
Механизм проведения возбуждения по мякотному и безмякотному нервным волокнам
В безмякотных:
ПД беспрепятственно распространяется по всей мембране, в точке раздражения возникает ПД вокруг которого создаётся электрическое поле, приводящее к деполяризации соседнего участка и открытию потенциалуправляемых натриевых каналов. Деполяризация достигает уровня КУД, возникает новый ПД, который распространяется на соседний участок.
В мякотных:
В зоне перехвата Ранвье - высокая плотность каналов Na. Если 1 перехват возбуждается, то электрическое поле вызывает деполяризацию соседнего перехвата и открытие Na-евых каналов и при достижении уровня КУД –генерализацию ПД. Возбуждение распространяется скачкообразно, что обеспечивает высокую скорость.
19.Как и почему скорость распространения возбуждения по нервным волокнам зависит от их диаметра?
Чем диаметр больше, тем сопротивление аксоплазмы меньше, скорость проведения возбуждения больше.Это связано с меньшим внутренним сопротивлением волокна.
20.Как зависит скорость распространения возбуждения по нервным волокнам от амплитуды потенциала действия? Гарантийный фактор.
Связь между амплитудой ПД и величиной возбудимости ( U) характеризует гарантийный фактор(фактор надёжности)
21.Назовите и охарактеризуйте законы проведения возбуждения в нервах. Как доказать двустороннее проведение возбуждения по нервам?
1)закон двустороннего проведения возбуждения:
Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от места его возникновения, т.е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения.
2)закон изолированного проведения возбуждения по нервным волокнам:
Возбуждение не распространяется с волокна на волокно в пределах целого нерва.
3)закон анатомической и физиологической целостности нервного волокна:
Если волокно повреждено или находится под анестезией, то проведение возбуждения нарушается.
22. Синапсы: строение, классификация, общие свойства.
Нервно-мышечный синапс-структура,обеспечивающая проведение возбуждение с нерва на мышцу.
В пределах ЦНСобразуются синапсы:
•По способу передачи информацииразличают:
•В зависимость от влияния освобожденных медиаторов на ионную проницаемостьпостсинаптической мембраны выделяют:
•В зависимости от выделяемого медиатора синапсы могут классифицироваться так:
· серотониненергические и т.д.
Строение:
Это нервное окончание,покрытое:1)пресинаптической мембраной.В её составе-множество пузырьков медиатора. Медиатор-ацетилхолин.
2)синаптическая щель-пространство между пресинаптической мембраной и мембраной мышечного волокна. Здесь находится фермент – ацетилхолинэстераза, разрушающий ацетилхолин.
3)постсинаптическая мембрана- покрывает участок мышцы, контактирующий с нервом.
Общие свойства:
1)Синапс является полярной структурой, то есть проводит возбуждение только в сторону с про- на постсинаптическую мембрану.
2)синаптическая задержка-проведение ПД через синапс составляет примерно 2,3 мсек. Время,необходимое для выделения медиатора,его миграции и взаимодействия с постсинаптической мембраной.
23.Охарактеризуйте процесс передачи возбуждения (электросекреторного сопряжения) в нервно-мышечном синапсе. Что такое холинорецептор?
1)ПД распространяется на пресинаптической мембране;
2)Её деполяризация приводит к раскрытию Са-евых каналов,которые поступают внутрь пресинапса;
3)Са вызывает движение пузырьковс медиатором, которые сливаются с мембраной, и ацетилхолин поступает в синаптическую щель;
4)Ацетилхолин начинает взаимодействовать с рецепторами постсинаптической мембраны, которые чувствительны к ацетилхолину и носят название холинрецепторы;
5)При взаимодействии ацетилхолина с холинрецепторами открываются хемовозбудимые каналы на постсинаптической мембране, через которые начинают поступать ионы Na,Cl,K, развивается деполяризация постсинаптической мембраны.
6)Поскольку постсинаптическая мембрана не способна генерировать ПД, деполяризация распространяется на внесинаптическую мембрану, где есть электровозбудимые Na- евые каналы, достигает уровня КУД,и возникает ПД, распространяется по мышечной мембране.Ацетилхолин разрушается ацетилхолинэстеразой, и синаптический процесс прекращается до принятия очередной порции медиатора.
24.Чем отличается постсинаптическая мембрана от внесинаптической мембраны?
1) Постсинаптическая мембрана является хемовозбудимой и не чувствительна к электрическому току, внесинаптическая мембрана электровозбудима;
2) постсинаптической мембраны (имеет сенсор медиатора), а внесинаптическая мембрана (имеется сенсор напряжения);
3)Ионные каналы постсинаптической мембраны не специализированны, а внесинаптической мембраны- специализированны;
4) Постсинаптическая мембрана не способна генерировать ПД даже при выраженной деполяризации. ПД генерирует внесинаптическая мембрана.
Причина – деполяризация мембраны (возникновение ПД).
Заряжены только оголенные участки, возникает разность потенциалов, кольцевой ток, перезарядка, разность потенциалов, ток, ПД вызывает ПД у другого участка. Скорость 100-120 м/с.
По миелинизированному волокну потенциал действия распространяется скачкообразно (сальтаторное проведение). Для миелинизированных волокон характерна концентрация потенциалзависимых ионных каналов только в областях перехватов Ранвье; здесь их плотность в 100 раз больше, чем в мембранах безмиелиновых волокон. В области миелиновых муфт потенциалзависимых каналов почти нет. Потенциал действия, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до критического уровня, что приводит к возникновению в них новых потенциал действия, то есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает 2-й, 3-ий, 4-й и даже 5-й, поскольку электроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля.
13. Механизм проведения по безмякотным нервным волокнам.
Из-за действия раздражителя происходит смена заряда, возникает разность потенциалов, местный ток (кольцевой), происходит перезарядка, возникает разность потенциалов, местный ток, ПД вызывает ПД у другого участка. Скорость 5-15 м/с.
По немиелинизированному волокну потенциал действия распространяется непрерывно. Проведение нервного импульса начинается с распространением электрического поля. Возникший потенциал действия за счет электрического поля способен деполяризовать мембрану соседнего участка до критического уровня, в результате чего на соседнем участке генерируются новые потенциалы. Сам потенциал действия не перемещается, он исчезает там же, где возник. Главную роль в возникновении нового потенциал действия играет предыдущий.
Если внутриклеточным электродом раздражать аксон посередине, то потенциал действия будет распространяться в обоих направлениях. Обычно же потенциал действия распространяется по аксону в одном направлении (от тела нейрона к нервным окончаниям), хотя деполяризация мембраны происходит по обе стороны от участка, где в данный момент возник потенциал.
При прочих равных условиях распространение потенциала действия по аксону происходит тем быстрее, чем больше диаметр волокна. По гигантским аксонам кальмара потенциал действия может распространяться почти с такой же скоростью, как и по миелинизированным волокнам позвоночных (около 100 м/c).
14. Законы проведения возбуждения по нервным волокнам.
Существует четыре закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Закон изолированного проведения возбуждения.
Возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно.
В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
Закон бездекрементного проведения возбуждения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются, проходя по ним.
1. Функциональное состояние. Физиологические механизмы сна.
Функциональные состояния – фоновая активность высших отделов мозга, в условиях которой осуществляется деятельность. Изменение ФС сопровождается изменением физиологических свойств. Уровень ФС коррелирует с эффективностью деятельности. Оптимальный уровень ФС зависит от вида деятельности, качества сна, образа жизни, эмоционального фона. Методы оценки: уровень общей двигательной активности, напряжение скелетных мышц, изменение сердечного ритма, ЧСС, поведенческие показатели.
Сон – потеря психических связей субъекта с окружающим миром. Виды: монофазный (приурочен к суточной смене дня и ночи), полифазный (смена сна и бодрствования происходит несколько раз в сутки), сезонный (спячка); наркотический, гипнотический, патологический (травмы).
Сон человека имеет правильную циклическую организацию. В течение сна различают пять стадий. Четыре стадии медленноволнового сна и одна стадия быстрого. Завершенным циклом считается отрезок сна, в котором происходит последовательная смена стадий медленноволнового сна быстрым сном. В среднем отмечается 4-6 таких циклов за ночь, продолжительностью примерно 1,5 часа каждый.
· I стадия (дремота) занимает в среднем 12,1%. Является переходной от состояния бодрствования ко сну, что сопровождается уменьшением альфа-активности и появлением низкоамплитудных медленных тета- и дельта-волн. Длительность первой стадии обычно не больше 10-15 мин. В поведении эта стадия соответствует периоду дремоты с полусонными мечтаниями, она может быть связана с рождением интуитивных идей, способствующих успешности решения той или иной проблемы.
· II стадия (сонные веретена) — 38,1 %. Наиболее яркой ее чертой является наличие в ЭЭГ веретенообразной ритмической активности с частотой колебания 12-16 Гц. Длительность этих "веретен", хорошо выделяющихся из фоновой высокоамплитудной ЭЭГ со смешанной частотой колебаний, составляет от 0,2 до 0,5 секунд.
· III стадия (дельта сон) — 14,2%. Характеризуется всеми чертами второй стадии, к которым добавляется наличие в ЭЭГ медленных дельта колебаний с частотой 2 Гц и менее, занимающих от 20 до 50% эпохи записи. Этот переходный период длится всего несколько минут.
· IV стадия (дельта сон) — 12,1%. Характеризуется преобладанием в ЭЭГ медленных дельта колебаний с частотой 2 Гц и менее, занимающих более 50% эпохи записи ночного сна. Третья и четвертые стадии обычно объединяют под названием дельта-сна. Глубокие стадии дельта-сна более выражены в начале и постепенно уменьшаются к концу сна. В этой стадии разбудить человека достаточно трудно. Именно в это время возникают около 80% сновидений, и именно в этой стадии возможны приступы лунатизма и ночные кошмары, однако человек почти ничего из этого не помнит.
· V стадия (парадоксальный сон, быстрых движений глаз) — 23,5%. Во время этой стадии человек находится в полной неподвижности вследствие резкого падения мышечного тонуса, и лишь глазные яблоки под сомкнутыми веками совершают быстрые движения с частотой 60-70 раз в секунду. Электроэнцефалограмма приобретает признаки, характерные для состояния бодрствования (в спектре преобладают низкоамплитудные высокочастотные составляющие). Название "парадоксальная" возникло из-за видимого несоответствия между состоянием тела и активностью мозга. Если в это время разбудить спящего, то приблизительно в 90% случаев можно услышать рассказ о ярком сновидении, причем точность деталей будет существенно выше, чем при пробуждении из медленного сна.
По некоторым представлениям, существует иерархически построенная мозговая система, регулирующая циклы сна и бодрствования. В регулируемый контур этой системы входит ретикулярная активирующая система, поддерживающая уровень бодрствования, синхронизирующие аппараты, ответственные за медленный сон и ретикулярные ядра варолиева моста, ответственные за быстрый сон. В регулирующем контуре осуществляется взаимодействие между этим аппаратами, их включение и смена. Там же сон и бодрствование связаны с другими системами организма — вегетативной, соматической, психической. Несомненно, что регулирующий контур находится в пределах лимбико-ретикулярного комплекса. Внутри этого комплекса имеются и специализированные аппараты (сосудодвигательный и дыхательный центры, ядра гипоталамуса, регулирующие деятельность гипофиза и усиливающие сдвиги во внутренней среде, зоны, обеспечивающие сон и бодрствование) и нейронные образования, осуществляющие интеграцию деятельности мозговых систем.
2. ФС. Стресс.
Стресс — неспецифическая реакция организма на воздействие (физическое или психологическое), нарушающее его гомеостаз, а также соответствующее состояние нервной системы организма (или организма в целом). Основатель теории стресса канадский ученый Ганс Селье.
Этапы развития стресса. Любой раздражитель, вызывающий стрессовую реакцию, должен вначале быть воспринят (хотя и не обязательно осознанно) сенсорными рецепторами периферической нервной системы. Восприняв это раздражение, рецепторы посылают импульсы по сенсорным путям периферической нервной системы к мозгу. В конечном счете все потоки нервной импульсации по восходящим путям поступают в кору больших полушарий, где осуществляется их содержательная, смысловая интерпретация. Результаты этой интерпретации по каналам обратной связи попадают в лимбическую систему. Если раздражитель истолковывается как угроза или вызов, провоцирующий ярко выраженную эмоциональную оценку, возникает стрессогенная реакция. Для многих людей активация эмоций (как отрицательных, так и положительных) является стимулом для стресса.
Селье выделил 3 стадии общего адаптационного синдрома:
· реакция тревоги (мобилизация адаптационных возможностей — возможности эти ограничены)
ü Дистресс. Негативный тип стресса, с которым организм не в силах справиться. Он подрывает здоровье человека и может привести к тяжелым заболеваниям. От стресса страдает иммунная система.
ü Эмоциональный стресс. Эмоциональным стрессом называют эмоциональные процессы, сопровождающие стресс, и ведущие к неблагоприятным изменениям в организме. Во время стресса, эмоциональная реакция развивается раньше других, активизируя вегетативную нервную систему и её эндокринное обеспечение. При длительном или многократно повторяющемся стрессе эмоциональное возбуждение может застаиваться, а функционирование организма — разлаживаться.
ü Психологический стресс. Психологический стресс, как вид стресса, понимается разными авторами по-разному, но многие авторы определяют его как стресс, обусловленный социальными факторами и их когнитивной интерпретацией.
Из-за действия раздражителя происходит смена заряда, возникает разность потенциалов, местный ток (кольцевой), происходит перезарядка, возникает разность потенциалов, местный ток, ПД вызывает ПД у другого участка. Скорость 5-15 м/с.
По немиелинизированному волокну потенциал действия распространяется непрерывно. Проведение нервного импульса начинается с распространением электрического поля. Возникший потенциал действия за счет электрического поля способен деполяризовать мембрану соседнего участка до критического уровня, в результате чего на соседнем участке генерируются новые потенциалы. Сам потенциал действия не перемещается, он исчезает там же, где возник. Главную роль в возникновении нового потенциал действия играет предыдущий.
Если внутриклеточным электродом раздражать аксон посередине, то потенциал действия будет распространяться в обоих направлениях. Обычно же потенциал действия распространяется по аксону в одном направлении (от тела нейрона к нервным окончаниям), хотя деполяризация мембраны происходит по обе стороны от участка, где в данный момент возник потенциал.
При прочих равных условиях распространение потенциала действия по аксону происходит тем быстрее, чем больше диаметр волокна. По гигантским аксонам кальмара потенциал действия может распространяться почти с такой же скоростью, как и по миелинизированным волокнам позвоночных (около 100 м/c).
Законы проведения возбуждения по нервным волокнам.
Существует четыре закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Закон изолированного проведения возбуждения.
Возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно.
В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
Закон бездекрементного проведения возбуждения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются, проходя по ним.
17 . Физиология синапсов, классификация. Строения и механизм работы.
Синапс - это специализированная структура, которая обеспечивает передачу возбуждения с одной возбудимой структуры на другую. Термин "синапс" введен Ч. Шеррингтоном и означает "сведение", "соединение", "застежка".
Классификация синапсов. Синапсы можно классифицировать по:
1) их местоположению и принадлежности соответствующим структурам:
• периферические (нервно-мышечные, нейро-секреторные, рецеп-торнонейрональные);
• центральные (аксо-соматические, аксо-дендритные, аксо-аксо-нальные, сомато-девдритные, сомато-соматические);
2) знаку их действия - возбуждающие и тормозящие;
3) способу передачи сигналов - химические, электрические, смешанные.
4) медиатору, с помощью которого осуществляется передача - холинергические, адренергические, серотонинергические, глицинергические и т. д.
Строение синапса. Все синапсы имеют много общего, поэтому строение синапса и механизм передачи возбуждения в нем можно рассмотреть на примере нервно-мышечного синапса (рис. 7).
Синапс состоит из трех основных элементов:
• пресинаптической мембраны (в нервно-мышечном синапсе - это утолщенная концевая пластинка);
Пресинаптическая мембрана - это часть мембраны нервного окончания в области контакта его с мышечным волокном. Постси-наптическая мембрана - часть мембраны мышечного волокна. Часть постсинаптической мембраны, которая расположена напротив пресинаптической, называется субсинаптической мембраной. Осо-бенностью субсинаптической мембраны является наличие в ней специальных рецепторов,чувствительных к определенному медиатору, и наличие хемозависимых каналов. В постсинаптической мем-брне, за пределами субсинаптической, имеются потенциалозависимые каналы.
Механизм передачи возбуждения в химических возбуждающих синапсах. В синапсах с химической передачей возбуждение передатся с помощью медиаторов (посредников). Медиаторы - это химические вещества, которые обеспечивают передачу возбуждения в синапсах.
18.Медиаторы, их классификация.
Медиатор– это группа химических веществ, которая принимает участие в передаче возбуждения или торможения в химических синапсах с пресинаптической на постсинаптическую мембрану.
Критерии, по которым вещество относят к группе медиаторов:
1) вещество должно выделяться на пресинаптической мембране, терминали аксона;
2) в структурах синапса должны существовать ферменты, которые способствуют синтезу и распаду медиатора, а также должны быть рецепторы на постсинаптической мембране, которые взаимодействуют с медиатором;
3) вещество, претендующее на роль медиатора, должно при очень низкой своей концентрации передавать возбуждение с пресинаптической мембраны на постсинаптическую мембрану. Классификация медиаторов:
1) химическая, основанная на структуре медиатора;
2) функциональная, основанная на функции медиатора.
Химическая классификация.
1. Сложные эфиры – ацетилхолин (АХ).
2. Биогенные амины:
1) катехоламины (дофамин, норадреналин (НА), адреналин (А));
1) гаммааминомасляная кислота (ГАМК);
2) глютаминовая кислота;
1) опиоидные пептиды:
3) вазоактивный интестинальный пептид;
5. Пуриновые соединения: АТФ.
6. Вещества с минимальной молекулярной массой:
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).
Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим.
Проведение возбуждения в нервных волокнах
Проведение нервных импульсов по безмякотным и мякотным нервным волокнам принципиально отличается друг от друга. Нервный импульс по безмякотному нервному волокну, не имеющему на своей поверхности электрического изолятора в виде миелина, проводится непрерывно(от точки к точке), вовлекая последовательно в процесс возбуждения все участки волокна (такой способ проведения возбуждения в безмякотных нервных волокнах был впервые описан Германом в 1899 г.).
Проведение возбуждения по безмякотному нервному волокну
На рис. 1. Участок А нервного волокна, изображенный на данном рисунке, только что был возбужден, его мембрана уже частично реполяризована после предыдущего возбуждения, но он находится пока еще в состоянии относительной рефрактерности. Этот участок, будучи ранее возбужденным, уже вызвал возбуждение соседнего с ним участка В. В данный момент (указанный на рисунке) возбужден участок В: его мембрана заряжена противоположно по отношению к заряду мембраны соседних с ним участков – участка А (только что частично реполяризованного, но еще остающегося рефрактерным) и участка С (покоящегося). Вследствие разных зарядов мембраны участка В и соседних с ним участков А и С, между этими участками возникают местные ионные токи, направленные от "плюса" к "минусу". Причем, ток, направленный от участка В к участку А, будет деполяризующим по отношению к участку А, поскольку по аксоплазме волокна в сторону участка А, заряженного изнутри отрицательно, движутся положительные ионы. Но такой деполяризующий ток не способен вызвать снова возбуждение в участке А, поскольку участок А еще находится в состоянии относительной рефрактерности после только что прошедшего в нем возбуждения. Ток, от участка А к возбужденному соседнему участку В, идущий снаружи нервного волокна (по межклеточной жидкости), будет деполяризующим по отношению кучастку В, но никаких изменений в нем в данный момент этот ток вызвать не может, поскольку участок В возбуждени абсолютно рефрактерен. Аналогично, ток, идущий от покоящегося участка С к возбужденному в данный момент участку В, хотя и является деполяризующим по отношению к участку В, никоим образом не влияет на него, поскольку участок В временно абсолютно рефрактерен (его мембрана заряжена противоположно по отношению к уровню покоя, что соответствует верхней трети пика потенциала действия, в момент которой возбудимая структура абсолютно рефрактерна). Наконец, ионный ток, идущий от возбужденного участка В к покоящемуся участку С по аксоплазме нервного волокна, будет деполяризующим по отношению кпокоящемуся и нормально возбудимому участку С и вызовет начальную пассивную деполяризацию этого участка, которая повлечет за собой активную деполяризацию и последующую генерацию нервного импульса в нем. Затем возбуждение, аналогично описанной схеме, от участка С сможет передаться на следующий за ним покоящийся участок нервного волокна, но при этом вернуться назад на предыдущий, ранее бывший возбужденным, участок не сможет в связи с временной относительной его рефрактерностью после возбуждения. Таким образом, возбуждение в виде нервного импульса будет проходить последовательно по всему безмякотному нервному волокну, захватывая все новые и новые его участки. Скорость проведения возбуждения по безмякотному нервному волокну пропорциональна корню квадратному из диаметра волокна.
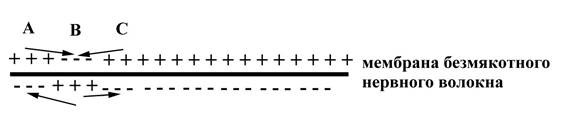
Рис. 1. Схема, отражающая особенности проведения возбуждения по безмякотному нервному волокну (по Косицкому Г.И., 1985). Стрелками показано направление местных ионных токов, возникающих между неодинаково заряженными участками мембраны волокна.
1.2. Возбуждение по мякотным нервным волокнам
Возбуждение в данном типе волокон проводится скачкообразно (сальтоторно): оно возникает только на мембране перехватов Ранвье, не имеющих электрического изолятора (миелина), и, перепрыгивая через межперехватный участок (покрытый миелином), достигает соседнего перехвата Ранвье, инициируя генерацию в нем нового нервного импульса. Так, например, если мембрана перехвата Ранвье, обозначенного буквой В (см. рис. 2), возбуждена (деполяризована), а мембрана соседнего перехвата (С) нормально поляризована (покоится), то между этими перехватами Ранвье, имеющими различный заряд на мембране, будут возникать местные ионные токи. Ионный ток, направленный от участка В к участку С, приведет к деполяризации участка С. Подобным образом возбуждение будет постепенно последовательно передаваться на следующие перехваты Ранвье нервного волокна. Ионный же ток от участка В (возбужденного) к участку А (реполяризующемуся после возбуждения) не приведет к генерации в нем нового нервного импульса, потому что участок А находится в состоянии невозбудимости (рефрактерности) после предыдущего потенциала действия, вызвавшего возбуждение в самом участке В.
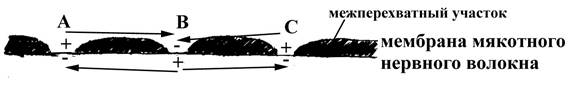
Рис. 2. Схема, отражающая особенности проведения возбуждения по мякотному нервному волокну (по Косицкому Г.И., 1985). Стрелками показано направление местных ионных токов, возникающих между участками мембраны по-разному заряженных перехватов Ранвье.
Время проведения нервного импульса от одного перехвата Ранвье к соседнему не зависит от длины межперехватного участка и при температуре 24°С составляет 0,07 мс. Вместе с тем длина межперехватного участка тем больше, чем больше диаметр волокна. Следовательно, скорость проведения возбуждения по мякотному нервному волокну зависит от его диаметра (пропорциональна диаметру волокна). Впервые предположение о сальтоторном способе проведении возбуждения было высказано Вериго в 1899 г, а экспериментально подтвержден такой способ проведения возбуждения Като (1924 г.) и Тасаки (1953 г.).
Сальтоторный способ проведения возбуждения по мякотному волокнуимеет ряд преимуществ по сравнению с непрерывным проведением в безмякотных:
Ø более быстрый (поскольку возбуждение перепрыгивает через межперехватные участки);
Ø энергетически более экономный (поскольку в состояние возбуждения приходит не вся мембрана волокна, а только лишь перехваты Ранвье. Как следствие, потери ионов на единицу длины волокна при возбуждении гораздо меньше таковых в безмякотном волокне, а, значит, и меньше тратится энергии на работу Na + /К + -насоса, восстанавливающего нарушенные трансмембранные градиенты для ионов Na + и К + после каждого возбуждения).
Читайте также:
