
Механизм возникновения и проведения нервного импульса
Механизм действия медиаторов.
Свойства сигнальных веществ нейромедиаторов, нейромодуляторов
· хранятся в везикулах;
· при поступлении нервного импульса выделяются в синаптическую щель,
· избирательно связываются со специфическим рецептором на постсинаптической мембране другого нейрона или мышечной клетки,
· индуцируют или ингибируют потенциал действия, контролируя возбуждение соседних нейронов и стимулируя клетки к выполнению специфических функций.
Синапсы образованы мембранами двух контактирующих клеток (пресинаптической и постсинаптической), которые разделены узкой синаптической щелью. Медиатор (или модулятор) выделяется в синаптическую щель за счет экзоцитоза, диффундирует к рецепторам постсинаптической мембраны, связывается с ними и передаёт сигнал соседней клетке.
Белки-рецепторы:
1) лиганд-активируемые ионные каналы – никотиновые холинэргические рецепторы (мышечные и нейрональные), ГАМК-рецепторы и глициновый рецептор. Вещество, действующее на ионотропные рецепторы, выступает медиатором. Большинство нейромедиаторов стимулирует открывание ионных каналов, и лишь немногие — закрывание.
2) чаще встречаются мембранные белки, которые управляют ионными каналами опосредованно с участием G-белков. Вещество, действущее на метаботропные рецепторы, выступает модулятором.
Клетки сообщаются друг с другом путём передачи сигналов, которые регистрируются в виде электрических процессов. Характер изменения мембранного потенциала постсинаптической клетки зависит от типа канала.
1)Изменение мембранного потенциала от -60 до +30 мВ за счёт открывания Nа + -каналов ведет к возникновению постсинаптического потенциала действия.
2)Изменение мембранного потенциала с -60 мВ до -90 мВ за счёт открывания Cl – -каналов ингибирует потенциал действия (гиперполяризация), в результате возбуждение не передается (тормозной синапс).
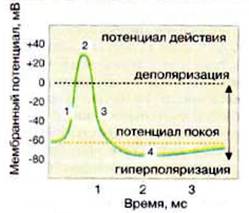
Рис. 6.Мембранный потенциал постсинаптической клетки
В состоянии покоя внутренняя сторона мембраны нейрона заряжена отрицательно. Это потенциал покоя, величина его достигает 60-75 мВ. Он обеспечивается в основном работой натриевого насоса (Na + ,К + -АТФазы). За счет энергии гидролиза каждой молекулы АТФ три иона Na + выкачиваются из клетки во внеклеточное пространство, а два иона К + перемещаются в противоположном направлении.
Разность потенциалов возникает в основном за счет следующих явлений. Несмотря на то что мембрана достаточно проницаема для ионов К + , их концентрация внутри аксона в 20-50 раз выше, чем во внешней среде. Главные внутриклеточные анионы (белки и нуклеиновые кислоты) не могут выходить наружу, а ионы С1 – , которых много во внешней среде, проходят через мембрану очень медленно. Проницаемость для ионов Na + составляет лишь 1/20 по сравнению с проницаемостью для ионов К + . При таких условиях трансмембранный потенциал определяется распределением ионов К + . Этот потенциал существует благодаря тому, что ионы К + стремятся покинуть клетку, чтобы уравнять внешнюю и внутреннюю концентрации. Однако в клетке при этом остается избыток анионов, что создает отрицательный электрический заряд, ограничивающий дальнейшее выравнивание концентраций ионов К + . Ионы С1 – ведут себя наоборот: они должны остаться снаружи, чтобы сбалансировать электрический заряд плохо проникающего Na + , но в то же время стремятся проникнуть в клетку по градиенту концентрации.
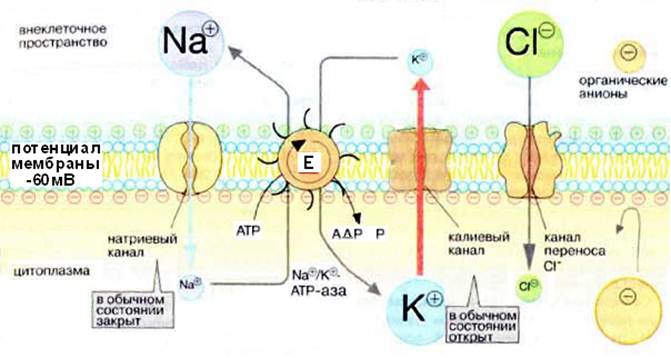
Рис. 6.Механизмы поддержания мембранного потенциала покоя
в постсинаптической клетке
За один импульс через мембрану проходит небольшое количество ионов Na + и К + , и концентрационные градиенты обоих ионов сохраняются: в клетке выше уровень К + , а вне клетки выше уровень Na + . Поэтому по мере получения клеткой новых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно.
Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращение мембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку.
Нервный импульс
Строго говоря, движущееся по нервам возбуждение представляет собой нервные импульсы, а не потенциалы действия.
Можно сказать короче:
Но в физиологической литературе в качестве синонима для нервного импульса принято использовать также и термин "потенциал действия". Хотя потенциал действия - это только электрический компонент нервного импульса.
Потенциал действия – это резкое скачкообразное изменение мембранного потенциала с отрицательного на положительный и обратно.
Потенциал действия - это электрическая характеристика (электрическая составляющая) нервного импульса.
Нервный импульс - это сложный структурно-электро-химический процесс, распространяющийся по мембране нейрона в виде бегущей волны изменений.
Потенциал действия - это только электрический компонент нервного импульса, характеризующий изменения электрического заряда (потенциала) на локальном участке мембраны во время прохождения через него нервного импульса (от -70 до +30 мВ и обратно). (Кликните на изображение слева, чтобы увидеть анимацию.)
Сравните два приведённых выше рисунка (покликайте по ним) и, как говорится, почувствуйте разницу!
Где рождаются нервные импульсы?
Как ни странно, не все студенты, изучившие физиологию возбуждения, могут ответить на этот вопрос. ((
Хотя ответ не сложен. Нервные импульсы рождаются на нейронах всего в нескольких местах:
1) аксонный холмик (это переход тела нейрона в аксон),
2) рецепторное окончание дендрита,
3) первый перехват Ранвье на дендрите (триггерная зона дендрита),
4) постсинаптическая мембрана возбуждающего синапса.
Места возникновения нервных импульсов:
1. Аксонный холмик - главный породитель нервных импульсов.
Аксонный холмик - это самое начало аксона, там где он начинается на теле нейрона. Именно аксонный холмик является главным породителем (генератором) нервных импульсов на нейроне. Во всех остальных местах вероятность рождения нервного импульса намного меньше. Дело в том, что у мембраны аксонного холмика повышена чувствительность к возбуждению и понижен критический уровень деполяризации (КУД) по сравнению с остальными участками мембраны. Поэтому, когда на мембране нейрона начинают суммироваться многочисленные возбуждающие постсинаптические потенциалы (ВПСП), которые возникают в самых разных местах на постсинаптических мембранах всех его синаптических контактов, то раньше всего КУД достигается именно на аксонном холмике. Там-то эта сверхпороговая для холмика деполяризация и открывает потенциал-чувствительные натриевые каналы, в которые входит поток ионов натрия, порождающий потенциал действия и нервный импульс.
Итак, аксонный холмик является интегративной зоной на мембране, он интегрирует все возникающие на нейроне локальные потенциалы (возбуждающие и тормозные) - и первый срабатывает на достижение КУД, порождая нервный импульс.
Важно также учесть следующий факт. От аксонного холмика нервный импульс разбегается по всей мембране своего нейрона: как по аксону к пресинаптическоим окончаниям, так и по дендритам к постсинаптическим "начинаниям". Все локальные потенциалы при этом снимаются с мембраны нейрона и со всех его синапсов, т.к. они "перебиваются" потенциалом действия от пробегающего по всей мембране нервного импульса.
2. Рецепторное окончание чувствительного (афферентного) нейрона.
Если нейрон имеет рецепторное окончание, то на него может воздействовать адекватный раздражитель и порождать на этом окончании сначала генераторный потенциал, а затем и нервный импульс. Когда генераторный потенциал достигает КУД, то на этом окончании открываются потенциал-зависимые натриевые ионные каналы и рождается потенциал действия и нервный импульс. Нервный импульс бежит по дендриту к телу нейрона, а затем по его аксону к пресинаптическим окончаниям для передачи возбуждения на следующий нейрон. Так работают, к примеру, болевые рецепторы (ноцицепторы), являющиеся дендритными окончаниями болевых нейронов. Нервные импульсы в болевых нейронах вознимают именно на рецепторных окончаниях дендритов.
3. Первый перехват Ранвье на дендрите (триггерная зона дендрита).
Локальные возбуждающие постсинаптические потенциалы (ВПСП) на окончаниях дендрита, которые формируются в ответ на возбуждения, приходящие к дендриту через синапсы, суммируются на первом перехвате Ранвье этого дендрита, если он, конечно, миелинизирован. Там находится участок мембраны с повышенной чувствительностью к возбуждению (пониженным порогом), поэтому именно в этом участке легче всего преодолевается критический уровень деполяризации (КУД), после чего открываются потенциал-управляемые ионные каналы для натрия - и возникает потенциал действия (нервный импульс).
4. Постсинаптическая мембрана возбуждающего синапса.
В редких случаях ВПСП на возбуждающем синапсе может быть настолько силён, что прямо там же достигает КУД и порождает нервный импульс. Но чаще это бывает возможно только в результате суммации нескольких ВПСП: или с нескольких соседних синапсов, сработавших одновременно (пространственная суммация), или за счёт того, что на данный синапс пришло несколько импульсов подряд (временная суммация).
Видео: Проведение нервного импульса по нервному волокну
Потенциал действия как нервный импульс
Ниже размещён материал, взятый из учебно-методического пособия автора данного сайта, на который вполне можно ссылаться в своём списке литературы:
Сазонов В.Ф. Понятие и виды торможения в физиологии центральной нервной системы: Учебно-методическое пособие. Ч. 1. Рязань: РГПУ, 2004. 80 с.
Все процессы мембранных изменений, происходящих в ходе распространяющегося возбуждения, достаточно хорошо изучены и описаны в научной и учебной литературе. Но не всегда это описание легко понять, поскольку в данном процессе задействовано слишком много компонентов (с точки зрения обычного студента, а не вундеркинда, конечно).
Для облегчения понимания мы предлагаем рассматривать единый электрохимический процесс распространяющегося динамичного возбуждения с трех сторон, на трех уровнях:
Электрические явления – развитие потенциала действия.
Химические явления – движение ионных потоков.
Структурные явления – поведение ионных каналов.
Три стороны процесса распространяющегося возбуждения
1. Потенциал действия (ПД)
Потенциал действия – это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно.
Обычно мембранный потенциал в нейронах ЦНС изменяется от –70 мВ до +30 мВ, а затем вновь возвращается к исходному состоянию, т.е. к –70 мВ. Как видим, понятие потенциала действия характеризуется через электрические явления на мембране.
На электрическом уровне изменения начинаются как смена поляризованного состояния мембраны на деполяризацию. Сначала деполяризация идет в виде локального возбуждающего потенциала. Вплоть до критического уровня деполяризации (примерно –50 мВ) это относительно простое линейное уменьшение электроотрицательности, пропорциональное силе воздействующего раздражителя. А вот потом начинается более крутая самоусиливающаяся деполяризация, она развивается не с постоянной скоростью, а с ускорением . Говоря образно, деполяризация так разгоняется, что перескакивает через нулевую отметку, не заметив этого, и даже переходит в положительную поляризацию. После достижения пика (обычно +30 мВ) начинается обратный процесс – реполяризация , т.е. восстановление отрицательной поляризации мембраны.
Кратко опишем электрические явления во время течения потенциала действия:
Восходящая ветвь графика:
потенциал покоя – исходное обычное поляризованное электроотрицательное состояние мембраны (–70 мВ);
нарастающий локальный потенциал – пропорциональная раздражителю деполяризация;
критический уровень деполяризации (–50 мВ) – резкое ускорение деполяризации (за счет самораскрытия натриевых каналов), с этой точки начинается спайк – высокоамплитудная часть потенциала действия;
самоусиливающаяся круто нарастающая деполяризация;
переход нулевой отметки (0 мВ) – смена полярности мембраны;
пик (+30 мВ) – вершина процесса изменения полярности мембраны, вершина потенциала действия.
Нисходящая ветвь графика:
реполяризация – восстановление прежней электроотрицательности мембраны;
переход нулевой отметки (0 мВ) – обратная смена полярности мембраны на прежнюю, отрицательную;
переход критического уровня деполяризации (–50 мВ) – прекращение фазы относительной рефрактерности (невозбудимости) и возврат возбудимости;
следовые процессы (следовая деполяризация или следовая гиперполяризация);
восстановление потенциала покоя – норма (–70 мВ).
Итак, сначала – деполяризация, затем – реполяризация. Сначала – утрата электроотрицательности, затем – восстановление электроотрицательности.
2. Ионные потоки
Итак, важно осознать, что все электрические токи, которые идут через мембрану, являются ионными потоками . Привычного нам из физики тока в виде потока электронов в клетках, как в водных системах, просто нет. Ссылки на потоки электронов будут ошибкой.
На химическом уровне мы, описывая распространяющееся возбуждение, должны рассмотреть, как изменяются характеристики ионных потоков, идущих через мембрану. Главное в этом процессе то, что при деполяризации резко усиливается поток ионов натрия внутрь клетки, а затем он внезапно прекращается на спайке потенциала действия. Входящий поток натрия как раз и вызывает деполяризацию, так как ионы натрия приносят с собой положительные заряды в клетку (чем и снижают электроотрицательность). Затем, после спайка, значительно нарастает выходящий наружу поток ионов калия, что вызывает реполяризацию. Ведь калий, как мы неоднократно говорили, выносит с собой из клетки положительные заряды. Отрицательные заряды остаются внутри клетки в большинстве, и за счет этого усиливается электроотрицательность. Это и есть восстановление поляризации за счет выходящего потока ионов калия. Заметим, что выходящий поток ионов калия возникает практически одновременно с появлением натриевого потока, но нарастает медленно и длится в 10 раз дольше. Несмотря на продолжительность калиевого потока самих ионов расходуется немного – всего одна миллионная доля от запаса калия в клетке (0,000001 часть).
Подведем итоги. Восходящая ветвь графика потенциала действия образуется за счет входа в клетку ионов натрия, а нисходящая – за счет выхода из клетки ионов калия.
3. Ионные каналы
Все три стороны процесса возбуждения – электрическая, химическая и структурная – необходимы для понимания его сущности. Но все-таки все начинается с работы ионных каналов. Именно состояние ионных каналов предопределяет поведение ионов, а поведение ионов в свою очередь сопровождается электрическими явлениями. Начинают процесс возбуждения натриевые каналы .
Пожар как метафора распространяющегося возбуждения
Когда мы искали образный пример, аналогию или метафору, которая может наглядно передать смысл распространяющегося возбуждения, то остановились на образе пожара. Действительно, распространяющееся возбуждение похоже на лесной пожар, когда горящие деревья остаются на месте, а фронт огня распространяется и уходит все дальше и дальше во все стороны от очага возгорания.
Как же в этой метафоре будет выглядеть явление торможения?
Ответ очевиден – торможение будет выглядеть как тушение пожара, как уменьшение горения и затухание огня. Но если огонь распространяется сам по себе, то тушение требует усилий. Из потушенного участка процесс тушения сам по себе не пойдет во все стороны.
Существует три варианта борьбы с пожаром: (1) либо надо ждать, когда все сгорит и огонь истощит все горючие запасы, (2) либо надо поливать водой горящие участки, чтобы они погасли, (3) либо надо поливать заранее ближайшие нетронутые огнем участки, чтобы они не загорелись.
АВТОВОЛНЫ В АКТИВНО-ВОЗБУДИМЫХ СРЕДАХ (АВС)
При распространении волны в активно-возбудимых средах не происходит переноса энергии. Энергия не переносится, а освобождается, когда до участка АВС доходит возбуждение. Можно провести аналогию с серией взрывов зарядов, заложенных на некотором расстоянии друг от друга (например, при тушении лесных пожаров, строительстве, мелиоративных работах), когда взрыв одного заряда вызывает взрыв рядом расположенного и так далее. Лесной пожар также является примером распространения волны в активно- возбудимой среде. Пламя распространяется по области с распределенными запасами энергии - деревья, валежник, сухой мох.
Основные свойства волн, распространяющихся в активно-возбудимых средах (АВС)
Волна возбуждения распространяется в АВС без затухания; прохождение волны возбуждения связано с рефрактерностью - невозбудимостью среды в течение некоторого промежутка времени (периода рефрактерности).
Видео: Потенциал действия (Action potential)
Возникновение нервного импульса связано с состоянием мембраны нейрона. Биологические мембраны, будучи липидно-белковым комплексом, непроницаемы для заряженных ионов. Перемещение ионов через мембрану осуществляется с помощью специальных транспортных систем: натриевого насоса, работающего с затратой энергии (Na + , К + -АТФ-аза) и ионпроводящих каналов – натриевых и калиевых.
В состоянии физиологического покоя по разные стороны мембраны имеет место разность концентраций ионов, главным образом Na + и К + . Концентрация ионов К + выше в клетке, ионов Na + – в межклеточной жидкости. Это влияет и на распределение других ионов. Разность концентраций ионов поддерживается с помощью натриевого насоса, перекачивающего ионы против градиента концентрации (в противном случае диффузия ионов по градиенту концентрации привела бы к выравниванию концентраций ионов по обе стороны мембраны и гибели клетки) Таким образом, внутри аксона образуется избыток (–) зарядов, снаружи – (+) зарядов, то есть возникает разность электрических потенциалов – потенциал покоя (ПП). Его значение составляет – 60-70 мВ. ПП одинаков по всей длине нервного волокна (рис. 4).
Раздражение нервного волокна приводит сначала к открытию Na + - и К + - каналов. Раньше открываются Na + -каналы, и в клетку устремляется поток ионов натрия. Это изменяет трансмембранный потенциал: сначала он становится равным 0 (деполяризация мембраны), затем происходит перезарядка мембраны, внутренняя сторона мембраны приобретает (+) заряд, наружная (-). Разность потенциалов при этом достигает 40 мВ. Эта величина и есть потенциал действия (ПД)
Натриевые каналы закрываются, открываются калиевые, начинается выход калия из клетки, потенциал изменяется от – 40 мВ до – 70 мВ, то есть идет реполяризация мембраны. После закрытия ионных каналов ионный насос восстанавливает исходное распределение ионов по сторонам мембраны.
Механизмы памяти.
Концепции биохимического кодирования индивидуального опыта в памяти опираются на две группы фактов:
· возможность передачи приобретенной информации необученному мозгу с помощью этих факторов.
Первые гипотезы, связывающие запечатление информации с биохимическими изменениями в нервной ткани родились на основе широко известных в 60-е гг. опытов Г. Хидена, когда было показано, что образование следов памяти сопровождается изменениями свойств РНК и белка в нейронах.
Выяснилось, что раздражение нервной клетки увеличивает в ней содержание РНК и оставляет длительные биохимические следы, сообщающие клетке способность резонировать в ответ на повторные действия одних и тех же раздражителей.
Таким образом, было установлено, что РНК играет важную роль в механизмах формирования и сохранения следов памяти.
Однако в более поздних работах было показано, что в консолидации энграмм памяти ведущую роль играет ДНК, которая может служить хранилищем не только генетической, но и приобретенной информации, а РНК обеспечивает передачу специфического информационного кода.
В настоящее время идея существования биохимических факторов, способных к сохранению и переносу информации, большинством исследователей воспринимается критически. Считается, что гипотеза молекулярного кодирования индивидуального опыта не имеет прямых фактических доказательств.
Медиаторам - химическим посредникам в синаптической передаче информации - придается большое значение в обеспечении механизмов долговременной памяти. Основные медиаторные системы головного мозга принимают самое непосредственное участие в обучении и формировании энграмм памяти. Так, экспериментально установлено, что уменьшение количества норадреналина замедляет обучение, вызывает амнезию и нарушает извлечение следов из памяти.
Р.И. Кругликов (1986) разработал концепцию, в соответствии с которой в основе долговременной памяти лежат сложные структурно-химические преобразования на системном и клеточном уровнях головного мозга. Показано, что под влиянием обучения увеличивается количество холинорецепторов, то есть рецепторов, расположенных на теле нейрона и отвечающих за обнаружение медиатора ацетилхолина. В процессе образования условного рефлекса повышается чувствительность соответствующих нейронов к ацетилхолину, что облегчает обучение, ускоряет запоминание и способствует более быстрому извлечению следа из памяти. В то же время вещества, препятствующие действию ацетилхолина, нарушают обучение и воспроизведение, вызывая амнезию (потерю памяти).
Биохимические методы, которые позволяют проникнуть в последовательность процессов, происходящих в синаптических мембранах с последующим синтезом новых белков, привлекают многих исследователей памяти. На этом пути ожидаются новые яркие открытия. Предполагается, например, что для различных видов памяти в ближайшем будущем будут выявлены различия в биохимических процессах.
Тем не менее следует подчеркнуть, что интенсивные биохимические исследования привели к явной переоценке и автономизации клеточно-молекулярного уровня изучения механизмов памяти. Как указывает С. Роуз, эксперименты, проводимые только на клеточном уровне, слишком ограничены и, по-видимому, не способны ответить на вопрос, как мозг человека запоминает, например, сложные симфонические партитуры или извлекает из памяти данные, необходимые для разгадывания простого кроссворда.
Для более полного знания специфики функционирования процессов памяти необходим переход на уровень сложных мозговых систем, где многие нейроны соединены между собой морфологическими и функциональными связями. При этом психофизиологические исследования на здоровых людях позволяют изучать процессы переработки и хранения информации, а изучение больных с различного рода амнезиями, возникающими после повреждения мозга, позволяет глубже проникать в тайны памяти.
Березов стр. 641-642
Дата добавления: 2019-07-15 ; просмотров: 735 ;
Рассмотрим химические основы возникновения и поддержания биоэлектрических потенциалов (потенциала покоя и потенциала действия). Большинство исследователей придерживаются мнения, что явления электрической поляризации клетки обусловлены неравномерным распределением ионов К + и Na + по обе стороны клеточной мембраны. Мембрана обладает избирательной проницаемостью: большей для ионов К + и значительно меньшей для ионов Na + . Кроме того, в нервных клетках существует механизм, который поддерживает внутриклеточное содержание натрия на низком уровне вопреки градиенту концентрации. Этот механизм получил название натриевого насоса.
При определенных условиях резко повышается проницаемость мембраны для ионов Na + .
В состоянии покоя внутренняя сторона клеточной мембраны заряжена электроотрицательно по отношению к наружной поверхности. Объясняется это тем, что количество ионов Na + , выкачиваемых из клетки с помощью натриевого насоса, не вполне точно уравновешивается поступлением в клетку ионов К + . В связи с этим часть катионов натрия удерживается внутренним слоем противоионов (анионов) на наружной поверхности клеточной мембраны. Таким образом, на мембранах, ограничивающих нервные клетки, поддерживается разность электрических потенциалов (трансмембранная разность электрических потенциалов); эти мембраны электрически возбудимы.
При возбуждении, вызванном тем или иным агентом, селективно изменяется проницаемость мембраны нервной клетки (аксона): увеличивается избирательно для ионов Na + (примерно в 500 раз) и остается без изменения для ионов К + . В результате ионы Na + устремляются внутрь клетки. Компенсирующий поток ионов К + , направляющийся из клетки, несколько запаздывает. Это приводит к возникновению отрицательного заряда на наружной поверхности клеточной мембраны. Внутренняя поверхность мембраны приобретает положительный заряд; происходит перезарядка клеточной мембраны (в частности, мембраны аксона, т.е. нервного волокна), и возникает потенциал действия, или спайк. Продолжительность спайка не превышает 1 мс. Он имеет восходящую фазу, пик и нисходящую фазу. Нисходящая фаза (падение потенциала) связана с нарастающим преобладанием выхода ионов К + над поступлением ионов Na + – мембранный потенциал возвращается к норме. После проведения импульса в клетке восстанавливается состояние покоя. В этот период ионы Na + , вошедшие в нейрон при возбуждении, заменяются на ионы К + . Этот переход происходит против градиента концентрации, так как ионов Na + во внешней среде, окружающей нейроны, намного больше, чем в клетке после момента ее возбуждения. Переход ионов Na + против градиента концентрации, как отмечалось, осуществляется с помощью натриевого насоса, для работы которого необходима энергия АТФ. В конце концов все это приводит к восстановлению исходной концентрации катионов калия и натрия внутри клетки (аксона), и нерв готов для получения следующего импульса возбуждения. Заметим, что миелиновые мембраны, образуемые шванновскими клетками, окутывают нервные волокна и служат электрическим изолятором. Этот изоляционный слой покрывает большинство нервных волокон и сильно ускоряет распространение электрической волны (сигнала); при этом ионы входят в клетку и выходят из нее только в тех местах, где изолятор отсутствует. Как уже отмечалось, миелиновая мембрана состоит из фос-фолипидов, в частности из сфингомиелина, холестерина, а также белков и гликосфинголипидов. Некоторые заболевания, например рассеянный склероз, характеризуются демиелинизацией и нарушением проведения нервного импульса. Другим не менее важным процессом для нервной ткани является передача нервного импульса от одной нервной клетки к другой или воздействие на клетки эффекторного органа.
Нервная система человека выступает своеобразным координатором в нашем организме. Она передаёт команды от мозга мускулатуре, органам, тканям и обрабатывает сигналы, идущие от них. В качестве своеобразного носителя данных используется нервный импульс. Что он собой представляет? С какой скоростью работает? На эти, а также на ряд других вопросов можно будет найти ответ в этой статье.
Чем является нервный импульс?
Исследование строения и работы
Впервые прохождение нервного импульса было продемонстрировано немецкими учеными Э. Герингом и Г. Гельмгольцем на примере лягушки. Тогда же и было установлено, что биоэлектрический сигнал распространяется с указанной ранее скоростью. Вообще, такое является возможным благодаря особенному построению нервных волокон. В некотором роде они напоминают электрический кабель. Так, если проводить параллели с ним, то проводниками являются аксоны, а изоляторами – их миелиновые оболочки (они являют собой мембрану шванновской клетки, которая намотана в несколько слоев). Причем скорость нервного импульса зависит в первую очередь от диаметра волокон. Вторым по важности считается качество электрической изоляции. Кстати, в качестве материала организмом используется липопротеид миелин, который обладает свойствами диэлектрика. При прочих равных условиях, чем больше будет его слой, тем быстрее будут проходить нервные импульсы. Даже на данный момент нельзя сказать, что эта система полноценно исследована. Многое, что относится к нервам и импульсам, ещё остаётся загадкой и предметом исследования.
Особенности строения и функционирования
Где они создаются?
Типы клеток
- Рецепторные (чувствительные). Ими кодируются и превращаются в нервные импульсы все температурные, химические, звуковые, механические и световые раздражители.
- Вставочные (также называются кондукторными или замыкательными). Они служат для того, чтобы перерабатывать и переключать импульсы. Наибольшее их число находится в головном и спинном мозге человека.
- Эффекторные (двигательные). Они получают команды от центральной нервной системы на то, чтобы были совершены определённые действия (при ярком солнце закрыть рукой глаза и так далее).
Каждый нейрон имеет тело клетки и отросток. Путь нервного импульса по телу начинается именно с последнего. Отростки бывают двух типов:
- Дендриты. На них возложена функция восприятия раздражения расположенных на них рецепторов.
- Аксоны. Благодаря им нервные импульсы передаются от клеток к рабочему органу.
Интересный аспект деятельности
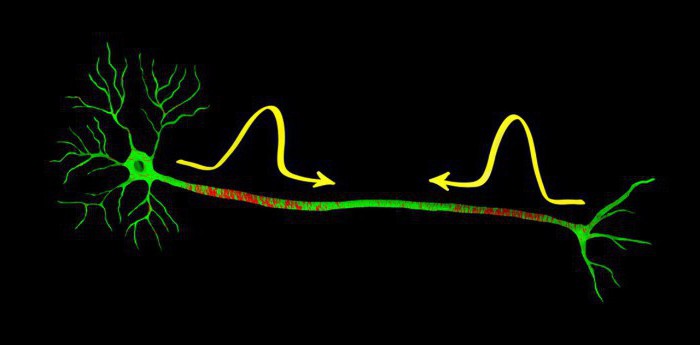
О потенциале действия
Как всё работает в мозгу?
Работа нейромедиаторов
Когда они передают нервные импульсы, то существует несколько вариантов, что произойдёт с ними:
- Они будут диффундированы.
- Подвергнутся химическому расщеплению.
- Вернутся назад в свои пузырьки (это называется обратным захватом).
В конце 20-го века сделали поразительное открытие. Ученые узнали, что лекарства, что влияют на нейромедиаторы (а также их выброс и обратный захват), могут изменять психическое состояние человека коренным образом. Так, к примеру, ряд антидепрессантов вроде "Прозака" блокируют обратный захват серотонина. Есть определённые причины считать, что в болезни Паркинсона виноват дефицит в головном мозге нейромедиатора дофамина.
Если кратко, то они могут работать с тысячами нейромедиаторов, которые посылаются их соседями. Детали относительно обработки и интеграции данного типа импульсов нам почти не известны. Хотя над этим работает много исследовательских групп. На данный момент получилось узнать, что все полученные импульсы интегрируются, а нейрон выносит решение – необходимо ли поддерживать потенциал действия и передавать их дальше. На этом фундаментальном процессе базируется функционирование головного мозга человека. Ну что ж, тогда это неудивительно, что мы не знаем ответа на эту загадку.
Некоторые теоретические особенности
Где же создаются нервные импульсы?
Откуда они начинают свой путь? Ответ на этот вопрос может дать любой студент, который прилежно изучал физиологию возбуждения. Есть четыре варианта:
- Рецепторное окончание дендрита. Если оно есть (что не факт), то возможным является наличие адекватного раздражителя, что создаст сначала генераторный потенциал, а потом уже и нервный импульс. Подобным образом работают болевые рецепторы.
- Мембрана возбуждающего синапса. Как правило, такое возможно только при наличии сильного раздражения или их суммирования.
- Триггерная зона дентрида. В этом случае локальные возбуждающие постсинаптические потенциалы формируются как ответ на раздражитель. Если первый перехват Ранвье миелинизирован, то они на нём суммируются. Благодаря наличию там участка мембраны, которая обладает повышенной чувствительностью, здесь возникает нервный импульс.
- Аксонный холмик. Так называют место, где начинается аксон. Холмик – это наиболее частый создать импульсов на нейроне. Во всех остальных местах, которые рассматривались ранее, их возникновение гораздо менее вероятное. Это происходит из-за того, что здесь мембрана имеет повышенную чувствительность, а также пониженный критический уровень деполяризации. Поэтому, когда начинается суммирование многочисленных возбуждающих постсинаптических потенциалов, то раньше всего на них реагирует холмик.
Пример распространяющегося возбуждения
Вспомните сводки из новостей прошлого лета (также это скоро можно будет услышать опять). Пожар распространяется! При этом деревья и кустарники, которые горят, остаются на своих местах. А вот фронт огня идёт всё дальше от места, где был очаг возгорания. Аналогичным образом работает нервная система.
Часто бывает необходимо успокоить начавшееся возбуждение нервной системы. Но это не так легко сделать, как и в случае с огнем. Для этого совершают искусственное вмешательство в работу нейрона (в лечебных целях) или используют различные физиологические средства. Это можно сравнить с заливанием пожара водой.
Читайте также:
