
Межклеточные контакты в нервной системе
ФЕДЕРАЛЬНОЕ АГЕНСТВО ПО ОБРАЗОВАНИЮ
ПЕНЗЕНСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ ИМ. В.Г. БЕЛИНСКОГО
ПО БИОХИМИИ МЕМБРАН:
Выполнила студентка IV курса группы БХ-41
Научный руководитель Соловьев В.Б.
1.Классификация межклеточных контактов
2. Замыкающие межклеточные контакты
2.1.Рыхлые или простые контакты
2.2.Плотный замыкающий контакт
2.3.Замыкающие межклеточные контакты
3.Адгезионные или сложные межклеточные контакты
3.1.Точечные межклеточные контакты
3.3.Адгезионные соединения между клеткой и матриксом
5.Заболевание аутоиммунное, поражающее десмосомы
6.Проводящие межклеточные контакты
1. Классификация межклеточных контактов
1.Замыкающие межклеточные контакты.
а) простой или рыхлый контакт;
б) плотный замыкающий контакт.
2.Адгезионные межклеточные контакты.
а) точечные контакты;
б) адгезионные пояски;
в) адгезионные соединения между клеткой и внутриклеточным матриксом;
2. Замыкающие межклеточные контакты
2.1 Рыхлые или простые контакты
Простой контакт — соединение клеток за счет пальцевидных впячиваний и выпячиваний цитомембран соседних клеток. Специфических структур, формирующих контакт, нет.
Простые контакты занимают наиболее обширные участки соприкасающихся клеток. Расстояние между билипидными мембранами соседних клеток составляет 15-20 нм, а связь между клетками осуществляется за счет взаимодействия макромолекул соприкасающихся гликокаликсов.
Посредством простых контактов осуществляется слабая механическая связь - адгезия, не препятствующая транспорту веществ в межклеточных пространствах. Разновидностью простого контакта является контакт "типа замка", когда плазмолеммы соседних клеток вместе с участком цитоплазмы как бы впячивается друг в друга (интердигитация), чем достигается большая поверхность соприкосновения и более прочная механическая связь.
2.2 Плотный замыкающий контакт
Плотный замыкающий контакт — соприкасаются билипидные слои мембран соседних клеток. В области зоны плотных контактов между клетками не проходят практически никакие вещества.
Постоянные клеточные контакты скрепляют клетки в эпителиальном клеточном слое таким образом, что предотвращается перетекание даже малых молекул с одной стороны слоя на другую. Латеральная подвижность многих мембранных белков ограничена. Ограничение подвижности достигается с помощью барьеров, образованных при участии плотных контактов.
Клоны эпителиальных тканей (эпителии) функционируют в качестве избирательно-проницаемых барьеров, разделяющих жидкости с разным химическим составом по обе стороны слоя. В выполнении этой функции плотные контакты играют две роли.
Осуществляемый эпителиальными клетками трансклеточный транспорт (например, питательных веществ полости тонкого кишечника во внутриклеточную жидкость по другую сторону слоя) зависит от двух групп мембранных белков-переносчиков: одна находится на апикальной (обращенной в полость) поверхности клетки и активно транспортирует отдельные молекулы в клетку; другая находится на базолатеральной поверхности клетки и позволяет тем же молекулам покидать клетку путем облегченной диффузии . Для поддержания этого направленного транспорта не должно происходить перемещения апикальных белков-переносчиков на базолатеральную поверхность и наоборот.
Кроме того, промежутки между эпителиальными клетками должны быть скреплены таким образом, чтобы транспортированные молекулы не могли бы продиффундировать назад в полость через межклеточные промежутки.
Плотные контакты и выполняют эти две функции: барьеров для диффузии мембранных белков между апикальной и базолатеральной поверхностями и скрепления соседних клеток вместе так, что водорастворимые молекулы не могут перетечь на другую сторону слоя. При этом плотные контакты непроницаемы для макромолекул, а их проницаемость для малых молекул сильно варьирует в разных эпителиях. Эпителиальнные клетки могут временно модифицировать плотные контакты с тем, чтобы допустить увеличенный ток жидкости через бреши в контактных барьерах. Такой параклеточный транспорт особенно важен при абсорбции аминокислот и моносахаридов из полости тонкого кишечника.
Важнейшим элементом в структуре избирательно проницаемых барьеров эпителиальных и эндотелиальных являются плотные контакты. Избирательная проницаемость варьирует от ткани к ткани, пропуская или целые клетки и макромолекулы, или только протоны и ионы. Плотный контакт выглядит как пояс из переплетающихся скрепляющих нитей, который полностью окружает апикальный конец каждой клетки эпителиального слоя. Полагают, что скрепляющие нити состоят из длинных рядов специфических трансмембранных белков в каждой из двух взаимодействующих плазматических мембран, и которые (белки) соединяются напрямую друг с другом, что приводит к закупориванию межклеточного пространства. Интегральным мембранным белком плотного соединения оказался окклудин (взаимодействует с двумя цитоплазматическими белками, ZO-1 и ZO-2 ( zonula occludence 1, 2 ). Их функция окончательно не ясна. Возможно, их роль заключается в локализации оккулдина в сайтах между апикальной и базолатеральной поверхностями клетки. Некоторые ассоциированные с цитоскелетом белки были также обнаружены в участках плотных контактов. Среди них зингулин, антиген и актин (по данным электронной микроскопии, актиновые филаменты состоят из двух цепей глобулярных молекул, диаметром 4 нм и образующих двойную спираль, на каждый виток которой приходится 13,5 молекулы). Эти цепи составляют основу тонких филаментов скелетных мышц, которые кроме актина содержат также несколько других белков; глобулярный актин имеет молекулярную массу около 42 кД. Он содержит одну полипептидную цепь, состоящую из 375 или 374 аминокислотных остатков; различия в аминокислотной последовательности у разных актинов, как в пределах одного вида, так и межвидовые, крайне незначительны. Они составляют не более 25 аминокислотных замен; в настоящее время у позвоночных животных различают 6 изоформ актина, в зависимости от изоэлектрической точки они делятся на 3 класса - альфа, бета и гамма; бета- и гамма-актины характерны для немышечных клеток , а альфа-актины - для мышечных). Ras играет определенную роль в регулировании функционирования плотных соединений. Таким образом, в клетках имеются, по-видимому, сходные механизмы построения и регуляции адгезионных структур, и эти механизмы тесно взаимосвязаны с изменениями в цитоскелете. Однако, каким образом перестройки цитоскелета влияют на процессы межклеточной адгезии, пока окончательно не ясно. Механизмы адгезии и межклеточной сигнализации тесно сопряжены с давно известным феноменом контактного торможения , природа которого до сих пор до конца не выяснена.
2.3 Замыкающие межклеточные контакты
Интегрины - представляют собой поверхностные гетеродимерные белки, которые обеспечивают адгезию клеток к компонентам внеклеточного матрикса и иногда к другим клеткам. Многие интегрины проявляют сродство к гликопротеидам и базальной мембраны, и внеклеточного матрикса. Утрата некоторых интегринов (при раке молочной железы, раке предстательной железы, раке толстой кишки) или их избыток (при меланоме, плоскоклеточном раке полости рта, носоглотки, гортани) сопряжены с высокой степенью злокачественности опухоли. Связывание интегринов с лигандами и сближение клеток необходимы для перестройки базальной мембраны, идущей при ангиогенезе . Взаимодействие интегринов с белками внеклеточного матрикса в некоторых случаях препятствует апоптозу . Так, клетки меланомы избегают апоптоза в дерме за счет связывания альфа(ню)-бета3-интегрина с коллагеном. Нейтрализация этого интегрина антителами, напротив, способствует апоптозу. Таким образом, информация, которую интегрины передают от внеклеточного матрикса внутрь клетки, в одних случаях стимулирует адгезию и миграцию опухолевых клеток, в других - приводит к их гибели. Иными словами, интегрины играют роль своеобразного "переключателя", определяющего дальнейшую судьбу опухолевой клетки.
Соединения клеток, присутствующих в тканях и органах многоклеточных организмов, образуются сложными структурами, которые именуются межклеточными контактами . Особенно часто они обнаруживаются в эпителиях, пограничных покровных слоях.
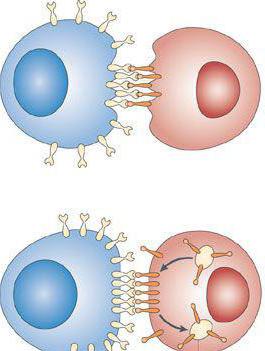
Ученые полагают, что первичное отделение пласта элементов, связанных между собой межклеточными контактами , обеспечило формирование и последующее развитие органов и тканей.
Благодаря использованию методов электронной микроскопии удалось накопить большой объем сведений об ультраструктуре этих связей. Однако их биохимический состав, а также молекулярная структура изучены сегодня недостаточно точно.
Далее рассмотрим особенности, группы и виды межклеточных контактов .
Общие сведения
В образовании межклеточных контактов мембрана участвует очень активно. У многоклеточных за счет взаимодействия элементов формируются сложные клеточные образования. Их сохранение может обеспечиваться разными способами.
В эмбриональных, зародышевых тканях, в особенности на начальных этапах развития, клетки поддерживают связи друг с другом благодаря тому, что их поверхности обладают способностью слипаться. Такая адгезия (соединение) может быть связана со свойствами поверхности элементов.
Специфика возникновения
Исследователи полагают, что образование межклеточных контактов обеспечивается за счет взаимодействия гликокаликса с липопротеидами. При соединении всегда остается небольшая щель (ее ширина порядка 20 нм). В ней находится гликокаликс. При обработке ткани ферментом, способным нарушать его целостность или повреждать мембрану, клетки начинают обосабливаться друг от друга, диссоциируются.
Если удалить диссоциирующий фактор, клетки снова могут собраться вместе. Такое явление называют реагрегацией. Так можно разобщать клетки различных по своей окраске губок: желтых и оранжевых. В ходе экспериментов было установлено, что в соединении клеток возникает только 2 типа агрегатов. Одни состоят исключительно из оранжевых, а другие – только из желтых клеток. Смешанные суспензии, в свою очередь, самоорганизуются и восстанавливают первичную многоклеточную структуру.
Подобные результаты исследователи получили в ходе экспериментов с суспензиями разделенных эмбриональных клеток амфибий. В данном случае клетки эктодермы обосабливаются в пространстве избирательно от мезенхимы и энтодермы. Если для восстановления связей использовать ткани более поздних этапов развития зародышей, в пробирке будут самостоятельно собираться разные клеточные группы, отличающиеся органной и тканевой специфичностью, сформируются эпителиальные агрегаты, имеющие сходство с почечными канальцами.
Физиология: виды межклеточных контактов
Ученые выделяют 2 основные группы связей:
- Простые. Они могут образовывать соединения, различающиеся по форме.
- Сложные. В их число входят щелевидные, десмосомальные, плотные межклеточные контакты , а также адгезивные пояски и синапсы.
Рассмотрим их краткие характеристики.
Простые связи
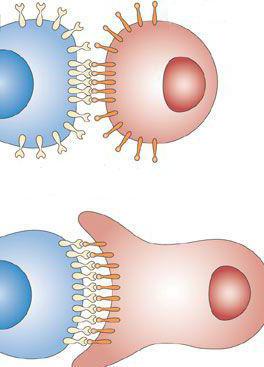
Простые межклеточные контакты представляют собой участки взаимодействия надмембранных клеточных комплексов плазмолеммы. Расстояние между ними не более 15 нм. Межклеточные контакты обеспечивают прилипание элементов за счет взаимного "узнавания". Гликокаликс снабжен специальными рецепторными комплексами. Они строго индивидуальны для каждого отдельного организма.
Образование рецепторных комплексов является специфичным в пределах конкретной популяции клеток или определенных тканей. Они представлены интегринами и кадгеринами, имеющими сродство с аналогичными структурами клеток, находящихся по соседству. При взаимодействии с родственными молекулами, расположенными на прилежащих цитомембранах, происходит их слипание – адгезия.
Межклеточные контакты в гистологии
Среди адгезивных протеинов выделяют:
- Интегрины.
- Иммуноглобулины.
- Селектины.
- Кадгерины.
Некоторые белки, обладающие адгезивными свойствами, не относятся ни к одному из указанных семейств.
Характеристики семейств
Некоторые гликопротеины поверхностного клеточного аппарата относятся к основному комплексу гистосовместимости 1-го класса. Как и интегрины, они строго индивидуальны для отдельного организма и специфичны для тканевых образований, в которых располагаются. Некоторые вещества обнаруживаются только в определенных тканях. К примеру, Е-кадгерины являются специфичными для эпителия.
Интегринами называют интегральные белки, которые состоят из 2-х субъединиц – альфа- и бета-. В настоящее время выявлено 10 вариантов первых и 15 видов вторых. Внутриклеточные участки связываются с тонкими микрофиламентами с помощью специальных белковых молекул (танина или винкулина) либо напрямую с актином.
Селектины представляют собой мономерные белки. Они узнают определенные углеводные комплексы и прикрепляются к ним на поверхности клеток. В настоящее время наиболее изученными являются L, Р и Е-селектины.
Иммуноглобулиноподобные адгезивные белки по своему строению похожи на классические антитела. Некоторые из них являются рецепторами при иммунологических реакциях, другие предназначены только для реализации адгезивных функций.
Межклеточные контакты кадгеринов возникают только при наличии кальциевых ионов. Они задействованы в образовании постоянных связей: Р и Е-кадгерины в эпителиальных тканях, а N-кадгерины – в мышечной и нервной.
Назначение
Следует сказать, что межклеточные контакты предназначены не только для простого сцепления элементов. Они необходимы для обеспечения нормального функционирования тканевых структур и клеток, в формировании которых задействованы. Простые контакты контролируют созревание и перемещение клеток, предупреждают гиперплазию (излишнее увеличение количества структурных элементов).
Разнообразие соединений
В ходе исследований установлены разные типы межклеточных контактов по форме. Они могут быть, к примеру, в виде "черепицы". Такие связи формируются в роговом слое плоского многослойного ороговевающего эпителия, в артериальном эндотелии. Известны также зубчатый и пальцевидный типы. В первом выпячивание одного элемента погружается в вогнутую часть другого. За счет этого существенно увеличивается механическая прочность соединения.
Сложные связи
Эти виды межклеточных контактов специализированы для реализации какой-то определенной функции. Такие соединения представлены небольшими парными специализированными участками плазматических мембран 2-х соседних клеток.
Существуют следующие виды межклеточных контактов:
- Запирающие.
- Сцепляющие.
- Коммуникационные.
Десмосомы
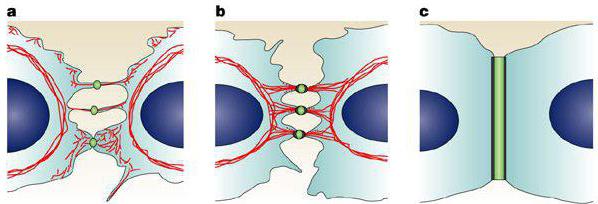
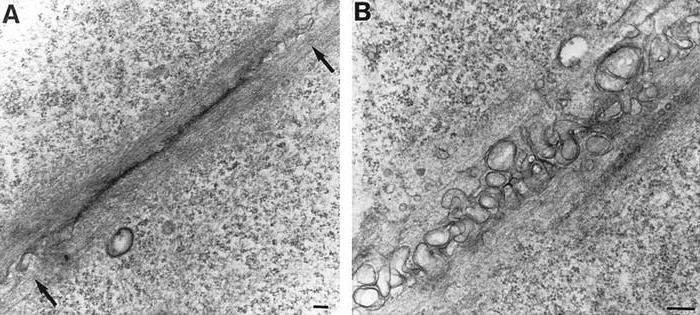
Они являются сложными макромолекулярными образованиями, посредством которых обеспечивается прочное соединение соседних элементов. При электронной микроскопии такой вид контакта заметен очень хорошо, поскольку его отличает высокая электронная плотность. Локальный участок выглядит в форме диска. Его диаметр порядка 0,5 мкм. Мембраны соседних элементов в нем расположены на расстоянии от 30 до 40 нм.
Рассмотреть участки высокой электронной плотности можно и на внутренних мембранных поверхностях обеих взаимодействующих клеток. К ним прикреплены промежуточные филаменты. В ткани эпителия эти элементы представлены тонофиламентами, которые образуют скопления – тонофибриллы. В тонофиламентах присутствуют цитокератины. Между мембранами также обнаруживается электронно-плотная зона, которая соответствует сцеплению протеиновых комплексов соседних клеточных элементов.
Как правило, десмосомы встречаются в эпителиальной ткани, но выявить их можно и в других структурах. В таком случае в промежуточных филаментах содержатся вещества, свойственные этой ткани. К примеру, в соединительных структурах присутствуют виментины, в мышцах – десмины и пр.
Внутренняя часть десмосомы на макромолекулярном уровне представлена десмоплакинами – опорными белками. С ними соединяются промежуточные филаменты. Десмоплакины, в свою очередь, скреплены с десмоглеинами с помощью плакоглобинов. Это тройное соединение проходит сквозь липидный слой. Десмоглеины связываются с белками, находящимися в соседней клетке.
Возможен, однако, и другой вариант. Прикрепление десмоплакинов осуществляется к интегральным белкам, находящимся в мембране, - десмоколинам. Они, в свою очередь, связываются с подобными протеинами соседней цитомембраны.
Поясная десмосома
Она также представлена в виде механического соединения. Однако отличительной ее чертой является форма. Выглядит поясная десмосома в виде ленты. Подобно ободку поясок сцепления охватывает цитолемму и соседние клеточные мембраны.
Этот контакт отличается высокой электронной плотностью и в области мембран, и на участке расположения межклеточного вещества.
В пояске сцепления присутствует винкулин – опорный белок, выступающий в качестве участка прикрепления микрофиламентов к внутренней части цитомембраны.
Адгезивную ленту можно обнаружить в апикальном участке однослойного эпителия. Она часто примыкает к плотному контакту. Отличительной чертой этого соединения является то, что в его структуру входят актиновые микрофиламенты. Они располагаются параллельно относительно поверхности мембраны. За счет их способности сокращаться при наличии минимиозинов и нестабильности целый пласт эпителиальных клеток, а также микрорельеф поверхности органа, которую они выстилают, могут изменять свою форму.
Щелевидный контакт
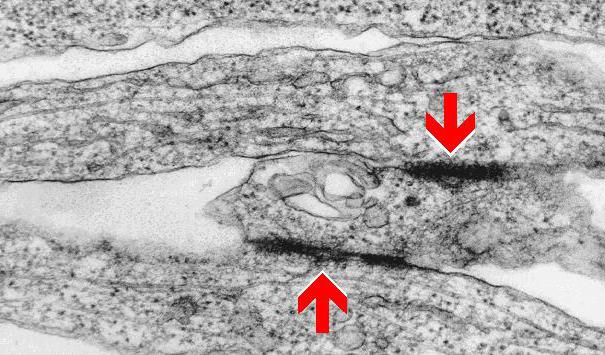
Его называют также нексусом. Как правило, так соединяются эндотелиоциты. Межклеточные контакты щелевидного типа имеют форму диска. Его протяженность составляет 0,5-3 мкм.
На участке соединения соседние мембраны находятся на расстоянии 2-4 нм друг от друга. В поверхности обеих контактирующих элементов присутствуют интегральные протеины – коннектины. Они, в свою очередь, интегрируются в коннексоны – белковые комплексы, состоящие из 6 молекул.
Коннексоновые комплексы прилегают друг к другу. В центральной части каждого расположена пора. Через нее свободно могут проходить элементы, молекулярная масса которых не превышает 2 тыс. Поры в соседних клетках плотно присоединяются друг у другу. За счет этого происходит перемещение молекул неорганических ионов, воды, мономеров, низкомолекулярных биологически активных веществ только в соседнюю клетку, а в межклеточное вещество они не проникают.
Функции нексусов
За счет щелевидных контактов передается возбуждение соседним элементам. К примеру, так проходят импульсы между нейронами, гладкими миоцитами, кардиомиоцитами и пр. За счет нексусов обеспечивается единство биореакций клеток в тканях. В нервных тканевых структурах щелевидные контакты именуются электрическими синапсами.
Задачи нексусов состоят в формировании межклеточного внутритканевого контроля над биоактивностью клеток. Кроме того, такие контакты выполняют несколько специфических функций. К примеру, без них не было бы единства сокращения сердечных кардиомиоцитов, синхронных реакций клеток гладких мышц и пр.
Плотный контакт
Его называют также запирающей зоной. Он представлен в виде участка слияния поверхностных мембранных слоев соседних клеток. Эти зоны формируют непрерывную сеть, которая "сшита" интегральными белковыми молекулами мембран соседних клеточных элементов. Эти белки формируют структуру, похожую на ячеистую сеть. Ею окружен периметр клетки в виде пояска. При этом структура соединяет соседние поверхности.
Часто к плотному контакту прилегают ленточные десмосомы. Этот участок непроницаем для ионов и молекул. Следовательно, он запирает межклеточные щели и, собственно, внутреннюю среду всего организма от внешних факторов.
Значение запирающих зон
Плотный контакт препятствует диффузии соединений. К примеру, содержимое желудочной полости защищено от внутренней среды его стенок, белковые комплексы не могут перемещаться от свободной эпителиальной поверхности в межклеточное пространство и пр. Запирающая зона способствует также поляризации клетки.
Плотные контакты являются основой разнообразных барьеров, присутствующих в организме. При наличии запирающих зон перенос веществ в соседние среды осуществляется исключительно через клетку.
Синапсы
Они представляют собой специализированные соединения, расположенные в нейронах (нервных структурах). За счет них обеспечивается передача информации от одних клеток к другим.
Синаптическое соединение обнаруживается в специализированных участках и между двумя нервными клетками, и между нейроном и другим элементом, включенным в состав эффектора либо рецептора. К примеру, выделяют нервно-эпителиальные, нервно-мышечные синапсы.
Эти контакты разделяют на электрические и химические. Первые аналогичны щелевидным связям.
Сцепление с межклеточным веществом
Клетки присоединяются за счет рецепторов цитолеммы к адгезивным белкам. К примеру, рецепторы к фибронектину и ламинину в клетках эпителия обеспечивают сцепление с этими гликопротеинами. Ламинин и фибронектин являются адгезивными субстратами с фибриллярным элементом базальных мембран (IV тип коллагеновых волокон).
Полудесмосома
Со стороны клетки ее биохимический состав и строение подобен дисмосоме. От клетки в межклеточное вещество отходят особые якорные филаменты. За счет них объединяется мембрана с фибриллярным каркасом и заякоривающие фибриллы коллагеновых волокон VII типа.
Точечный контакт
Его также называют фокальным. Точечный контакт входит в группу сцепляющих соединений. Наиболее характерным он считается для фибробластов. Клетка в таком случае сцепляется не с соседним клеточным элементами, а с межклеточными структурами. Рецепторные протеины взаимодействуют с адгезивными молекулами. К ним относят хондронектин, фибронектин и пр. Они связывают клеточные мембраны с внеклеточными волокнами.
Формирование точечного контакта осуществляется за счет актиновых микрофиламентов. Они закрепляются на внутренней части цитолеммы при помощи интегральных белков.
Плазмолемма многоклеточных организмов активно участвует в образовании специальных структур — межклеточных контактов, или соединений.
Поверхности клеток содержат специальные углеводные части интегральных белков, гликопротеинов, которые специфически связываются с подобными белками на поверхности соседних клеток.
Среди контактов выделяют следующие:
- простые, которые могут формировать различные по форме соединения;
- сложные: десмосомальные, щелевидные, плотные контакты, синапсы и адгезивные пояски.
Формирование рецепторных комплексов специфично в пределах отдельных тканей и популяций клеток. Данные комплексы представлены кадгеринами и интегринами, которые имеют сродство с подобными структурами соседних клеток. Взаимодействуя с родственными молекулами на прилежащих цитомембранах, они прилипают друг к другу — адгезия.
Среди адгезивных белков выделяют несколько семейств: интегрины, селектины, иммуноглобулины (иммуноглобулинподобные белки) и кадгерины. Некоторые адгезивные белки не относятся ни к одному из приведенных семейств.
Часть гликопротеинов поверхностного аппарата клетки относи: к главному комплексу гистосовместимости первого класса (МНС I — major histocompatibility complex I). Они, как и интегрины, строго индивидуальны для каждого организма и специфичны для тканей, в которых находятся. Некоторые из веществ встречаются лишь в определенных тканях, так Е-кадгерины специфичны для эпителия.
Интегрины — это интегральные белки, состоящие из двух (α- и β-) субъединиц. Известно 10 вариантов β- и 15 видов β-субъединиц. Внутриклеточные участки (домены) связаны с тонкими микрофиламентами при помощи специальных белков: винкулина, танина или напрямую с актином.
Селектины — это мономерные белки, которые узнают и прикрепляются к определенным углеводным комплексам на поверхности клеток. Наиболее изучены L-, Р- и Е-селектины.
Адгезивные иммуноглобулинподобные белки по строению напоминают классические антитела. Часть из них выступает в качестве рецепторов при иммунологических реакциях, другая выполняет лишь адгезивные функции.
Кадгерины отличаются тем, что их адгезивная способность проявляется лишь в присутствии ионов кальция. Они участвуют в формировании постоянных клеточных контактов: Е- и Р-кадгерины в эпителии, а N-кадгерины в нервной и мышечной тканях.
Функция простых контактов далеко не ограничена лишь простым механическим сцеплением. Они необходимы для нормального функционирования клеток и тканевых структур, в образовании которых участвуют. Такие контакты контролируют созревание и миграцию клеток, предотвращают излишние митозы (гиперплазию).
Сложные контакты. Они специализированы для выполнения какой-либо функции. Сложные межклеточные контакты представляют собой небольшие парные специализированные участки плазматических мембран двух соседних клеток.
Сложные контакты подразделяют на запирающие (изолирующие), сцепляющие (заякоривающие) и коммуникационные (объединяющие). Имеются сложные контакты, обеспечивающие механическое сцепление клеток: десмосомы и пояски сцеплений (адгезивные пояски, или промежуточные контакты).
Десмосома представляет собой сложное макромолекулярное образование, обеспечивающее прочное сцепление соседних клеток между собой. Контакт хорошо заметен при электронной микроскопии, так как отличается высокой электронной плотностью. Этот локальный участок имеет форму диска диаметром около 0,5 мкм, в котором мембраны соседних клеток располагаются на расстоянии 30…40 нм.
Участки с высокой электронной плотностью видны на внутренних поверхностях мембран обеих взаимодействующих между собой клеток. К этим участкам прикрепляются промежуточные филаменты. В эпителиальной ткани промежуточные филаменты представлены тонофиламентами, образующими скопления — тонофибриллы. Тонофиламснты содержат цитокератины. Имеется также электронно-плотная зона между мембранами, соответствующая сцеплению белковых комплексов соседних клеток.
Чаще всего десмосомы встречаются в эпителии, но они имеются и в других тканях. В этом случае промежуточные филаменты содержат вещества, характерные для этой ткани: виментины в клетках соединительных тканей, десмины в мышцах и т. д.
На макромолекулярном уровне внутренняя поверхность десмосомы представлена опорными белками десмоплакинами. К ним прикрепляются промежуточные филаменты. В свою очередь, десмоплакины соединены с белками десмоглеинами через плакоглобины. Такое тройное соединение пронзает липидный слой мембраны. Десмоглеины, в свою очередь, связаны с подобными белками соседней клетки. Может быть и другой вариант: десмоплакины прикрепляются к интегральным белкам мембраны — десмоколинам, которые, в свою очередь, взаимодействуют с подобными белками соседней цитомембраны.
Поясок сцепления, или поясная десмосома (промежуточный контакт, ленточная десмосома) представляет собой механическое соединение между клетками (подобно десмосоме). В отличие от десмосомы имеет форму ленты. Поясок сцепления как ободок охватывает цитолемму вкупе с соседними клеточными мембранами. Контакт имеет высокую электронную плотность как в области мембран, так и в участке межклеточного вещества, аналогично десмосоме.
Для поясков сцепления характерен опорный белок винкулин, который служит местом прикрепления тонких микрофиламентов к внутренней поверхности цитомембраны. Другие структурные белки и их взаимодействия близки к десмосоме.
Адгезивный поясок встречается в апикальной зоне однослойного эпителия, часто примыкая к плотному контакту. Особенностью данного контакта является то, что к нему прикрепляются актиновые микрофиламенты, которые располагаются параллельно поверхности клеточной мембраны. Их способность сокращаться в присутствии минимиозинов и нестабильность позволяют значительно изменять форму целого пласта эпителиальных клеток и микрорельеф поверхности выстилаемого органа.
Щелевидный контакт, или нексус имеет форму диска протяженностью 0,5…3 мкм. В области щелевидного контакта соседние мембраны приближены друг к другу до 2…4 нм. В мембраны обеих контактирующих клеток встроены интегральные белки коннектины, которые интегрируются в комплексы из шести белков — коннексоны.
Коннексоновые комплексы соседних клеток прилежат друг к другу. В центральной части коннексона находится пора, через которую могут свободно диффундировать ряд молекул молекулярной массой до 2000. Поры соседних клеток плотно присоединены друг к другу, в результате чего перемещающиеся молекулы (неорганические ионы, вода, мономеры, низкомолекулярные биологически активные вещества) попадают только в соседнюю клетку, а не в межклеточное вещество.
Щелевидные контакты позволяют передать возбуждение к соседним клеткам (между нейронами, кардиомиоцитами, гладкими миоцитами и т. д.), через них из одной клетки в другую перемещаются некоторые биологически активные вещества и метаболиты. Нексусы обеспечивают единство биологических реакций клеток в тканях. В нервной ткани щелевидные контакты называют электрическими синапсами.
Значение нексусов заключается в формировании внутритканевого межклеточного контроля над биологической активностью клеток, выполнении ими ряда специфических функций. Без щелевидных контактов невозможны были бы единое сокращение кардиомиоцитов сердца, синхронные реакции гладких мышечных клеток и др.
Нередко к плотному контакту прилежат ленточные десмосомы. Эта область непроницаема для макромолекул и ионов и, следовательно, она запирает межклеточные щели и вместе с ними собственно внутреннюю среду организма от внешней среды. Запирающая зона препятствует диффузии веществ, например содержимого полости желудка от внутренней среды стенки желудка, перемещению белковых комплексов от свободной поверхности эпителия в участки межклеточного пространства; способствует поляризации клетки.
Плотные контакты — это основа многочисленных барьеров организма. При их наличии вещества в соседние среды переносятся только через клетку.
Синапсы — это специализированные контакты, имеющиеся в нервных клетках (нейронах) и обеспечивающие передачу информации от одной клетки к другой. Этот тип соединений встречается в специализированных участках как между двумя нейронами, так и между нейроном и каким-либо иным элементом, входящим в состав рецептора или эффектора, например нервно-мышечные, нервно-эпителиальные синапсы.
Синапсы подразделяют на электрические (аналогичные щелевидным соединениям) и химические.
Адгезивные соединения с межклеточным веществом. Адгезия (прилипание, сцепление) клеток к межклеточному веществу осуществляется через рецепторы цитолеммы к адгезивным белкам. Так, рецепторы к ламинину и фибронектину в эпителиальных клетках обеспечивают связь с этими гликопротеинами. Фибронектин и ламинин — адгезивные субстраты с фибриллярным компонентом базальных мембран (коллагеновыми волокнами IV типа).
Фокальный (точечный) контакт. Его относят к сцепляющим соединениям. Фокальный контакт характерен для фибробластов. В этом случае клетка соединяется не с соседней клеткой, а со структурами межклеточного вещества. Рецепторные белки клетки взаимодействуют с так называемыми адгезивными молекулами (фибронектин, хондронектин и т. д.), связывающими мембраны клетки с внеклеточными волокнами. В образовании фокального контакта участвуют актиновые микрофиламенты, которые прикрепляются к внутренней поверхности цитолеммы с помощью интегральных белков клеточных мембран.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Читайте также:
