
Компенсация функций и пластичность в нервных центрах
Нервный центр - центральный компонент рефлекторной дуги, где происходит переработка информации, вырабатывается программа действия, формируется эталон результата.
1. Односторонность проведения возбуждения. В рефлекторной дуге, включающей нервные центры, процесс возбуждения распространяется в одном направлении (от входа, афферентных путей к выходу, эфферентным путям).
2. Иррадиация возбуждения. Особенности структурной организации центральных нейронов, огромное число межнейронных соединений в нервных центрах существенно модифицируют направление распространения процесса возбуждения в зависимости от силы раздражителя и функционального состояния центральных нейронов. Значительное увеличение силы раздражителя приводит к расширению области вовлекаемых в процесс возбуждения центральных нейронов -- иррадиации возбуждения.
3. Суммация возбуждения.
4. Наличие синаптической задержки.
5. Высокая утомляемость.
7. Пластичность. Функциональная возможность нервного центра существенно модифицировать картину осуществляемых рефлекторных реакций. Поэтому пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами.
8. Конвергенция. Нервные центры высших отделов мозга являются мощными коллекторами, собирающими разнородную афферентную информацию.
9. Интеграция в нервных центрах.
10. Свойство доминанты. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в центральной нервной системе.
11. Цефализация нервной системы. Основная тенденция в эволюционном развитии нервной системы проявляется в перемещении, сосредоточении функции регуляции и координации деятельности организма в головных отделах ЦНС. Этот процесс называется цефализацией управляющей функции нервной системы.
Вегетативная (синонимы: автономная, ганглионарная, органная, висцеральная, чревная) нервная система - это отдел нервной системы, который регулирует деятельность внутренних органов, желез, гладких мышц, некоторых поперечно-полосатых, сосудов, обмен веществ, отвечает за регуляцию гомеостаза. - не подчинена воле человека
Автономная нервная система состоит из двигательных волокон и не имеет собственных чувствительных. Состоит из двигательных нейронов двух типов: преганглиолярных (тела лежат в головном или спинном мозге, а отросток выходит за их пределы и заканчивается в периферическом нервном узле.) и постганглиолярных (отросток, направленный к рабочему органу). Периферические волокна автономной нервной системы тоньше волокон соматической, и скорость проведения импульса по ним значительно медленнее.
Высшим центром контроля вегетативных функций является гипоталамус. От него сигналы поступают непосредственно в проводящие элементы автономной нервной системы. Кора больших полушарий осуществляет высший контроль и регуляцию функций автономной нервной системы.
На основании топографии вегетативных ядер и узлов, различий в длине аксонов первого и второго нейронов эфферентного пути, а также особенностей функции вегетативная нервная система подразделяется на симпатическую, парасимпатическую и метасимпатическую.
Симпатическая нервная система активируется при стрессовых реакциях, во время интенсивной работы. Ганглии расположены на значительном расстоянии от иннервируемых органов. Для неё характерно генерализованное влияние, при этом симпатические волокна иннервируют подавляющее большинство органов. Симпатические нервы выполняют роль трофических нервов, регуляторов обмена веществ в сердечной мышце и скелетной мускулатуре.
Парасимпатическая нервная система (околосимпатическая) способствует восстановлению запасов энергии во время сна и отдыха. В парасимпатической нервной системе ганглии расположены непосредственно в органах или на подходах к ним. Волокна парасимпатического отела входят в состав нервов гладких мышц глаза, слюнных желез, органов шеи, грудной клетки и брюшной полости. Известно, что парасимпатическая стимуляция одних органов оказывает тормозное действие, а других -- возбуждающее действие. В большинстве случаев действие парасимпатической и симпатической систем противоположно (исключение -- действие на слюнные железы, где и симпатическая, и парасимпатическая нервная система вызывают активацию желез).
Такое воздействие с двух разных отделов вегетативной нервной системы обеспечивает четкость работы органов и их систем в организме, определяет приспособляемость организма к изменяющимся условиям окружающей среды. В-целом симпатическая нервная система обеспечивает мобилизацию резервов, необходимых для активного взаимодействия со средой, растрачивания энергии, а парасимпатическая - восстановления ее.
Метасимпатическая нервная система -- часть автономной нервной системы, комплекс микроганглионарных образований (интрамуральных ганглиев) и соединяющих их нервов, а также отдельные нейроны и их отростки, расположенные в стенках внутренних органов, которые обладают сократительной активностью. Основными эффекторными аппаратами стенок полых висцеральных органов, которые регулируются МНС, являются: гладкая мышца, секреторный, всасывающий и экскреторный эпителий, капиллярная сеть, местные эндокринные и иммунные образования. Характеризуется высокой степенью относительной независимости от центральной нервной системы.
Метасимпатическая нервная система обеспечивает передачу возбуждения с эсктраорганной нервной системы на ткань органа и является посредником между симпатической и парасимпатической нервными системами и тканью органа. Метасимпатическая нервная система регулирует органный кровоток, а также имеет имеет непосредственное отношение к организации регулирующих влияний на такую важную функцию, как мембранное пищеварение.
Под контролем автономной системы находятся органы кровообращения, дыхания, пищеварения, выделения, размножения, а также обмен веществ и рост. Фактически эфферентный отдел ВНС осуществляет нервную регуляцию функций всех органов и тканей, кроме скелетных мышц, которыми управляет соматическая нервная система.
В отличие от соматической нервной системы, двигательный эффекторный нейрон в автономной нервной системе находится на периферии, и спинной мозг лишь косвенно управляет его импульсами. ВНС (вегетативная нервная система) приспосабливает работу внутренних органов к изменениям окружающей среды. ВНС обеспечивает гомеостаз (постоянство внутренней среды организма). ВНС также участвует во многих поведенческих актах, осуществляемых под управлением головного мозга, влияя не только на физическую, но и на психическую деятельность человека.
Нейроны ЦНС для осуществления сложных и многообразных функций объединяются в нервные центры. Нервный центр - это совокупность нейронов, принимающих участие в осуществлении конкретного рефлекса (мигания, глотания, кашля и т. д.) . В целом организме при формировании сложных адаптивных процессов происходит функциональное объединение нейронов, расположенных на различных уровнях ЦНС. Такое объединение (нервный центр в широком смысле слова) позволяет осуществлять наиболее адекватное для конкретных условий осуществления рефлекторной деятельности.
Нервные центры обладают рядом характерных функциональных свойств, обусловленных объединением нейронов в нейронные сети и наличием межнейрональных синапсов.
Основные из этих свойств:
1. Возбуждение в нервных центрах распространяется односторонне - от рецептора к эффектору, что обусловливается свойством химических синапсов односторонне проводить возбуждение от пресинаптической мембраны к постсинаптической.
2. Возбуждение в нервных центрах проводится медленнее, чем по нервному волокну. Это обусловлено замедленным проведением возбуждения через синапс (синаптическая задержка) .
3. В нервных центрах осуществляется суммация возбуждений. Различают два вида суммации:
• временная или последовательная, если импульсы возбуждения приходят к нейрону по одному и тему же пути через один синапс с интервалом меньше, чем время полной реполяризации постсинаптической мембраны. В этих условиях ВПСП на постсинаптической мембране суммируются и доводят ее деполяризацию до уровня, достаточного для генерации нейроном потенциала действия;
• пространственная или одновременная - наблюдается в том случае, когда импульсы возбуждения поступают к нейрону одновременно через разные синапсы
4. Трансформация ритма возбуждения - изменение количества импульсов возбуждения, выходящих из нервного центра, по сравнению с числом импульсов, приходящих к нему. Различают два вида трансформации:
• понижающая трансформация, в основе которой, в основном, лежит явление суммации возбуждений (пространственной и временной), когда в ответ на несколько возбуждений, пришедших к нервной клетке, в последней возникает только одно возбуждение;
• повышающая трансформация, в ее основе лежат механизмы умножения (мультипликации), способные резко увеличить количество импульсов возбуждения
5. Рефлекторное последействие - заключается в том, что рефлекторная реакция заканчивается позже прекращения действия раздражителя. Это явление обусловлено двумя причинами:
• длительной следовой деполяризацией мембраны нейрона, на фоне которой могут возникать несколько потенциалов действия, обеспечивающих кратковременное рефлекторное последействие;
• пролонгированием выхода возбуждения к эффектору в результате циркуляции (реверберации) возбуждения в нейронной сети типа "нейронной ловушки". Возбуждение, попадая в такую сеть, может длительное время циркулировать в ней, обеспечивая длительное рефлекторное последействие. Возбуждение в такой цепочке может циркулировать до тех пор, пока какое-либо внешнее воздействие затормозит этот процесс или в ней наступит утомление.
6. Нервные центры, как и синапсы, обладают высокой чувствительностью к недостатку кислорода.
7. Нервные центры, как и синапсы, обладают высокой чувствительностью к действию различных химических веществ, особенно ядов. На одном нейроне могут располагаться синапсы, обладающие различной чувствительностью к различным химическим веществам.
Поэтому можно подобрать такие химические вещества, которые избирательно будут блокировать одни синапсы, оставляя другие в рабочем состоянии. Это делает возможным корректировать состояния и реакции как здорового, так и больного организма.
8. Нервные центры, как и синапсы, обладают быстрой утомляемостью в отличие от нервных волокон, которые считаются практически неутомляемыми.
9. Нервные центры, как и синапсы, обладают низкой лабильностью.
10. В нервных центрах легко возникает процесс торможения.
11. Нервные центры обладают тонусом, который выражается в том, что даже при отсутствии специальных раздражении, они постоянно посылают импульсы к рабочим органам.
12. Нервные центры, как и синапсы, обладают низкой аккомодационной способностью, т. е. они способны реагировать на раздражающие факторы, медленно нарастающие по силе.
13. Нервные центры обладают пластичностью - способностью изменять собственное функциональное назначение и расширять свои функциональные возможности.
14. Посттетаническая потенциация - явление усиления рефлекторного ответа после длительного ритмического раздражения нервного центра. Оно обусловлено сохранением определенного уровня ВПСП на нейронах центра, что облегчает проведение последующих возбуждений через синапсы.
Под нервным центром понимают сложный комплекс (ансамбль) нейронов, совокупно участвующих в регуляции какой-то физиологической функции или осуществлении определенного рефлекторного акта. Существует "понимание" нервных центров в узком и широком смысле этого понятия. Под нервными центрами в узком смысле понимают лишь те структуры центральной нервной системы, которые являются жизненно необходимыми для осуществления или регуляции определенного физиологического процесса, тогда как нервный центр в широком смысле этого понятия представляет собой совокупность всех компонентов центральной нервной системы, имеющих отношение к осуществлению и регуляции определенного физиологического процесса. Так, например, жизненно важный дыхательный центр, обеспечивающий ритмическую смену вдоха выдохом, находится в продолговатом мозге, но в то же время сами мотонейроны, иннервирующие дыхательную мускулатуру, заложены в спинном мозге. Кроме того, нервные клетки, имеющие отношение к регуляции внешнего дыхания, находятся в мосте, гипоталамусе (части промежуточного мозга) и коре больших полушарий. В связи с отмеченным под дыхательным центром в узком смысле этого слова понимают лишь структуры продолговатого мозга, имеющие отношение к периодической смене вдоха выдохом, тогда как в широком смысле этого понятия – совокупность всех нервных элементов центральной нервной системы, имеющих отношение к регуляции дыхания.
Для всех нервных центров характерны следующие свойства:
1. Одностороннее проведение возбуждения (возбуждение проводится всегда в одном направлении от афферентного нейрона к эфферентному, а затем к структурам исполнительных органов). Достигается благодаря существованию синапсов химического типа между чувствительными, вставочными и эфферентными нейронами.
2. Замедленное проведение возбуждения через нервные центры. Обусловлено передачей информации от одних нейронов центральной нервной системы к другим, имеющим отношение к регуляции какой-то физиологической функции, преимущественно через химические синапсы, для которых характерна синаптическая задержка (0,3-0,5 мс при проведении информации через один синапс). Соответственно, чем больше синаптических контактов встречается на пути нервного импульса от чувствительного нейрона к эфферентному, тем длительнее будет общая задержка проведения через соответствующий нервный центр.
3. Зависимость рефлекторного ответа от силы и длительности раздражения рецептивного поля рефлекса. Существование такой связи обусловлено тем, что, чем сильнее действующий на рецептивное поле какого-то рефлекторного акта раздражитель и чем больше длительность его действия, тем соответственно больше будет количество элементов нервного центра, вовлекаемых в реакцию, и большим окажется количество структур иннервируемых рабочих органов, участвующих в рефлекторном ответе, а, следовательно, и выше будет амплитуда рефлекторного ответа.
4. Суммация возбуждений в нервном центре. Это свойство проявляется следующим образом: раздражение каких-то отдельных рецепторов рецептивного поля нервного центра слабыми раздражителями не вызывает возникновения рефлекторного ответа, тогда как последовательное с определенной частотой раздражение этого же рецептора или одновременное воздействие на несколько соседних рецепторных элементов теми же слабыми стимулами, приводит к возникновению рефлекторного ответа. В основе описанного феномена лежат явления суммации возбуждений, поступающих к одному и тому же центральному нейрону либо через какой-то один из его синаптических входов с определенной частотой (временная суммация), либо одновременно через несколько его синаптических входов (пространственная суммация).
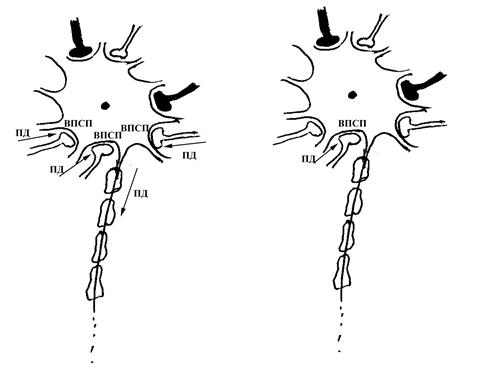
Рис. 15. Схема пространственной суммации информации, поступающей к мембране тела нейрона по различным его синаптическим входам. Обратите внимание на то, что на рисунке схематически изображен нейрон, на теле которого расположено большое количество синапсов (незатушеванные – возбуждающие, затушеванные – тормозные). На рисунке А показан такой вариант, когда к пресинаптическим полюсам сразу нескольких синапсов (в данном случае трех возбуждающих синапсов) одновременно приходит возбуждение, которое инициирует возникновение в постсинаптической мембране этих синапсов (являющейся частью мембраны тела рассматриваемого нейрона) возбуждающих постсинаптических потенциалов (ВПСП), суммирующихся в пространстве. Суммарный возбуждающий постсинаптический потенциал, возникающий в результате такой суммации, является достаточным для инициации потенциала действия (ПД) в области аксонного холмика данного нейрона.
На рисунке Б показан вариант, когда возбуждение приходит только к одному из пресинаптических полюсов возбуждающих синапсов, поэтому ВПСП возникает только в области постсинаптической мембраны данного синапса, и сам по себе оказывается недостаточным для инициации ПД в области аксонного холмика нейрона.
5. Трансформация ритма возбуждений. Заключается в том, что количество нервных импульсов, поступающих к какому-либо центральному нейрону (например, по чувствительным нервным волокнам), как правило, всегда отлично от такового, возникающего в ответ в этом нейроне. Так, частота импульсации, проводимой по чувствительным нервным волокнам, как правило, отлична от той, которая возникает во вставочных и эфферентных нервных волокнах. Отмеченное обстоятельство обусловлено разной лабильностью и электрофизиологическими свойствами (например, неодинаковой продолжительностью следовых потенциалов, сопровождающих пик потенциала действия) различных нервных элементов нервного центра
6. Рефлекторное последействие. Проявляется в том, что собственно рефлекторный ответ на действие раздражителя прекращается не мгновенно после прекращения раздражения соответствующего рецептивного поля рефлекса, а спустя какое-то время, что свидетельствует в пользу некоторой инерции возбуждения в нервном центре. Такая инерция возбуждения в нервном центре может быть следствием, как минимум, двух обстоятельств. Во-первых, возбуждение вставочного или эфферентного нейрона, возникающее в результате передачи информации от чувствительного нейрона, не исчезает сразу же после прекращения возбуждения чувствительного нейрона, а может сохраняться некоторое время благодаря выраженной следовой деполяризации в нейронах нервного центра после каждого потенциала действия, вызывающей возникновение новых импульсных ответов. Во-вторых, инерция возбуждения в нервных центрах может быть обусловлена способностью возбуждения циркулировать по замкнутым нейронным цепям нервного центра.
 |
Рис. 16. Схема циркуляции возбуждения по замкнутым нервным цепям в нервных центрах (по Бабскому Е.Б, 1972).
7. Утомление нервных центров. Состоит в снижении работоспособности нервных центров после длительного периода постоянной их активности и, как следствие, временном ослаблении или полном прекращении регуляции какой-то физиологической функции. Более высокая утомляемость нервных центров, по сравнению с нервными клетками, обусловлена наличием синаптических контактов химического типа между различными нейронными элементами в пределах нервного центра.
8. Тонус нервных центров, проявляющийся в постоянном их возбуждении. Возникает и поддерживается в большинстве нервных центров по причине постоянного поступления информации по афферентным нейронам от исполнительных органов рефлекторных актов (т.е. наличию постоянной афферентации от исполнительных органов в нервный центр).
9. Избирательная повышенная чувствительность нервных центров к определенным веществам, нарушающим их нормальную работу(т.н. нервным ядам). Предполагают, что нервные яды нарушают нормальное протекание обмена веществ в нервных центрах, действуя на определенные ферментативные системы их нейронов. Так, например, рвотный центр обладает высокой чувствительностью к апоморфину: это вещество вызывает стойкое возбуждение рвотного центра и связанную с этим длительную рвоту.
10. Существование постоянной обратной связи между нервным центром и исполнительными органами.Достигается благодаря постоянному поступлению по чувствительным нервным волокнам к нервному центру информации о функциональном состоянии рабочих (исполнительных) их органов, что, с одной стороны, обеспечивает поддержание постоянного тонуса (степени возбуждения нервного центра), а с другой – очень тонкую регуляцию деятельности исполнительных органов, поскольку эфферентные нервные посылки к органам постоянно модифицируются в зависимости от изменения функционального состояния исполнительных органов в момент осуществления определенного рефлекторного акта и соответственно поступающей к нервным центрам афферентной информации о состоянии регулируемых органов.
11. Пластичность нервных центров, проявляющаяся в компенсаторной их приспособляемости к повреждению и возможности функциональной перестройки. Так, частичное повреждение нервного центра вначале сопровождается возникновением определенных расстройств в регулируемой физиологической функции, которые со временем полностью или частично проходят по причине компенсаторных изменений в оставшихся нормальными структурах нервного центра, восполняющих функцию поврежденных структур. Функциональная перестройка нервных центров возникает также в случае искусственного, вызванного хирургическим вмешательством, изменения связи нервных центров с исполнительными органами. Так, например, перерезка каких-то нервов, иннервирующих различные органы, и последующее перекрестное их сшивание, в результате чего нервы начинают иннервировать другие внутренние органы, со временем сопровождается тем, что нервные центры начинают нормально регулировать деятельность других органов, с которыми оказались связанными (т.е. нервные центры приобретают новые, ранее им не свойственные, функции). Так, например, при параличе лицевого нерва иногда производят операцию по сшиванию периферического конца этого нерва (собственно вступающего в контакт с мимической мускулатурой лица) с какими-либо другими нормальными черепно-мозговыми нервами, и через некоторое время функция неработающих по причине паралича лицевого нерва мимических мышц лица восстанавливается.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Пластичность нервных центров
Компенсаторные приспособления
Приспособляемость нервных центров и изменчивость их функционального значения А. Бете назвал пластичностью .
В очень демонстративной форме пластичность нервных центров выявляется при различных оперативных вмешательствах на нервной системе, в частности в опытах с перекрестным сшиванием нервных стволов. Два разных нервных ствола перерезают и концы их — центральный одного и периферический другого — соединяют швами.
Перерезка нервных волокон вызывает дегенерацию их периферических концов, но затем происходит регенерация: из центрального конца нерва на периферию прорастают нервные волокна, которые вновь образуют окончания в том органе, куда ходил прежде периферический конец перерезанного нерва.
При этом окончания, вновь образующиеся на периферии, специфичны не для данного нервного волокна, а для того органа, куда врастает нервный ствол, блуждающий нерв, соединенный с нервом скелетной мышцы и врастающий в нее, даёт такие же концевые пластинки, как любой моторный нерв, а при врастании в симпатический нервный ствол образует окончания, характерные для симпатического нерва.
В результате такого сшивания нервов через определенный промежуток времени (обычно через несколько месяцев) происходит коренная перестройка нервных центров, приобретающих новые, несвойственные им ранее функции.
Например, после сшивания центрального конца подъязычного нерва, иннервирующего мышцы языка, с периферическим концом дпафрагмального нерва нервные клетки ядра подъязычного нерва приобрели способность посылать ритмические разряды импульсов к диафрагме, в результате чего восстанавливаются ее дыхательные движения. Иными словами, эти нервные клетки функционально включаются в состав дыхательного центра.
П. К. Анохин наблюдал восстановление движений и чувствительности конечностей собаки при вшивании блуждающего нерва в нервные стволы передней лапы.
При наличии медицинских показаний, например при параличе лицевого нерва, производили операции сшивания нервов разного функционального значения и у человека. При этом в периферический конец парализованного лицевого нерва вшивали центральный конец одного из соседних нервов.
В результате операции наблюдалось восстановление нормальной иннервации лицевых мышц.
Восстановление (компенсация) нарушенной функции наблюдается также после разрушения или удаления отдельных участков центральной нервной системы. Так, движения, резко нарушенные у животных в первое время после полного или частичного удаления мозжечка, удаления отдельных участков головного мозга или разрушения лабиринтов, могут через некоторое время в известной степени восстановиться вследствие пластичности нервных центров. Как правило, для такого восстановления требуется много месяцев.
Весьма большое значение в явлениях компенсаторной приспособляемости нервных центров к повреждению и в функциональной перестройке их у высших животных и человека имеет кора больших полушарий головного мозга как орган условнорефлекторных связей. Э. А. Асратяном показано, что если удалить у животного кору мозговых полушарий после операции-сшивания разнородных нервов или ампутации лап, или экстирпации мозжечка в период, когда произошло приспособление к поврежден и восстановление функций движения, то это вновь вызывает расстройства движений, характерные для ранее произведенной операции.
Очевидно, кора полушарий, наиболее молодой и совершенный отдел центральной нервной системы, обладает максимально выраженной пластичностью и обеспечивает приспособление всех остальных отделов к новым после нанесенного повреждения условиям функционирования.
биологические науки
- Ланская Ольга Владимировна , доктор наук, доцент, профессор
- Великолукская государственная академия физической культуры и спорта
- КОНЦЕПЦИЯ НЕЙРОПЛАСТИЧНОСТИ
- ПЛАСТИЧНОСТЬ НЕЙРОМЫШЕЧНОЙ СИСТЕМЫ
Похожие материалы
- Изменение иммунорезистентного профиля у баскетболистов в течение годичного тренировочно-соревновательного цикла
- Психофизиологические особенности представителей различных видов спорта
- Нейропластичность как основа восстановления после инсульта и двигательная реабилитация постинсультных больных
- Уровень физического развития детей младшего школьного возраста с нарушением зрения
- Психоневрологические расстройства и качество жизни больных остеохондрозом позвоночника
Возможности проявления разнообразных адаптивных реакций и поддержания уровня оптимального функционирования организма в немалой степени определяются состоянием центральной и периферической нервной системы, активностью скелетных мышц, резервами их энергетического обеспечения (И.Б. Козловская, 1976; Б.С. Шенкман с соавт., 2010).
Пластичность – важнейшее свойство нервной системы наряду с достаточной устойчивостью ее морфологической и функциональной организации (Ю.Г. Васильев, Д.С. Берестов, 2011). Существуют различные определения пластичности нервной системы. В словаре физиологических терминов под общей редакцией академика О.Г. Газенко (1987) пластичность применительно к ЦНС интерпретируется как способность нервных элементов к перестройке функциональных свойств под влиянием длительных внешних воздействий или при очаговых повреждениях нервной ткани. Посттравматическая пластичность выполняет компенсаторную (восстановительную) функцию, а пластичность, вызванная длительным афферентным раздражением, - приспособительную. Под пластичностью также понимается изменение эффективности или направленности связей между нервными клетками (П.Г. Костюк, 1981). Благодаря пластическим перестройкам межнейронных связей возникают микро- и макроструктурные объединения, системная (взаимосвязанная) деятельность которых лежит в основе врожденных и приобретенных форм целенаправленного поведения. Отмечается, что пластичность может проявляться как на уровне отдельной клетки, так и на уровне интегративной нервной деятельности.
В свою очередь, Merriam-Webster’s Medical Dictionary определяет пластичность, как способность к продолжительной альтерации невральных путей и синапсов при жизни мозга и нервной системы в ответ на опыт или повреждение (http://medical.merriam-webster.com/) (J.R. Wolpaw, A.M.Tennissen, 2001). Вместе с тем, Е.И. Гусев, П.Р. Камчатнов (2004) в своей работе интерпретируют пластичность как способность нервной ткани изменять структурно-функциональную организацию под влиянием экзогенных и эндогенных факторов. Именно это качество обеспечивает ее адаптацию и эффективную деятельность в условиях изменяющейся внешней и внутренней среды. Структурная пластичность относится к способности нервной системы изменять свою физическую структуру под влиянием обучения или повреждения. Например, аксональный и дендритный спраутинг, генезис нервной ткани, синаптогенез и другое на фоне реабилитационных вмешательств при повреждении в спинном мозге (V.R. Edgerton et al., 2004). Современный этап развития нейробиологии характеризуется усилением внимания и к вопросу о функциональной пластичности, которая является основой памяти, обучения, формирования новых рефлексов и функциональных систем, а также способствует восстановлению функции нервной системы после повреждения (E. Kandel et al., 1991; Г.Н. Крыжановский, 2001). Так, например, правильно организованная реабилитационная терапия после спинномозгового повреждения способствует нормализации рефлексов, усилению двигательных вызванных потенциалов и в целом восстановлению чувствительных, двигательных или автономных функций (V.R. Edgerton et al., 2004).
Пластичность участвует в возникновении и закреплении как биологически полезных, так и патологических изменений, то есть по своей биологической сущности различают физиологическую (полезную) и патологическую пластичность. Физиологическая пластичность проявляется адекватным реагированием на действие раздражителей, способствует закреплению в памяти возникающих изменений, необходимых для развития нервной системы. Патологическая пластичность проявляется в том, что при включении механизмов нейропластичности, например, при повреждении мозга возникают новые, иногда ошибочные межнейрональные связи, которые отсутствовали в норме. Они нередко усугубляют имеющиеся церебральные расстройства или способствуют возникновению новых нарушений. Постепенно под влиянием патологической пластичности повышается активность деятельности патологических функциональных систем, они становятся резистентными к различным, в том числе, медикаментозным воздействиям. Патологическая пластичность способствует возникновению генераторов патологического возбуждения.
Пластичностью обладают все нервные клетки, но наиболее высокий потенциал нейропластичности имеет кора головного мозга в связи с локализацией в ней различных в функциональном отношении клеток и наличием многочисленных их связей. Так, например, рядом авторов показаны различные примеры пластичности моторной коры у развивающихся (D.D. O'Leary, N.L. Ruff, R.H. Dyck, 1994) и взрослых животных (P.M. Rossini, F. Pauri, 2000). В экспериментальных работах на человеке, несмотря на трудности в интерпретации полученных данных, пластичность моторной коры показана рядом исследователей (E.M. Bütefisch et al., 2000; Е.В. Дамянович, Т.В. Орлова, 2004). Таким образом, корковая нейрональная пластичность является одним из основных источников компенсаторных изменений в системе моторного контроля как у развивающихся и взрослых животных, так и человека (А.Б. Вольнова, 2003). В то же время существенна роль и других отделов головного мозга - таламуса, ствола, ретикулярной формации, лимбической системы, а также глиальных структур. Любые регулирующие церебральные воздействия на деятельность органов и тканей возможны благодаря нейропластичности. Она регулирует лабильность, гомеостаз, нейротрофику, обмен веществ и различные приспособительные реакции организма (A. Sarkar et al., 2011).
Изучение функциональной пластичности макромодуля двигательной системы - мышцы, степени её вовлечённости в процесс, сохранности иннервации или определение объёма реиннервации, являются по существу основными вопросами, решаемыми при проведении электронейромиографических (ЭНМГ) исследований. Пластичность - одно из важнейших свойств скелетной мышцы. Оно позволяет мышечной ткани адаптироваться к изменению условий функционирования, воздействующих как на мышцу, так и на организм в целом. Важную роль в реализации феномена мышечной пластичности занимает регуляция размеров мышечных волокон на системном и местном уровне. При этом изменения показателей размеров волокон скелетных мышц, таких как площадь поперечного сечения (например, при силовой тренировке или гравитационной разгрузке) влечет за собой существенные изменения сократительных возможностей целой мышцы (А.И. Григорьев, Б.С. Шенкман, 2008).
Подводя некоторый итог вышеизложенных сведений, можно резюмировать: исследования отечественных и зарубежных специалистов в области генетики, цитологии, биохимии, клеточной биологии, нейрофизиологии, медицины и других смежных наук свидетельствуют о том, что под влиянием внешних и внутренних воздействий осуществляется реализация механизмов функциональной пластичности (от субклеточного до системного), в том числе, в нервной и нервно-мышечной системах, которые обладают высокими приспособительными возможностями, позволяющими эффективно функционировать в различных условиях.
Далее коснемся вопроса о классических и современных взглядах на концепцию нейропластичности. Нервная система играет важнейшую роль в процессе взаимодействия между живыми организмами и средой их обитания. В основе такого взаимодействия лежит способность нервной системы приобретать, хранить и воспроизводить информацию о прошлом опыте. Нервная система рассматривается как ведущая интегрирующая система организма, обеспечивающая высшие психические функции: сознание, память, мышление и регулирующая функциональный статус органов и систем. Эти и другие возможности обеспечиваются благодаря такому свойству нервной системы как пластичность.
Обычно, в ходе рассмотрения морфо-функциональной организации ЦНС оперируют следующими уровнями (Ю.Г. Васильев, Д.С. Берестов, 2011): минимальной структурно-функциональной единицей рассматривают нейрон, группы нервных клеток, в свою очередь, кооперируются в нейронные ансамбли, совокупность которых образует нервный центр. Функция нервных центров определяется морфологической и функциональной специализацией нейронов. Важную роль играют межнейронные взаимодействия, осуществляемые с помощью специальных контактов – синапсов, и способность нервных клеток формировать различные нейронные ансамбли.
Нейроглия (глия, глиоциты), в свою очередь, представляет собой полиморфное и гетерогенное по составу семейство клеток, которые искусственно объединены по признаку вспомогательной функции по отношению к нейронам. Глиоциты, или глиальные клетки, формируют весьма сложное и крайне важное микроокружение для нейронов, без которого собственно специфическая деятельность ведущей популяции клеток нервной ткани весьма затруднительна, если вообще возможна (Ю.Г. Васильев, Д.С. Берестов, 2011). Нейроглия формирует соответствующие условия для формирования потенциала действия и его последующей передачи на значительное удаление, контролирует процессы трофического обеспечения. В ЦНС выделяют макроглию (к ней относятся различные разновидности астроцитов; олигодендроциты, которые связывают как с белым, так и с серым веществом мозга, но большее их представительство наблюдается в зоне локализации нервных волокон, по функции и положению они более близки к леммоцитам периферической нервной системы), микроглию (глиальные макрофаги, образуется из моноцитов крови) и эпендимную глию (выстилает канал спинного мозга, полости желудочков головного мозга). В периферической нервной системе выделяют шванновские клетки и сателлитную глию периферических нервных ганглиев.
Рассмотрим развитие концепции нейропластичности. В разработке основных положений нейронной теории принимали участие А.С. Догель, С. Рамон-и-Кахаль, Б.И. Лаврентьев. Согласно классическим представлениям, нейроны составляют цепи и сети, в которых осуществляется обработка информации и обеспечиваются ответы, в том числе, определяющие поведение животного и человека. Каждый нейрон, наряду с обычными для всех клеток свойствами, обладает не только способностью к переработке, но и передаче информации к другим клеткам посредством отростков и синапсов. Стоит подчеркнуть, что современные концептуальные сведения касательно информации о нервной системе привнесли много нового к имеющимся классическим представлениям в этой области. Уже в 80-е годы XX века в нейроне, наряду с единственным вариантом входа и выхода информации через химические синапсы, обнаруживаются и иные способы ее передачи – щелевидные контакты. Предложено считать элементарной пространственной единицей не отдельную клетку, а ансамбль связанных между собой нейронов. В связи с этим возрастало осознание роли не только нейронов, но и прилежащего глиального и сосудистого окружения. Современные данные позволяют расширить это представление, указывая на возможность внесинаптических взаимодействий. Такое влияние оказывается не только на низкомолекулярные органические и неорганические монометры, но и на часть полимерных образований, с включением в систему узких межклеточных пространств как путей распространения веществ и весьма значимого элемента контроля нейронной активности. Таким образом, при изучении особенностей клеточных механизмов пластичности различных областей мозга необходимо учитывать не только нейронную организацию, но и все окружение, осуществляющее поддержание гомеостаза и способное существенно изменять функцию.
Данное предположение подкрепляется несколькими положениями, выдвинутыми Ч. Шеррингтоном (1969). Им были выявлены некоторые интересные закономерности функционирования нервной системы. В частности он указывал на более значительную изменчивость пороговых величин раздражения в рефлекторных дугах по сравнению с нервными стволами. Это, согласно современным представлениям, может быть связано с динамикой в синаптической передаче или с модуляцией сигнала, обусловленной влияниями ближайшего глиального окружения, а также перераспределением ионного содержимого межклеточного вещества при длительном возбуждении как самих активируемых, так и прилежащих к ним нейронов. Другое положение, выдвинутое Ч. Шеррингтоном, указывает на большую зависимость от кровообращения и снабжения кислородом в нейронных системах по сравнению с передачей сигнала в отдельном нервном волокне. Этот факт может заключать в себе как известную зависимость нервных клеток от поступления нутриентов (в первую очередь глюкозы), так и тривиальную информацию, касающуюся, например, прямой зависимости нейрона от уровня обеспечения процессов аэробного фосфорилирования.
Следует также отметить, что с 70-80-х гг. прошлого века накопившиеся экспериментальные данные поставили под сомнение достаточность классических представлений нейронной теории С. Рамон-и-Кахаля в объяснении механизмов функционирования мозга. Концепция о нейроне как о поляризованной единице, связанной с аналогичными единицами с помощью тесно пространственно расположенных синапсов, потребовала существенного пересмотра. Так, была показана структурная и функциональная гетероморфность самих нейронов. Несколько позднее были выявлены факты о возможности пространственно удаленных межнейронных взаимодействий, которые, являясь более медленными по скорости связей и, в основном, менее интенсивными, тем не менее, способны носить весьма устойчивый характер. Данные взаимодействия, как показано в исследованиях конца XX - начала XXI в., могут опосредоваться как через межклеточное пространство, так и с помощью клеток-посредников. Наиболее интенсивно в качестве такого посредника в ЦНС позвоночных изучаются астроциты, но немаловажная роль отводится и иным клеточным структурам ЦНС и нервной периферии.
Одним из интенсивно исследуемых направлений в современной нейробиологии является проблема пластичности ЦНС при ее повреждениях. Пластичность ЦНС включает процессы различных уровней функционирования и временной протяженности, по-разному проявляющиеся при центральном и периферическом поражении нервной системы. О пластичности, как важном свойстве живого организма, много писали крупнейшие представители биологии и медицины – И.М. Сеченов, А.Н. Северцов, И.П. Павлов, Ч.С. Шеррингтон, Ф.Л. Гольц, А. Бете, Р. Магнус, Л.А. Орбели, В.М. Бехтерев, Э.А. Асратян, П.К. Анохин и др. В течение многих лет накоплен разнообразный материал о процессах восстановления функций после различного рода повреждений, но общей теории пластичности до тридцатых годов прошлого столетия создано не было.
В нейрофизиологии значительные шаги в этом направлении были сделаны А. Бете в 30-ых годах прошлого столетия. Исходя из своих экспериментальных данных и клинических наблюдений, а также данных некоторых других исследователей, он предложил общую теорию пластичности, в соответствии с которой решающая роль в компенсации функций принадлежит не ЦНС, а периферическим импульсам и периферическим рецепторам. Понимая принципиальную ошибочность теории Бете, Э.А. Асратян, основываясь на своих собственных данных, а также достижениях других представителей передовой мировой нейрофизиологии, выдвинул концепцию, согласно которой у высших животных в процессе восстановления функций, нарушенных после повреждения различных отделов ЦНС и периферической нервной системы, решающая роль принадлежит коре больших полушарий головного мозга, и что пластичность является одним из важных свойств этого отдела нервной системы.
На основании проведенных к настоящему времени исследований предполагалось, что возможные механизмы пластичности поврежденной ЦНС можно разделить на две группы. К первой группе относятся: функциональная перестройка, преобразование запасных путей, повышение активности синапсов и вовлечение нервных связей, которые в норме не функционируют. Этот механизм чисто функциональный и в нем решающая роль отводится коре больших полушарий. Он основан на ее условнорефлекторной деятельности и таких явлениях как рекрутирование, вовлечение новых единиц, облегчение, усиление и т.д. Ко второй группе относятся: органические, структурные или вегетативные изменения, в число которых входят регенерация, гипертрофия нервных клеток, образование новых коллатералей от интактных аксонов, которые дают аксосоматические или аксодендритические контакты с частично деафферентированными нейронами, так называемое явление аксонального спраутинга и т.д., которые связаны с обменом веществ и обусловлены химическими механизмами. И та, и другая точки зрения не противоречат современной концепции нейропластичности, в частности, положению о том, что при поражении мозга возникающие симптомы нейродефицита отражают не проявления поврежденной области мозга, а функцию всего мозга, точнее, пластические изменения в оставшейся функционирующей части мозга, в числе которых могут быть и те, которые блокируют восстановление функции.
Изменения, выявляющие пластичность мозга, происходят на молекулярном, клеточном, синаптическом и анатомическом (охватывая значительные группы нейронов – нейронные ансамбли) уровнях. При этом могут быть вовлечены не только корковые отделы, но и подкорковые структуры. Кроме того, помимо структурных изменений отмечаются и динамические, последовательно сменяющие друг друга сдвиги функционального характера, как в окружающей очаг повреждения зоне, так и на расстоянии от этого очага. Процессы, связанные с нейропластичностью, могут носить как системный, так и локальный характер.
Изложенные сведения указывают, что в соответствии с современными представлениями нейрон, являясь ведущим исполнителем основных функций нервной системы, не является независимым элементом. Он весьма подвержен влиянию как клеток этой же популяции, так и прилежащего окружения. В то же время нейроны весьма разнообразны как по структурной, так и функциональной организации. Через описание и даже подробнейшее рассмотрение отдельного нейрона невозможно описать функцию всей системы в целом. Значима роль не только отдельного нейрона, но и взаимодействующей системы из нейронных ансамблей, неоднородных по качественной и количественной природе. Определенный интерес в этом отношении вызывает специализированная система межнейронных коммуникаций в виде синаптических контактов, что и будет рассмотрено в следующих работах.
Список литературы
Электронное периодическое издание зарегистрировано в Федеральной службе по надзору в сфере связи, информационных технологий и массовых коммуникаций (Роскомнадзор), свидетельство о регистрации СМИ — ЭЛ № ФС77-41429 от 23.07.2010 г.
Соучредители СМИ: Долганов А.А., Майоров Е.В.
Читайте также:
