
Лекции по физиологии мышц и нервов
В этой книге предельно сжато изложен курс лекций по нормальной физиологии. Благодаря четким определениям основных понятий студент может сформулировать ответ, за короткий срок усвоить и переработать важную часть информации, успешно сдать экзамен. Курс лекций будет полезен не только студентам, но и преподавателям.
- ЛЕКЦИЯ № 1. Введение в нормальную физиологию
- ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
- ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
- ЛЕКЦИЯ № 4. Физиология мышц
- ЛЕКЦИЯ № 5. Физиология синапсов
- ЛЕКЦИЯ № 6. Физиология центральной нервной системы
- ЛЕКЦИЯ № 7. Физиология различных разделов ЦНС
Приведённый ознакомительный фрагмент книги Нормальная физиология: конспект лекций (С. С. Фирсова) предоставлен нашим книжным партнёром — компанией ЛитРес.
ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
1. Физиология нервов и нервных волокон. Типы нервных волокон
Физиологические свойства нервных волокон:
1) возбудимость – способность приходить в состояние возбуждения в ответ на раздражение;
2) проводимость – способность передавать нервные возбуждение в виде потенциала действия от места раздражения по всей длине;
3) рефрактерность (устойчивость) – свойство временно резко снижать возбудимость в процессе возбуждения.
Нервная ткань имеет самый короткий рефрактерный период. Значение рефрактерности – предохранять ткань от перевозбуждения, осуществляет ответную реакцию на биологически значимый раздражитель;
4) лабильность – способность реагировать на раздражение с определенной скоростью. Лабильность характеризуется максимальным числом импульсов возбуждения за определенный период времени (1 с) в точном соответствии с ритмом наносимых раздражений.
Нервные волокна не являются самостоятельными структурными элементами нервной ткани, они представляют собой комплексное образование, включающее следующие элементы:
1) отростки нервных клеток – осевые цилиндры;
2) глиальные клетки;
3) соединительнотканную (базальную) пластинку.
Главная функция нервных волокон – проведение нервных импульсов. Отростки нервных клеток проводят сами нервные импульсы, а глиальные клетки способствуют этому проведению. По особенностям строения и функциям нервные волокна подразделяются на два вида: безмиелиновые и миелиновые.
Безмиелиновые нервные волокна не имеют миелиновой оболочки. Их диаметр 5–7 мкм, скорость проведения импульса 1–2 м/с. Миелиновые волокна состоят из осевого цилиндра, покрытого миелиновой оболочкой, образованной шванновскими клетками. Осевой цилиндр имеет мембрану и оксоплазму. Миелиновая оболочка состоит на 80 % из липидов, обладающих высоким омическим сопротивлением, и на 20 % из белка. Миелиновая оболочка не покрывает сплошь осевой цилиндр, а прерывается и оставляет открытыми участки осевого цилиндра, которые называются узловыми перехватами (перехваты Ранвье). Длина участков между перехватами различна и зависит от толщины нервного волокна: чем оно толще, тем длиннее расстояние между перехватами. При диаметре 12–20 мкм скорость проведения возбуждения составляет 70—120 м/с.
В зависимости от скорости проведения возбуждения нервные волокна делятся на три типа: А, В, С.
Наибольшей скорость проведения возбуждения обладают волокна типа А, скорость проведения возбуждения которых достигает 120 м/с, В имеет скорость от 3 до 14 м/с, С – от 0,5 до 2 м/с.
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Существует три закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Закон изолированного проведения возбуждения.
Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах.
В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно.
В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
- ЛЕКЦИЯ № 1. Введение в нормальную физиологию
- ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
- ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
- ЛЕКЦИЯ № 4. Физиология мышц
- ЛЕКЦИЯ № 5. Физиология синапсов
- ЛЕКЦИЯ № 6. Физиология центральной нервной системы
- ЛЕКЦИЯ № 7. Физиология различных разделов ЦНС
Приведённый ознакомительный фрагмент книги Нормальная физиология: конспект лекций (С. С. Фирсова) предоставлен нашим книжным партнёром — компанией ЛитРес.
Согласно современным представлениям, нервная и мышечная ткани могут находиться в трех основных состояниях — физиологическом покое, возбуждении и торможении.
Физиологический покой — это такое состояние, когда ткань или орган не проявляет признаков присущей им деятельности.
Возбуждение — деятельное состояние живой ткани, в которое она приходит под влиянием раздражения.
Торможение — такое состояние, когда деятельность ткани или органа ослабляется или полностью прекращается.
Раздражители.При раздражении мышца приходит в состояние возбуждения, которое распространяется по мышце и вызывает ее сокращение. По биологическому действию раздражители бывают адекватные и неадекватные. Адекватный — это такой раздражитель, к которому данный орган или ткань приспособились в процессе эволюции. Для мышцы адекватным раздражителем является нервный импульс. Неадекватными — будут такие раздражители, действию которых ткань или орган в естественных условиях обычно не подвергаются. Так, сокращение мышцы можно вызвать механическим раздражением (укол, кислота, ток) и т.д.
Все раздражители по своей силе деляг на пороговые, подпороговые и сверхпороговые. Пороговыми называют минимальные раздражители, которые могут вызвать возбуждение. Подпороговые — это раздражители, сила которых меньше пороговой, сверхпороговые — раздражители более сильные, чем пороговые.
Биоэлектрические явления.Возникновение и распространение возбуждения связано с изменением электрического заряда на поверхности клеточной мембраны и внутри клетки (рис. 4.19).
Потенциал покоя.Мембрана нервной или мышечной клетки в состоянии покоя с наружной стороны заряжена положительно, а с внутренней — отрицательно. Разность зарядов между наружной и внутренней поверхностью клеточной мембраны в состоянии физиологического покоя клетки называется потенциалом покоя, или мембранным потенциалом.
Потенциал действия. Под влиянием раздражения проницаемость клеточной мембраны для ионов натрия повышается в сотни
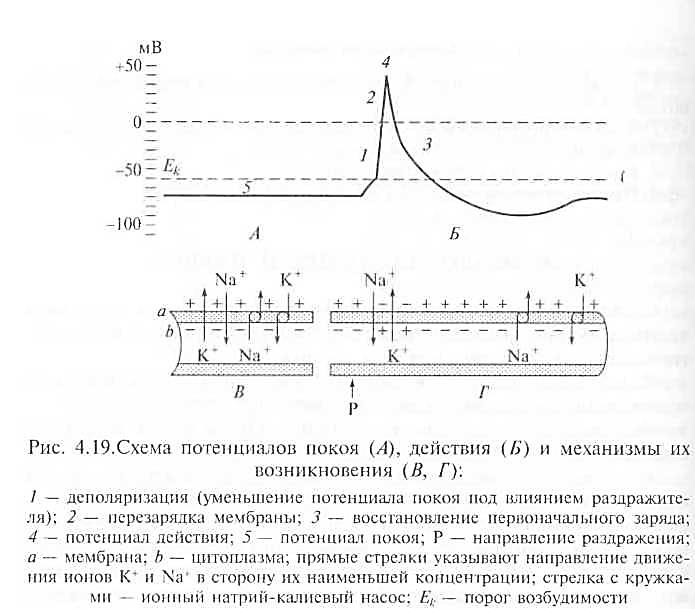 |
Рис. 4.19.Схема потенциалов покоя (А), действия (5) и механизмы их
возникновения (В, Г):
1 — деполяризация (уменьшение потенциала покоя под влиянием раздражите-
ля); 2 — перезарядка мембраны; 3 — восстановление первоначального заряда;
4 — потенциал действия; 5 — потенциал покоя; Р — направление раздражения;
а — мембрана; b — цитоплазма; прямые стрелки указывают направление движе-
ния ионов К + и в сторону их наименьшей концентрации; стрелка с кружка-
ми — ионный натрий-калиевый насос; Ек — порог возбудимости
раз. Происходит перезарядка клеточной мембраны: внутренняя ее сторона заряжается положительно, а наружная — отрицательно. Затем значительно повышается проницаемость мембраны для ионов калия, они начинают быстро перемещаться из клеток во внеклеточную жидкость и вновь восстанавливают первоначальный заряд мембраны. Пикообразное колебание потенциала, возникающее в результате кратковременной перезарядки мембраны и последующего восстановления ее заряда, называется потенциалом действия. Такие колебания потенциала продолжаются 1 — 5 мс.
Проведение возбуждения. Круговой ток, проходя через участок покоя, раздражает его и вызывает появление потенциала действия (рис. 4.20). Когда в одном из участков нервной клетки, нервного или мышечного волокна возникает возбуждение и появляется потенциал действия, между возбужденным и соседним невозбужденным участком возникает разность потенциалов. Эта разность потенциалов является причиной появления круговых (локальных) токов. Внутри клетки ток идет от возбужденного участка к участку покоя, по внешней стороне — от участка покоя к возбужденному.
В участке, который ранее был возбужден, происходит восста
Рис. 4.20. Схема распрост-
ранения возбуждения:
А — волокна мышц и безмякотных нервных волокон; Б —
мякотное нервное волокно; 1 — мембрана; 2 — волокно;
а, Ь, с — перехваты Ранвье; заштрихованы возбужденные
участки; светлые — участки покоя; стрелками показано движение круговых токов

новление потенциала покоя. Данный процесс многократно повторяется и обусловливает распространение импульсов возбуждения. Скорость проведения возбуждения по скелетным мышцам 12 —15 м/с, гладким мышцам — 1 — 18 см/с, в безмякотных нервах — 0,5 — 3 м/с.
Основные свойства живой ткани.Любая живая клетка обладает свойствами раздражимости, возбудимости и лабильности (функциональной подвижности).
Раздражимость. С раздражимостью связаны все процессы роста и размножения клеток и тканей. На разнообразные воздействия внешней или внутренней среды — раздражения протоплазма живой клетки отвечает специфической реакцией, при которой происходят усиление или ослабление обмена веществ, количественное и качественное его изменение. Эта реакция на раздражение получила название раздражимости, или реактивности.
Возбудимость — свойство нервной или мышечной ткани отвечать на действие раздражителя возбуждением. Для измерения величины возбудимости определяют порог возбудимости и хронаксию. Порогом возбудимости называется наименьшая сила раздражителя, способная вызвать возбуждение; его выражают в вольтах постоянного тока.
Лабильность — скорость, с которой в ткани возникает и успевает закончиться полный период отдельного импульса возбуждения. Мера лабильности — это максимальное число импульсов возбуждения за 1 св ответ на такое же максимальное число раздражений. Высокой лабильностью обладают мякотные нерпы, у без- мякотных нервов она низкая. Так, максимальный ритм возбуждений мякотного нервного волокна 500, безммкотного — 200. Максимальный ритм возбуждений скелетной мышцы 200 импульсов в 1 с, а гладкой мышцы — в десятки раз меньше.
При очень частых раздражениях сокращения мышцы уменьшаются или даже совсем прекращаются. Такая частота называется пессимальной, или пессимумом. Пессимум возникает вследствие того, что возбуждение еще не закончилось и ткань находится в состоянии абсолютной или относительной рефрактерное™, а на нее действует новое раздражение. Частые раздражения, превышающие меру лабильности, вызывают не возбуждение, а торможение.
Учение Н. Е. Введенского о парабиозе устанавливает связь между возбуждением и торможением. Возбуждение и торможение — это различные реакции ткани на раздражение, исход которогозависит от лабильности. При высокой лабильности возникает возбуждение, снижение лабильности вызывает торможение.
ФИЗИОЛОГИЯ МЫШЦ
Скелетные мышц, входящие в состав опорно-двигательного аппарата позвоночных животных, состоят из группы мышечных пучков, каждый из них составлен из тысяч мышечных волокон, которые представляют собой клетки цилиндрической формы длиной до 12 см и диаметром 10 — 100 мкм. Каждое волокно окружено оболочкой сарколеммой и содержит тонкие нити — миофибрил-лы (рис. 4.21). Поперечные мембраны делят каждую миофибриллу на отдельные участки — саркомеры. Сократимым веществом мышечного волокна являются миофибриллы, состоящие из множества (около 2500) тонких и толстых белковых нитей — протофибрилл. Толстые протофибриллы образованы из белка миозина, тонкие — из актина. Нити актина прикреплены к мембране саркомера; они образуют светлые участки миофибриллы. В темных участках находятся нити миозина. Акгиновые нити частично входят своими концами в промежутки между миозиновыми нитями. Нити актина и миозина соединяются между собой многочисленными поперечными мостиками, которые образованы скрученными в спираль отростками — мостиками миозиновой нити. Чередование нитей в миофибрилле обусловливает ее поперечную исчерченность.

Рис. 4.21. Схема чередова-
ния нитей в миофибрилле:
А — электронно-микроскопическая фотография мио-
фибриллы; Б — строение миофибриллы; Z — поперечная
мембрана, разделяющая миофибриллу на отдельные уча-
стки — саркомеры; 1 — миозиновая нить, 2 — актиновая нить
Гладкие мышцы, образующие стенки внутренних органов, состоят из клеток различной формы длиной от 30 до 500 мкм и диаметром 2—10 мкм; у клеток имеются боковые отростки, благодаря которым клетки гладких мышц группируются в длинные пучки. Они, в свою очередь, при помощи тяжей соединяются друг с другом, обеспечивая деятельность мышцы как единой системы. Сократительным аппаратом гладких мышц являются миофибриллы, в которых содержатся тонкие актиновьге нити с прикрепленными к ним короткими отростками миозиновых нитей, называемых димерами.
Свойства скелетных мышц.Проведение возбуждения в мышцах происходит изолированно, т. е. потенциалы действия не переходят с одного мышечного волокна на другое. Возбудимость скелетныхмышц меньше возбудимости нервов. Нервное волокно оканчивается в середине мышечного волокна, поэтому возбуждение распространяется в обе стороны со скоростью 4—15 м/с.
Скелетная мышца является упругим телом. Если к мышце подвесить груз, то она растягивается, это свойство называется растяжимостью. Эластичностью мышцы называется возвращение мышцы к первоначальной своей длине после растяжения, когда будет удален груз. Пластичностью мышцы называется свойство сохранять удлиненную форму после удаления груза, вызвавшего ее растяжение. В скелетных мышцах лучше выражено свойство эластичности, чем пластичности.
Виды мышечных сокращений.При нанесении на мышцу одиночного раздражения она отвечает одиночным сокращением. Записывая это сокращение на двигающейся ленте какого-либо самописца (например, в простейшем случае на кимограф), можно отметить три периода: латентный — период от раздражения до начала сокращения, период сокращения и период расслабления. У икроножной мышцы лягушки одиночное сокращение длится 0,1 с, у млекопитающих — 0,04 — 0,1 с. В латентный период в мышце происходят процессы, при которых освобождается энергия для мышечного сокращения.
Если к мышце поступает несколько частых импульсов возбуждения, наступает длительное сокращение мышцы, которое называется тетаническим, или тетанусом. В зависимости от частоты возбуждений тетанус будет зубчатым или гладким (рис. 4.22).
Зубчатый тетанус наблюдается при такой частоте импульсов возбуждений, когда каждый импульс действует на мышцу в тот момент, когда она уже начинает расслабляться. Если же импульсы возбуждения настолько частые, что они воздействуют на мышцу до начала ее расслабления, то получатся длительные непрерывные сокращения мышцы — гладкий тетанус.
 |
Рис. 4.22. Сокращение скелетной мышцы при различной частоте раздра-
жения:
А — одиночное сокращение в ответ на одно раздражение; Б — зубчатый тетанус
(до 15 раздражений в 1 с); В — гладкий тетанус (свыше 25 раздражений); М —
механограмма — запись сокращения мышц; Р — частота раздражений; 1 — фаза
сокращения; 2 — фаза расслабления
Механизм мышечного сокращения (теория скольжения).В состоянии покоя тонкие нити актина лежат частично своими концами в промежутках между толстыми миозиновыми нитями. При возбуждении мышцы под влиянием потенциала действия нити актина сдвигаются в промежутке между нитями миозина, т. е. скользят навстречу друг другу, вызывая сокращение миофибрилл (рис. 4.23). Считают, что скольжение происходит в результате укорочения мостиков — боковых отростков миозиновых нитей под влиянием ионов кальция. Укорачиваясь, мостики тянут актиновую нить навстречу миозиновой.
Работа и сила мышц.При сокращении мышца укорачивается и тем самым совершает работу. Она будет наибольшей при средних нагрузках и среднем ритме сокращений.
Силу мышцы определяют по максимальному сокращению ее при поднятии максимального груза. Наибольшей силой обладают мышцы с косыми волокнами, имеющими перистое строение.
Утомление мышц.В процессе сокращений мышцы утомляются, при этом понижаются их возбудимость, лабильность и величина сокращения. В утомленной мышце понижается содержание гликогена и накапливаются продукты обмена. В опытах на нервно-мышечном препарате Н. Е. Введенский установил, что прежде всего утомляются синапсы в связи с их низкой лабильностью. В целом организме утомление в первую очередь наступает в нервных центрах коры больших полушарий. И. М.Сеченов доказал, что быстрое восстановление работоспособности утомленных мышц наступает не при полном их покое, а при работе других, до этого не сокращавшихся мышц. Импульсы от вновь вовлеченных в работу мышц повышают возбудимость нервных центров, возбуждение одних нервных центров снижает и даже снимает утомление других центров. Утомление зависит от состояния симпатической нервной системы и желез внутренней секреции: утомленная мышца вновь
Рис. 4.23. Схема расслабленной (А) и сократившейся (Б) миофибриллы
 |
начинает сокращаться при раздражении симпатического нерва или введении адреналина, усиливающих обмен. Тренировка мышц позволяет отсрочить наступление утомления. При ней увеличивается объем мышц в результате утолщения мышечных волокон, повышается содержание гликогена, АТФ и креатинфосфата, ускоряются восстановительные процессы.
Тонус мышц. Тонус скелетных мышц играет важную роль для поддержания определенного положения тела в пространстве и деятельности двигательного аппарата. Скелетные мышцы в покое расслабляются не полностью, а находятся в некотором напряжении, т.е. тонусе. Он обусловлен поступлением к мышцам редких нервных импульсов, возбуждающих мышечные волокна не одновременно, а поочередно.
Гладкие мышцы обладают способностью длительно находиться в тонусе. Длительные тонические сокращения этих мышц особенно отчетливо выражены в сфинктерах полых органов, в стенках кровеносных сосудов.
Разделы сайта
Новости
Нервно-мышечная физиология
- Физиология нервов
- Распространение ПД по безмякотному волокну
- Типы нервных волокон
- Законы проведения возбуждения по нервам
- I закон
- II закон
- III закон
- Нервно-мышечная передача (синапс)
- Механизм нервно-мышечной передачи
- Физиология мышц
- Скелетная мышца
- Сердечная мышца
- Глакая мышца
- Сила мышц
- Сила сокращения
- Правило средних нагрузок
- Утомление мышц, отдых
- Список использованной литературы
Физиология нервов.
Центральная нервная система у человека состоит из нервных клеток, каждая из которых имеет один аксон и много дендритов. Нервные волокна делятся на: мякотные и безмякотные. Все они имеют шванновскую оболочку, а мякотные, кроме этого, покрыты еще миелиновой оболочкой между которой имеются перехваты Ранвье, в которых миелиновая оболочка отсутствует. Безмякотные волокна имееют малый диаметр, меньше 1,3 мкм, небольшую скорость распространения импульса до 2 м/сек, продолжительность ПД – 2 мсек. Мякотные возникли из безмякотных, диаметр до 25 мкм, скорость распространения возбуждения до 120 м/сек, продолжительность ПД – 0,4-0,5 мсек.

Рисунок 1. Проведение возбуждения по миелиновому нервному волокну.
При этом может охватывать не только один, но и два перехвата Ранвье, что обеспечивает надежность, а также увеличивает скорость распространения возбуждения и экономически более выгодна, так как на 1 импульс в безмякотном волокне энергия расходуется в 20 раз больше, чем в мякотном. Таким образом, скорость и расстояние, на которое распространяется возбуждение в безмякотных волокнах пропорциональна диаметру, сопротивлению мембраны и амплитуде ПД; в мякотных прямопропорциональна длине межперехватных участков, а их длина тем больше, чем больше диаметр волокна. Скорость не зависит от силы раздражения.
Теория Германа была экспериментально подтверждена.
Гельмгольц – определил скорость распространения импульса у лягушки; Бабский – определил скорость распространения импульса у человека. Эрландер и Гассер классифицировали различные нервные волокна на три группы А, В и С:
Типы нервных волокон
А – миелинизированные, наибольшего диаметра, скорость 120-70 м в сек, длительность ПД – минимальная, делятся на подгруппы: альфа, бета, гамма, дельта. Пример – аксоны мотонейронов.
В – миелинизированные волокна, меньшего диаметра, скорость 3-18 м в сек, ПД более длителен. Пример – преганглионарные волокна симпатической нервной системы.
С – немиелизированные нервные волокна, скорость менее 2 м в сек, длительность ПД наибольшая. Пример - постганглионарные волокна парасимпатической нервной системы.
Законы проведение возбуждения по нервам.
I закон анатомо-физиологический целостности нервного волокна. Чтобы возбуждение распространялось по нервному волокну необходимо не только его морфологическая целостность, но и физиологическая непрерывность. Препараты для проводниковой анестезии нарушают физиологическую непрерывность тем, что инактивируют натриевую проницаемость в нервных волокнах.
II закон изолированного проведения возбуждения по нервному волокну. В смешанном нерве возбуждение с одного нервного волокна не передается на соседние, так как сопротивление межклеточной жидкости меньше чем сопротивление мембран соседних волокон. Этим обеспечив ается точность проведения информации в нервных волокнах к иннервируемым структурам.
III закон двухстороннее проведение возбуждения. Распространение ПД по мембране нервного волокна возможно в обе стороны, так как строение мембраны на всем протяжении одинаково. В то же время возбуждение не может возвратиться в участок, где оно возникло, так как он находится в состоянии рефрактерности.
Парабиоз. Н.Е. Введенский, исследуя прохождение импульса через отрезок нерва на который воздействую химические или наркотические вещества (альтераторы), наблюдал резкое снижение лабильности. Парабиоз характеризуется постепенным развитием , в котором можно выделить четыре фазы:
I Продромальная (не всегда проявляется, так как очень кратковременная) характеризуется: повышением возбудимости, повышением лабильности.
II Уравнительная – эффекты от сильных и слабых раздражителей уравновешиваются.
III Парадоксальная - на сильные, либо частые раздражения эффект бывает меньше, чем на слабые или редкие.
IV Тормозная – ни сильные, ни слабые раздражения не вызывают сокращения мышц. Через поражённый участок не проходят импульсы.
Если второй парой электродов подействовать на поражённый участок, то возбуждение будет, т.е. ткань ещё жива.
Если снять альтератор, то ткань возвращается к исходному состоянию в обратном порядке фаз IV, III, II, I.
Парабиоз – это стойкое не распространяющееся возбуждение.
Возникают потенциалы меньшие по своей амплитуде, а дальше абортивные потенциалы, не способные распространяться: уменьшаются процессы Na-евой проницаемости, и увеличиваются процессы Na-евой инактивации.
1) работу Са-АТФ-азы, (активация насоса происходит за счет неорганического фосфата, образующегося при гидролизе АТФ),
2) за счет Э происходит откачивание против градиента концентрации Са2+ назад в саркоплазматический ретикулум (активный транспорт).
3) АТФ в мышечной клетке обеспечивает работу Na+-К-АТФ-азы, обеспечивающей удаление Na+ из клетки и восстановление потенциала покоя (а, следовательно, и возбудимости) мышечной клетки.
АТФ обеспечивает процессы, как сокращения, так и расслабления. Если Са2+ не будет транспортироваться назад в СПР, то расслабление не наступит, развивается ригидность мышцы (при трупном окоченении), или в живом организме – при посттетанической контрактуре – накопление Са2+ в саркоплазме инициирует длительное мышечное сокращение вне поступления ПД через синапс на мембрану мышечного волокна.
Гладкая мышца содержит также тропомиозин, но не имеет тропонина, соотношение актина к миозину 14-16 к 1, сравните в скелетных соотношение актина к миозину 2 к 1. Гладкая мышца имеет щелевые контакты – это мостики, соединяющие мембраны соседних клеток. Регуляция сократительной активности гладких мышц происходит благодаря связыванию Са2+ с кальмодулином, активирующим киназу легкой цепи миозина, которая приводит к гидролизу АТФ и запускает цикл образования поперечных мостиков.
ПД скелетной мышцы длится около 2-4- мс и проходит по мембране мышечного волокна со скоростью около 5 м/с. 1 ПД вызывает одиночное мышечное сокращение, которое начинается через 2 мс после начала деполяризации мембраны (латентный период) и завершается сокращение почти одновременно с реполяризацией. Длительность одиночного сокращения различна и зависит от типа мышечной ткани. При частых стимулах развивается суммарное мышечное сокращение всех мышечных волокон, обладающих различным сопротивлением мембран к электрическому току. Однако, незначительные отличия порогов возбуждения обеспечивают синхронность мышечного сокращения целой мышцы. Наличие абсолютного рефрактерного периода около 1-3 мс, обуславливает возникновение различных видов тетанусов (суммарных мышечных сокращений). Существует зубчатый и гладкий тетанусы. Частота стимуляции мышцы для развития гладкого тетануса должна быть выше, чем для развития зубчатого. Стимул должен попадать в фазу укорочения мышцы, если же мышцы начала расслабляться, а мы ее стимулирует, то получаем – зубчатый тетанус. Минимальный промежуток времени между последовательными эффективными стимулами во время тетануса не может быть меньше рефрактерного периода, которые приблизительно соответствует длительности ПД. Поскольку мышцы состоят из мышечных волокон с различным уровнем возбудимости, имеется определенная зависимость между величиной стимула и ответной реакцией. Увеличение силы сокращения возможно до определенного предела, после которого амплитуда сокращения остается неизменной при увеличении амплитуды стимула (надо отметить, то в мышце суммируются не ПД, а сокращения). При этом все волокна, входящие в состав мышцы принимают участие в сокращении.
В организме человека имеются быстрые, фазные мышечные волокна (белые), длительность сокращения которых до 7,5 мс, и медленные, тонические (красные), которые обеспечивают сильные и мощные движения, длящиеся до 100 мс. Красные (тонические) имеют много волокон миозина типа I, которые отличаются низкой активностью АТФ-азы миозина. Скорость расщепления АТФ является фактором, предопределяющим частоту гребковых движений, и таким образом, скорость скольжения нитей актина вдоль миозина. Из СПР Са2+ транспортируется медленно, высокая окислительная способность, много капилляров, много миоглобина в структуре миозина (тяжелые цепи), много митохондрий. На стимул реагируют медленно, имеют длительный латентный период сокращения, поэтому способны к длительным, медленным, тоническим сокращениям, более резистентны к утомлению. Главная функция – поддержание положения тела. Белые мышцы содержат волокна миозина II типа. Гликолитический тип окисления, мало миоглобина, митохондрий, это волокна большого диаметра с высокой активность АТФ-азы миозина, способны развить значительную силу, но быстро утомляются.
Сила мышц определяется тем максимальным грузом, который мышца в состоянии чуть-чуть приподнять. Сила различных мышц неодинакова. Для сравнения силы разных мышц максимальный груз, который мышца в состоянии поднять делят на число квадратных сантиметров ее физиологического поперечного сечения. Силовые характеристики выше у мышц с перистым (косым) расположением волокон, при этом физиологическое сечение больше геометрического поперечного сечения. Сумма поперечного сечения не всегда совпадает с физиологическим сечением мышцы (только при параллельном расположении волокон.
Читайте также:
