический уровень деполяризации (–50 мВ) – резкое ускорение деполяризации (за счет самораскрытия натриевых каналов), с этой точки начинается спайк – высокоамплитудная часть потенциала действия;
самоусиливающаяся круто нарастающая деполяризация;
переход нулевой отметки (0 мВ) – смена полярности мембраны;
пик (+30 мВ) – вершина процесса изменения полярности мембраны, вершина потенциала действия.
Нисходящая ветвь графика:
реполяризация – восстановление прежней электроотрицательности мембраны;
переход нулевой отметки (0 мВ) – обратная смена полярности мембраны на прежнюю, отрицательную;
переход критического уровня деполяризации (–50 мВ) – прекращение фазы относительной рефрактерности (невозбудимости) и возврат возбудимости;
следовые процессы (следовая деполяризация или следовая гиперполяризация);
восстановление потенциала покоя – норма (–70 мВ).
Итак, сначала – деполяризация, затем – реполяризация. Сначала – утрата электроотрицательности, затем – восстановление электроотрицательности.
2. Ионные потоки
Итак, важно осознать, что все электрические токи, которые идут через мембрану, являются ионными потоками . Привычного нам из физики тока в виде потока электронов в клетках, как в водных системах, просто нет. Ссылки на потоки электронов будут ошибкой.
На химическом уровне мы, описывая распространяющееся возбуждение, должны рассмотреть, как изменяются характеристики ионных потоков, идущих через мембрану. Главное в этом процессе то, что при деполяризации резко усиливается поток ионов натрия внутрь клетки, а затем он внезапно прекращается на спайке потенциала действия. Входящий поток натрия как раз и вызывает деполяризацию, так как ионы натрия приносят с собой положительные заряды в клетку (чем и снижают электроотрицательность). Затем, после спайка, значительно нарастает выходящий наружу поток ионов калия, что вызывает реполяризацию. Ведь калий, как мы неоднократно говорили, выносит с собой из клетки положительные заряды. Отрицательные заряды остаются внутри клетки в большинстве, и за счет этого усиливается электроотрицательность. Это и есть восстановление поляризации за счет выходящего потока ионов калия. Заметим, что выходящий поток ионов калия возникает практически одновременно с появлением натриевого потока, но нарастает медленно и длится в 10 раз дольше. Несмотря на продолжительность калиевого потока самих ионов расходуется немного – всего одна миллионная доля от запаса калия в клетке (0,000001 часть).
Подведем итоги. Восходящая ветвь графика потенциала действия образуется за счет входа в клетку ионов натрия, а нисходящая – за счет выхода из клетки ионов калия.
3. Ионные каналы
Все три стороны процесса возбуждения – электрическая, химическая и структурная – необходимы для понимания его сущности. Но все-таки все начинается с работы ионных каналов. Именно состояние ионных каналов предопределяет поведение ионов, а поведение ионов в свою очередь сопровождается электрическими явлениями. Начинают процесс возбуждения натриевые каналы .
Пожар как метафора распространяющегося возбуждения
Когда мы искали образный пример, аналогию или метафору, которая может наглядно передать смысл распространяющегося возбуждения, то остановились на образе пожара. Действительно, распространяющееся возбуждение похоже на лесной пожар, когда горящие деревья остаются на месте, а фронт огня распространяется и уходит все дальше и дальше во все стороны от очага возгорания.
Как же в этой метафоре будет выглядеть явление торможения?
Ответ очевиден – торможение будет выглядеть как тушение пожара, как уменьшение горения и затухание огня. Но если огонь распространяется сам по себе, то тушение требует усилий. Из потушенного участка процесс тушения сам по себе не пойдет во все стороны.
Существует три варианта борьбы с пожаром: (1) либо надо ждать, когда все сгорит и огонь истощит все горючие запасы, (2) либо надо поливать водой горящие участки, чтобы они погасли, (3) либо надо поливать заранее ближайшие нетронутые огнем участки, чтобы они не загорелись.
АВТОВОЛНЫ В АКТИВНО-ВОЗБУДИМЫХ СРЕДАХ (АВС)
При распространении волны в активно-возбудимых средах не происходит переноса энергии. Энергия не переносится, а освобождается, когда до участка АВС доходит возбуждение. Можно провести аналогию с серией взрывов зарядов, заложенных на некотором расстоянии друг от друга (например, при тушении лесных пожаров, строительстве, мелиоративных работах), когда взрыв одного заряда вызывает взрыв рядом расположенного и так далее. Лесной пожар также является примером распространения волны в активно- возбудимой среде. Пламя распространяется по области с распределенными запасами энергии - деревья, валежник, сухой мох.
Основные свойства волн, распространяющихся в активно-возбудимых средах (АВС)
Волна возбуждения распространяется в АВС без затухания; прохождение волны возбуждения связано с рефрактерностью - невозбудимостью среды в течение некоторого промежутка времени (периода рефрактерности).
Видео: Потенциал действия (Action potential)
Читайте также:
