
Молекулярные механизмы развития болезни альцгеймера
До недавних пор биохимические механизмы, лежащие в основе почти всех нейродегенеративных болезней с началом во взрослом возрасте, были полностью неясными. Одно из наиболее частых таких заболеваний — болезнь Альцгеймера. Болезнь Альцгеймера обычно проявляется на шестом-девятом десятке лет, но есть моногенные формы, часто дебютирующие раньше, иногда даже на третьем десятилетии жизни.
Клинические проявления болезни Альцгеймера характеризуются прогрессирующим ухудшением памяти и высших корковых функций, например аргументации, а также поведенческими изменениями. Эти аномалии отражают вырождение нейронов в специфических областях коры мозга и гиппокампе.
Болезнь Альцгеймера поражает около 1,4% лиц в развитых странах и вызывает за год только в Соединенных Штатах 100 000 смертей.
Родственники пациентов с болезнью Альцгеймера первой степени родства имеют 38% риска развития болезни к 85-летнему возрасту. Следовательно, оказывается, что большинство случаев с семейным накоплением имеет сложный генетический вклад. Этот вклад может создаваться одним или более независимо действующих неполно пенетрантных генов, несколькими взаимодействующими генами или некоторой комбинацией генетических и средовых факторов.
От 7 до 10% пациентов имеют моногенную высокопенетрантную форму болезни Альцгеймера, наследуемую по аутосомно-доминантному типу. В 1990-е годы обнаружено четыре гена, связанных с болезнью Альцгеймера. Мутации в трех из них, кодирующих бета-АРР, пресенилин 1 и пресенилин 2, ведут к аутосомно-доминантной болезни Альцгеймера. Четвертый ген, АРОЕ, кодирует АроЕ, белковый компонент нескольких плазменных липопротеинов.
Мутации в АРОЕ не связаны с моногенными формами болезни Альцгеймера. Аллель е4 АРОЕ несколько увеличивает восприимчивость к несемейной болезни Альцгеймера и влияет на возраст начала, по крайней мере, при некоторых моногенных формах.

Наиболее важные патологические аномалии при болезни Альцгеймера — накопление в мозге двух фибриллярных белков, А-бета и белка тау. Пептид А-бета образуется из большего белка бета-АРР и обнаруживается во внеклеточном амилоиде или сенильных бляшках во внеклеточном пространстве мозга при болезни Альцгеймера.
Амилоидные бляшки, кроме пептида А-бета, содержат и другие белки, особенно АроЕ. Тау-белок — микротубулярный, обильно экспрессирующий в нейронах мозга. Гиперфосфорилированные формы тау-белка формируют нейрофибриллярные клубки, обнаруживаемые при болезни Альцгеймера, в отличие от внеклеточных амилоидных бляшек, внутри нейронов.
Тау-белок в норме обеспечивает сборку и устойчивость микротрубочек, эта функция уменьшается при фосфорилировании. Хотя образование клубков нейрофибрилл оказалось одной из причин гибели нейронов при болезни Альцгеймера, мутации в гене тау-белка связаны не с болезнью Альцгеймера, а с другим аутосомно-доминантным заболеванием, лобновисочной деменцией.
Бета-АРР — трансмембранный белок, подвергающийся трем различным видам протеолиза, в зависимости от относительной активности трех разных протеаз: а- и бета-секретаз — поверхностных клеточных протеаз; и у-секретазы — атипичной протеазы, расщепляющей мембранные белки в трансмембранных областях. Преобладающая судьба приблизительно 90% бета-АРР — расщепление а-секретазой, что предотвращает образование А-бета-пептида, так как а-секретаза расщепляет белок внутри него.
Оставшиеся приблизительно 10% бета-АРР расщепляются бета- и у-секретазами, формируя или нетоксичный пептид А-бета-40, или пептид А-бета-42, обладающий нейротоксичностью. Пептид А-бета-42 считают нейротоксичным, поскольку он более склонен к формированию нейрофибрилл, чем его аналог А-бета-40, признак, делающий болезнь Альцгеймера конформационной болезнью, подобно недостаточности а1-антитрипсина.
В норме образуется небольшое количество пептида А-бета-42; факторы, определяющие, будет ли белок расщепляться у-секретазой с образованием А-бета-40 или А-бета-42, не определены. При моногенной болезни Альцгеймера вследствие миссенс-замен в гене, кодирующем бета-АРР, тем не менее, несколько мутаций в гене бета-АРР избирательно увеличивают образование пептида А-бета-42. Это увеличение приводит к накоплению нейротоксичного А-бета-42 — основе патогенеза всех форм болезни Алыдгеймера, как моногенных так и спорадических.
Эта модель подтверждается тем, что пациенты с синдрома Дауна, имеющие три копии гена бета-АРР (расположенного в хромосоме 21), обычно имеют нейропатологические изменения болезни Альцгеймера уже в 40-летнем возрасте. Кроме того, мутации в генах пресенилина 1 и 2 также ведут к повышенному образованию А-бета-42. Примечательно, что в сыворотке больных с мутациями в генах бета-АРР, пресенилина 1 и 2 количество нейротоксичного пептида А-бета-42 повышается, и в культивируемых клетках экспрессия мутантных генов бета-АРР, пресенилина 1 и 2 увеличивает относительное образование пептида А-бета-42 в 2-10 раз.
Гены, кодирующие пресенилин 1 и пресенилин 2, обнаружены стратегией позиционного клонирования в семьях с аутосомно-доминантной формой болезни Альцгеймера. Пресенилин 1 необходим для расщепления у-секретазой производных бета-АРР. На самом деле, существуют подтверждения того, что пресенилин 1 — важный белковый кофактор у-секретазы.
Мутации в пресенилине 1 связаны с болезнью Альцгеймера через до сих пор неясный механизм, увеличивающий образование пептида А-бета-42. Белок пресенилин 2 имеет на 60% идентичную последовательность аминокислот с пресенилином 1, что указывает на их общие функции. Основное различие между мутациями в гене пресенилина 1 и 2 в том, что возраст начала во втором случае более вариабелен (пресенилин 1 — от 35 до 60 лет; пресенилин 2 — от 40 до 85 лет), в одной семье бессимптомный восьмидесятилетний носитель мутации в гене пресенилина 2 передал болезнь своему потомству. Эта разница частично зависит от числа е4 аллелей АРОЕ у носителей мутации в гене пресенилина 2; два е4 аллеля приводят к более раннему возрасту начала, чем один аллель, также обусловливающий более раннее начало по сравнению с другими аллелями АРОЕ.
Один аллель гена АРОЕ, е4 аллель, — основной фактор риска развития болезни Альцгеймера. Роль АРОЕ как основного локуса восприимчивости к болезни Альцгеймера была доказана четырьмя независимыми способами: анализом сцепления в семьях с накоплением болезни Альцгеймера с поздним началом, сильной ассоциацией аллеля е4 с болезнью Альцгеймера по сравнению с группой контроля, открытием того, что белок АроЕ — компонент амилоидных бляшек при болезни Альцгеймера, и обнаружением факта, что АроЕ связан с пептидом А-бета.
Белок АроЕ имеет три частых формы, кодируемые соответствующими аллелями АРОЕ. Аллель е4 значительно преобладает среди пациентов с болезнью Альцгеймера (40% по сравнению с 15% в общей популяции) и связан с ранним началом болезни (для гомозигот по аллелю е4 возраст начала болезни Альцгеймера на 10-15 лет меньше, чем в общей популяции). Кроме того, отношение между аллелем е4 и болезнью дозозависимое; две копии е4 связаны с более ранним началом (средний возраст начала до 70 лет), чем одна копия (средний возраст начала после 70 лет). В отличие от этого, е2 аллель имеет защитный эффект и соответственно чаще встречается у пожилых, незатронутых болезнью Альцгеймера.
Механизмы, лежащие в основе данных эффектов, неизвестны, но полиморфные варианты АроЕ могут влиять на процессинг бета-АРР и плотность амилоидных отложений в мозге пациентов с болезнью Альцгеймера. Например, мыши без АроЕ имеют выраженное снижение депонирования пептида А-бета, получаемого из мутантного гена бета-АРР, связанного с семейной формой болезни Альцгеймера. Предполагают и другие механизмы, например измененный ответ на повреждение, так как ген АРОЕ управляется в мозге в процессе повреждения и восстановления. Важно иметь в виду, что е4 аллель АроЕ неоднозначно связан с повышенным риском болезни Альцгеймера. Таким образом, носители аллелей е4 имеют плохие неврологические результаты после черепно-мозговых травм, инсультов и других неврологических нарушений.
Хотя носители е4 аллеля АРОЕ имеют четко повышенный риск развития болезни Альцгеймера, к настоящему времени скрининг на присутствие этого аллеля у здоровых индивидуумов нецелесообразен; такое тестирование имеет высокие цифры ложноположительных и ложноотрицательных ответов и приводит к неопределенным оценкам риска болезни Альцгеймера.
Другие гены болезни Альцгеймера. Статистический анализ показывает, что риск болезни Альцгеймера могут значительно изменять еще 4-8 генов. Сущность их неясна. Кроме того, исследования типа случай-контроль при болезни Альцгеймера указывают на длинный список возможных генов (>100), но лишь несколько из них получили подтверждение при повторном анализе, и их роль в генетическом определении риска при болезни Альцгеймера остается неизвестной.
МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ БОЛЕЗНИ АЛЬЦГЕЙМЕРА: ГЕНЕТИЧЕСКИЙ ПОДХОД
Е. И. Рогаев Научный центр психического здоровья РАМН, Москва
Центральной задачей при исследовании патогенеза сложного заболевания является идентификация первичных молекулярных нарушений.
Предполагается, что первые гистопатологические изменения (например, накопление амилоидных бляшек) при болезни Альцгеймера могут возникать задолго (за 10-20 лет) до появления первых клинических симптомов. Это, разумеется, затрудняет идентификацию первичных дефектов, ведущих к нейродегенерации. Прорывом в данной области явился генетический подход, в частности, стратегия позиционного клонирования генов.
Генетическая предрасположенность и возраст являются наиболее четко определенными факторами риска развития болезни Альцгеймера (БА).
1. Роль генетических факторов при болезни Альцгеймера
Еще в 40-х годах была предсказано, что генетические факторы играют значительную роль в патогенезе болезни Альцгеймера как с ранним, так и с поздним началом болезни. Суммируя данные близнецового анализа, характера наследования БА в семьях и недавние результаты анализа генов, вовлеченных в болезнь Альцгеймера, можно сделать следующие выводы:
1) семейные формы с ранним началом БА (до 65, в среднем 35-55 лет) характеризуются аутосомно-доминантным наследованием. Необходимой и достаточной причиной патологии при этом является мутация в единственном гене. Число больных с такими генетическими формами БА, видимо, не превышает 10 % от всех случаев БА.
Обнаружение и клонирование генов БА, в целом, подтверждают данную схему.
2. Гены, мутации в которых ведут к развитию болезни Альцгеймера
К настоящему времени выявлено 4 гена, мутации или вариации, которые бесспорно вызывают или являются факторами риска БА.
Анализ многих популяций различного этнического происхождения, в том числе русских (см. в данном сборнике), показал, что полиморфизм в гене аполипопротеина АРОЕ (аллель е4, хромосома 19) является наиболее широко распространенным фактором риска как ранних, так и поздних форм БА. Следует отметить, что в отличие от мутации в гене PS 1 наличие в генотипе индивида е4 варианта не является необходимым или достаточным условием развития БА.
Несколько мутаций в экзоне 16 и 17 у пациентов с ранней семейной БА были ранее описаны для гена предшественника амилоида (APP, хромосома 21).
Суммируя, можно сказать, что мутации в генах пресенилина 1 и 2 ответственны за 30-70 % (по подсчетам разных авторов), а мутации в гене APP менее 5 % случаев семейных форм БА с ранним началом. АРОЕ е4 является одним из факторов риска в 30-50 % всех случаев БА.
3. Функции генов болезни Альцгеймера в норме и патологии. Биологическая модель
На первый взгляд, не существует очевидной функциональной или структурной связи между белковыми продуктами тех генов, мутации в которых ассоциированы с БА. Пресенилины — внутриклеточные белки со множественными трансмембранными доменами. APP — белок с одним трансмембранным доменом, локализованным на плазматической мембране. Их экспрессия происходит во всех тканях, в том числе и в нейронах. Аполипопротеин Е — белок со множественными функциями, экспрессируется в мозге, но в глиальных клетках, а не в нейронах.
Исследования клеточных моделей и трансгенных животных показывают, что мутантные формы данных генов изменяют:
1) процессинг APP и аккумуляцию наиболее нейротоксичных форм бета-амилоида;
2) вызывают повышение чувствительности клеток различного типа, в том числе нейронов, к апоптозу;
3) могут влиять на изменение внутриклеточного Са 2+ -гомеостаза и различные системы сигнальной трансдукции и окислительного стресса.
Аполипопротеин Е участвует также в процессах регенерации ЦНС при повреждениях и является фактором риска (е4 форма) при сердечно-сосудистых заболеваниях.
Для выявления непосредственных молекулярных элементов, взаимодействующих с продуктами пресенилинов, APP или АРОЕ, необходимо дальнейшее развитие подходящих клеточных моделей или моделей трансгенных животных. Помимо традиционной модели трансгенной мыши (см. Григоренко и др. в данном сборнике) мы начали создание принципиально новых моделей беспозвоночных (дрозофила и моллюск) для исследования генов БА, и в частности, пресенилинов. В настоящее время клонированы ортологичные гены пресенилинов из Drosophila melanogaster (в сотрудничестве с Университетом г. Торонто) и из улитки Helix L . (неопубликованные данные). Данные модели, как предполагается, будут наиболее подходящими для идентификации элементов сигнальной трансдукции, взаимодействующих с пресенилинами.
Перспективы генной диагностики и терапии
Для проведения генной диагностики БА необходимо: выяснение распространенности мутаций (например, в генах пресенилинов) не только в семейных, но и в спорадических случаях БА и других деменций в разных популяциях; поиск новых генов (например, на хромосоме 12), полиморфизмы в которых являются факторами риска БА или протектирующими факторами при БА.
Уже в настоящее время представляется перспективным тестирование на мутации гена пресенилина 1 у пациентов и родственников в семьях с ранним началом БА. Обнаружение мутации в эволюционно-консервативном сайте пресенилина 1 позволяет, по существу, однозначно предсказать развитие болезни до 60 лет (и наиболее вероятно в возрасте 35-55 лет).
Модели клеточных линий и животных, содержащих трансгенные мутантные формы генов БА, позволят тестировать различные лекарства и молекулы, влияющие на клеточные механизмы
1) процессинга амилоида;
2) апоптоза (програмированной клеточной смерти)
3) сигнальной трансдукции в данных системах.
Это дает надежды на создание лекарственных препаратов, имеющих не симптоматический эффект, а влияющих на первичные молекулярные нарушения при БА.
Главным представителем нейродегенеративных заболеваний с пораженим коры больших полушарий является болезнь Альцгеймера, которая проявляется деменцией — прогрессирующей утратой когнитивных функций вне зависимости от напряженности внимания. Существует несколько видов деменции: фронтотемпоральная деменция, сосудистая деменция, деменция с тельцами Леви, болезнь Крейтцфельда-Якоба и нейросифилис. При этих заболеваниях также поражаются субкортикальные структуры, но многие симптомы связаны с изменениями в коре больших полушарий. Независимо от этиологии деменция не является нормальным процессом старения, а всегда служит проявлением патологии.
Болезнь Альцгеймера — самая частая причина деменции в пожилом возрасте. Заболевание проявляется постепенными нарушениями интеллекта и расстройствами в эмоциональной сфере и поведении. По мере прогрессирования поражения коры присоединяются дезориентация, мнестические нарушения и афазия. В течение 5-10 лет пациент становится инвалидом, недоступным для контакта и лишенным способности к самостоятельному передвижению.
Симптомы редко возникают до 50 лет, однако частота заболевания с возрастом увеличивается в 2 раза каждые 5 лет: в возрастной группе от 60 до 64 лет частота составляет 1 %, в возрасте от 85 до 89 лет — 40% и выше. Прогрессирующая частота заболевания в связи с возрастом создала множество медицинских, социальных и экономических проблем в странах со стареющим населением. Большинство случаев болезни являются спорадическими, на долю семейной формы приходится лишь 5-10%. Тем не менее изучение именно семейного заболевания пролило свет на патогенез спорадической формы.
Хотя морфологическое исследование ткани мозга по-прежнему необходимо для постановки достоверного диагноза, современная клинико-рентгенологическая диагностика позволяет определить наличие болезни Альцгеймера в 80-90% случаев.
а) Морфология. При аутопсийном исследовании головного мозга видна атрофия коры различной степени выраженности с расширением борозд. Эти изменения особенно заметны в лобной, височной и теменной долях. Из-за выраженной атрофии происходит расширение желудочковой системы (гидроцефалия ex vacuo), поскольку объем мозговой ткани уменьшается. Поражение медиальных отделов височной доли, в т.ч. гиппокампа, энторинальной коры и миндалевидного комплекса, происходит раньше всего, поэтому на поздних стадиях болезни атрофические изменения этих структур выражены особенно сильно.
Главной характеристикой болезни Альцгеймера, позволяющей поставить гистологический диагноз при микроскопическом исследовании, являются сенильные бляшки и сети нейрофибрилл. Наблюдаются прогрессирующая и впоследствии массовая гибель нейронов и реактивный глиоз в местах образования сенильных бляшек и сетей нейрофибрилл.
Сенильные (нейритные) бляшки представляют собой фокальные сферические скопления расширенных и извитых отростков нейронов (дистрофически измененных отростков), расположенных обычно вокруг амилоидного центра, который может быть окружен светлым ободком. Диаметр сенильных бляшек варьирует от 20 до 200 мкм, по периферии располагаются клетки микроглии и реактивные астроциты. Сенильные бляшки обнаруживаются в гиппокампе, миндалевидном комплексе и неокортексе, однако поражение первичной двигательной и соматосенсорной коры выражено в меньшей степени (что также справедливо и для сетей нейрофибрилл).
Амилоидный центр, который можно выявить при окрашивании конго красным, содержит патологический белок. Главный компонент амилоидного центра сенильных бляшек — белок Ар, производное более крупной молекулы — АРР. Аминокислотные последовательности двух основных разновидностей белка Aβ, обозначаемых Aβ40 и Aβ42, имеют одинаковый N-конец и отличающийся на 2 аминокислотных остатка С-конец. Другие белки, присутствующие в сенильных бляшках, малочисленны, к ним относятся компоненты системы комплемента, провоспалительные цитокины, а1-антихимотрипсин и аполипопротеины.

Болезнь Альцгеймера.
Атрофия коры больше выражена в правом полушарии,
где удален лептоменинкс.
В ряде случаев происходит отложение белка Аβ со всеми тинкториальными характеристиками амилоида в отсутствие окружающей нейрональной реакции. Эти образования, называемые диффузными сенильными бляшками, обнаруживаются в поверхностных слоях коры больших полушарий, а также в базальных ядрах и коре мозжечка. Диффузное поражение является маркером ранней стадии формирования зрелых сенильных бляшек. Это заключение было сделано при исследовании мозга лиц с трисомией по 21-й хромосоме (синдромом Дауна). Для лиц с синдромом Дауна характерно более раннее начало болезни Альцгеймера.
В некоторых участках мозга (коре мозжечка и стриатуме) диффузные сенильные бляшки сами по себе или наряду с другими изменениями отражают пик развития болезни Альцгеймера. Сенильные бляшки содержат и Aβ40, и Aβ42, а диффузные сенильные бляшки состоят преимущественно из Aβ42.
Сети нейрофибрилл — это пучки филаментов в цитоплазме нейронов, смещающие или окружающие ядро. В пирамидных клетках пучки филаментов внешне часто напоминают рисунок языков пламени; в клетках более округлой формы переплетения волокон вокруг ядра формируют шаровидные сети нейрофибрилл. Эти сети имеют базофильную окраску при окрашивании гематоксилином и эозином и очень хорошо импрегнируются серебром. Сети нейрофибрилл обнаруживаются, как правило, в корковых нейронах, особенно в энторинальной коре, но могут иметь и другую локализацию.
Другими компонентами являются МАР2 (белок, связанный с системой микротрубочек) и убиквитин. Парные спиралевидные филаменты обнаруживаются также в дистрофически измененных отростках нейронов, которые образуют наружные слои сенильных бляшек, и в аксонах, проходящих через пораженное серое вещество (нити нейропиля). Сети нейрофибрилл выявляются при других заболеваниях, т.е. не специфичны для болезни Альцгеймера.
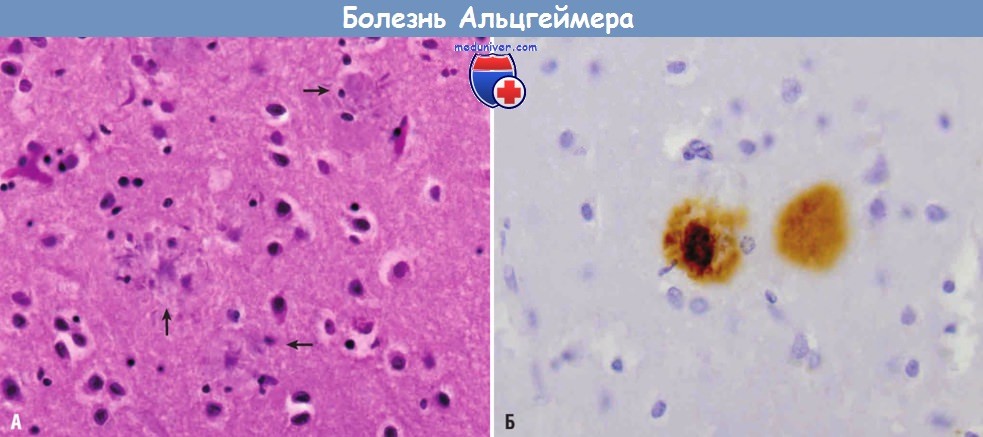
Болезнь Альцгеймера:
(А) Сенильные бляшки с дистрофическими отростками нейронов вокруг амилоидного центра (стрелки).
(Б) В центре сенильной бляшки и окружающем нейропиле с помощью иммуногистохимического исследования выявляется белок Aβ.
Для болезни Альцгеймера характерны и другие патоморфологические изменения. Церебральная амилоидная ангиопатия — неизменный спутник болезни Альцгеймера, но встречается и у лиц, не страдающих болезнью Альцгеймера. При церебральной амилоидной ангиопатии в сосудах откладывается преимущественно Aβ40. Гранулярно-вакуольная дегенерация представляет собой образование мелких (5 мкм в диаметре) светлых вакуолей в цитоплазме нейронов, каждая их которых содержит аргирофильные гранулы.
Вакуоли могут возникать при нормальном процессе старения, но при болезни Альцгеймера такие изменения выявляются преимущественно в гиппокампе и обонятельных луковицах. Тельца Хирано, очень характерные для болезни Альцгеймера, представляют собой удлиненные стекловидные эозинофильные структуры, состоящие из рядов, построенных из паракристаллиновых филаментов, основным компонентом которых является актин. Тельца Хирано обнаруживаются в основном в пирамидных нейронах гиппокампа.
Поскольку изредка сенильные бляшки и сети нейрофибрилл могут быть у лиц, не страдающих деменцией, окончательный диагноз ставят на основании клинических и патоморфологических данных. Прогрессирование болезни идет непрерывно. Патоморфологические изменения (появление сенильных бляшек, сетей нейрофибрилл, гибель нейронов и глиальная реакция) раньше всего появляются в энторинальной коре, затем распространяются через гиппокампальную формацию на мезокортекс и, наконец, достигают неокортекса. Сенильные бляшки оценивают в каждом участке коры (отсутствуют, мало, умеренное количество, много), а сети нейрофибрилл описывают по распространенности в головном мозге.
По этим характеристикам в сочетании с критериями NIA-Reagan устанавливают причастность патоморфологических изменений, типичных для болезни Альцгеймера, к развитию деменции у данного пациента.
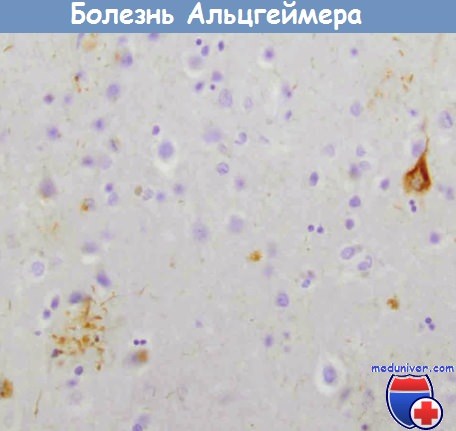
Болезнь Альцгеймера.
Сеть нейрофибрилл (вверхуслева) и нейриты вокруг сенильной бляшки (внизу справа),
содержащей тау-белок (иммуногистохимическое исследование). 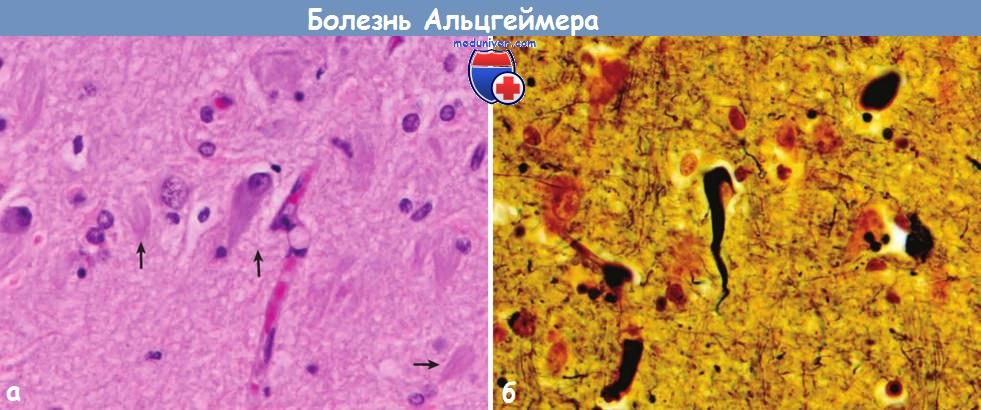
Болезнь Альцгеймера:
(А) Внутри нейронов видны сети нейрофибрилл, которые присутствуют и во внеклеточном пространстве (стрелки).
(Б) При импрегнации серебром видны сети нейрофибрилл в цитоплазме нейронов.
б) Молекулярная генетика и патогенез. В основе развития болезни Альцгеймера лежит отложение белков Aβ, которые образуются в результате процессинга АРР. АРР — это поверхностный белок, содержащий один трансмембранный домен. АРР может выполнять функцию рецептора, хотя лиганды этого белка до сих пор не обнаружены. Фрагмент Aβ АРР простирается от внеклеточного пространства до трансмембранного домена. Процессинг АРР начинается с расщепления внеклеточной части домена, затем расщепляется внутримембранная часть домена. Существует два пути, которые различаются по начальной стадии протеолиза. Если первое расщепление происходит в сайте связывания а-секретазы на участке последовательности, соответствующей Aβ, то белок Aβ не образуется (неамилоидогенный путь).
Такой процессинг АРР обычно происходит на поверхности клетки, где различные ферменты с а-секретазной активностью расщепляют поверхностные белки. Поверхностный АРР также может быть подвергнут эндоцитозу, тогда его расщепление происходит с помощью b-секретазы, которая разрывает цепочку аминокислот у N-конца Aβ (амилоидогенный путь). После расщепления АРР в одном из сайтов, у-секретаза осуществляет расщепление внутримембранной части домена. Если первый этап был выполнен а-секретазой, то образуется растворимый фрагмент, если b-секретазой — продуктом расщепления является белок Aβ. Различия в длине белков (Aβ40 vs Aβ42) обусловлены вариабельностью места разрыва аминокислотной цепочки при ее расщеплении у-секретазой. у-Секретазный комплекс, содержащий пресенилин, никастрин, pen-2 и aph-1, также участвует в процессинге сигнальной молекулы Notch, которая наряду с множеством других мембранных белков определяет судьбу клетки.
Белок Aβ имеет очень большую склонность образовывать агрегаты — сначала олигомеры (токсичные для нейронов), затем крупные агрегаты и фибриллы.
Точечные мутации гена, кодирующего АРР, тоже могут приводить к появлению семейной формы болезни Альцгеймера. Сайты некоторых мутаций находятся вблизи сайтов связывания b-секретазы и у-секретазы, другие локализуются в пределах последовательности Aβ АРР и усиливают способность белка к агрегации. Два локуса, ответственные за развитие большинства случаев семейной формы болезни Альцгеймера с ранним началом, кодируют два пресенилина: PS1 на 14-й хромосоме и PS2 на 1-й хромосоме. Мутации в этих локусах приводят к появлению патологических функций, например усиленной выработки у-секретазой белка Aβ, особенно Aβ42. Таким образом, генетические данные доказывают ведущую роль белка Aβ в патогенезе болезни Альцгеймера.
Белки Aβ сразу образуют агрегаты, обладающие нейротоксичностью. Есть свидетельства, что мелкие агрегаты вызывают дисфункцию синапсов путем долговременного блокирования передачи синаптических сигналов и изменений свойств других мембран других клеток. Агрегаты очень плохо распадаются, но мономерный Aβ может быть лизирован протеазами. И мелкие, и крупные агрегаты вызывают воспалительную реакцию со стороны микроглии и астроцитов. Не исключено, что эта реакция способствует удалению агрегатов, однако в то же время индуцирует секрецию повреждающих медиаторов. Другими следствиями активации каскада воспалительных реакций являются изменения процесса фосфорилирования и окислительное повреждение нейронов.
Генный локус на 19-й хромосоме кодирует апоЕ, который влияет на риск развития болезни Альцгеймера. Из-за полиморфизма двух аминокислот существуют 3 аллеля белка апоЕ (е2, е3 и е4). Увеличенная порция аллеля е4 повышает риск болезни Альцгеймера и снижает возраст манифестации заболевания. Установлено, что среди всех пациентов с болезнью Альцгеймера носители этого аллеля составляют абсолютное большинство. Аллель е4 вызывает образование белка Aβ и отложение его агрегатов по неизвестному механизму. В целом этот аллель ответственен за 25% случаев спорадической формы болезни Альцгеймера. Вероятно, другие аллели тоже являются факторами риска, но со значительно меньшим влиянием в популяции. В выявлении этих, более слабых локусов может быть полезен внедряемый новый подход к геномному скринингу.
Известно, что сети нейрофибрилл содержат тау-белок, поэтому его роль в развитии болезни Альцгеймера представляет большой интерес. Tay-белок связан с системой микротрубочек аксонов. При образовании сетей нейрофибрилл на фоне болезни Альцгеймера этот белок перемещается в тела и дендриты нейронов, где происходит его гиперфосфорилирование и он утрачивает способность к связыванию с микротрубочками.
Считается, что главным триггером развития болезни Альцгеймера является образование патологической формы белка Aβ, а не влияние тау-белка, поскольку мутации гена, кодирующего белок Aβ, приводят к формированию сетей нейрофибрилл и развитию болезни Альцгеймера, а мутации гена МАРТ, кодирующего тау-белок, вызывают развитие одной из форм фронтотемпоральной деменции, но не накопление белка Aβ. Механизм повреждения нейронов сетями нейрофибрилл мало изучен.
Вопрос о морфологическом субстрате деменции у пациентов с болезнью Альцгеймера остается открытым, однако доказана тесная взаимосвязь широкой распространенности сенильных бляшек и сетей нейрофибрилл с грубыми когнитивными нарушениями, причем количество сетей нейрофибрилл сильнее коррелирует со степенью выраженности деменции, чем количество сенильных бляшек. К биохимическим маркерам, коррелирующим с выраженностью деменции, относятся дефицит холинацетилтрансферазы, иммунореактивность синаптофизина и распространенность отложений амилоида.
в) Клинические признаки. Прогрессирование болезни Альцгеймера — длительный и непрерывный процесс (длительность симптоматического течения часто превышает 10 лет). Начальными симптомами являются мнестические нарушения, по мере развертывания клинической картины заболевания присоединяются речевые расстройства, акалькулия, утрата приобретенного динамического праксиса. Для терминальной стадии болезни Альцгеймера характерны нарушения контроля функции тазовых органов по типу недержания, мутизм и невозможность самостоятельного передвижения. Часто наблюдаются интеркуррентные заболевания (прежде всего пневмония), которые, как правило, приводят к смерти пациента. Одним из важнейших направлений исследований болезни Альцгеймера является выявление специфических биомаркеров. В настоящее время используют позитронно-эмиссионную томографию с амилоидсвязывающим препаратом PiB.
Механизмы процессинга белка-предшественника амилоида.
Существует 2 пути процессинга: неамилоидогенный (расщепление с помощью β-секретазы и y-секретазы) и амилоидогенный (путь, который приводит к образованию агрегатов белка Аβ и амилоидных фибрилл).
Читайте также:
