
Молекулярный слой нервных клеток
Толщина коры больших полушарий (от греч. pallium — мантия) варьирует от 2 до 4 мм, наиболее тонкая ее часть находится в области первичной зрительной коры, а наиболее утолщенная — в первичной моторной области. Больше половины общей поверхности коры скрыто стенками борозд. Мозг содержит приблизительно 86 млрд, нейронов (кора больших полушарий содержит только 19% общего количества, но составляет 81% массы мозга), аналогичное число глиальных клеток и разветвленную капиллярную сеть.
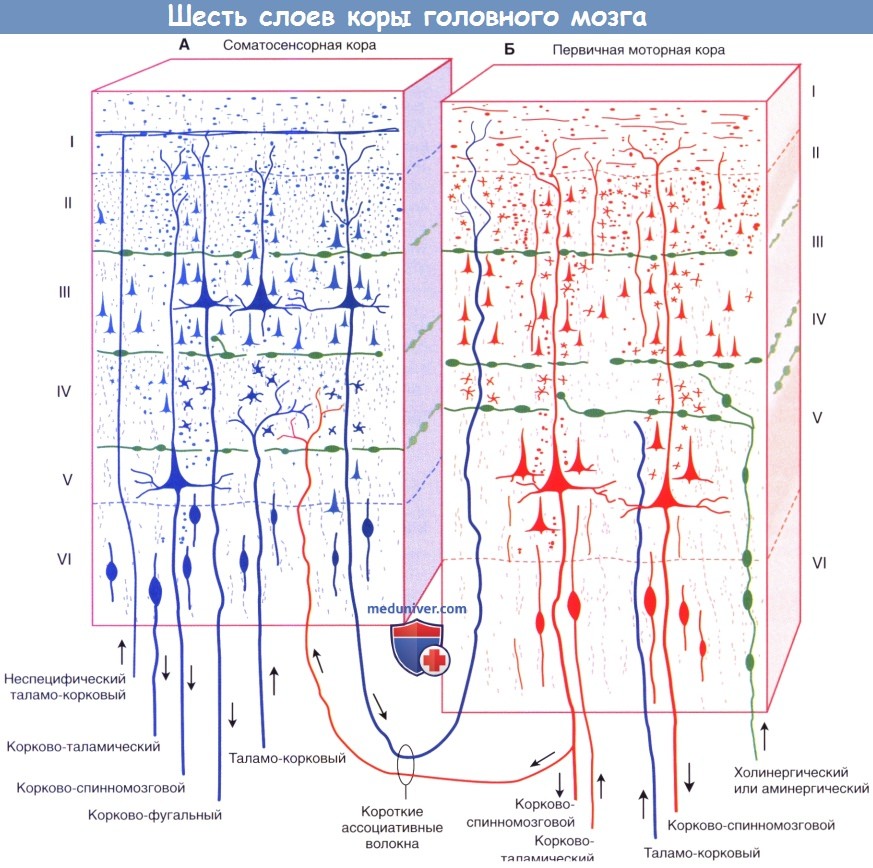
Шестислойное строение изокортекса коры головного мозга.
(А) Соматосенсорная кора. (Б) Первичная моторная кора, кортикальные пластинки обозначены цифрами от I до VI.
Клеточные пластинки новой коры (неокортекса):
I. Молекулярный слой образован концами апикальных дендритов пирамидных клеток и наиболее дистальными ветвями аксонов, проходящих к коре от интраламинарных ядер таламуса.
II. Наружный зернистый слой образован мелкими пирамидными и звездчатыми клетками.
III. Наружный пирамидный слой состоит из средних пирамидных клеток и звездчатых клеток.
IV. Внутренний зернистый слой образован звездчатыми клетками, к которым подходят восходящие волокна от релейных таламических ядер. Звездчатые (зернистые) клетки особенно многочисленны в первичной соматосенсорной коре, первичной зрительной коре и первичной слуховой коре; к ним подходят афферентные чувствительные волокна. Эти отделы обозначают как гранулярная кора. Наоборот, первичная моторная кора — место начала корково-спинномозгового и корково-бульбарного путей, имеет относительно небольшое количество зернистых клеток в слое IV и большое число пирамидных клеток в слоях III и V, которые смазывают (перекрывают) отдельные слои. Эту область обозначают как агранулярная кора.
V. Внутренний пирамидный слой образован крупными пирамидными клетками, волокна которых направляются к полосатому телу, стволу мозга и спинному мозгу.
VI. Слой веретеновидных клеток образован видоизмененными пирамидными клетками, волокна которых направляются к таламусу.
Взаимодействие между группами колонн позволяет осуществлять более сложную деятельность, модели поведения или решать когнитивные задачи.
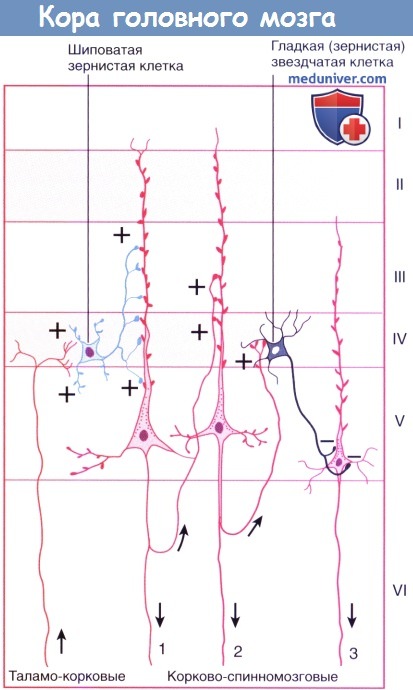
Восходящие/нисходящие проводящие пути. Стрелки указывают направление проведения импульса. Знаки +/- обозначают возбуждение/торможение.
Пирамидная клетка 1 возбуждается шиповатой зернистой клеткой; она возбуждает клетку 2 внутри своей колонны клеток; клетка 3 в соседней колонне тормозится гладкой зернистой клеткой.
в) Типы клеток. Морфологически кортикальные нейроны разделяют на две большие группы. Большая часть (60-85%) представлена пирамидными нейронами (из-за их формы) — единственными, волокна которых покидают кору (и к которым подходит большая часть волокон), что объясняет их альтернативное название — корковые проекционные нейроны; их волокна — возбуждающие глутаматергические. К оставшимся 15-40% нейронам относят непирамидные, или вставочные, нейроны; несмотря на то, их взаимосвязи не выходят за пределы коры, они осуществляют регуляцию и значительное влияние на ее деятельность; их тип — преимущественно тормозной ГАМК-ергический.
Внутри каждой группы можно выделить множество подгрупп в зависимости от морфологии, взаимосвязей, электрофизиологических свойств, типа развития, физиологических характеристик, молекулярных маркеров и т. д. (Примерами основных морфологических и функциональных клеточных типов служат пирамидные клетки, шиповатые зернистые клетки (измененные пирамидные клетки) и группа непирамидных тормозных вставочных нейронов.)
• Шиповатые зернистые клетки — один из вариантов атипичных пирамидных клеток, лежащих в слое IV и наиболее многочисленных в первичной сенсорной коре. Их шиповатые дендриты не выходят за пределы слоя IV, а аксоны могут подниматься или опускаться, образуя возбуждающие глутаматергические синаптические контакты с пирамидными клетками. На них переключается большая часть волокон восходящих таламических путей к слою IV с дальнейшим радиальным распространением.
Для организации их можно разделить на три крупных семейства в зависимости от экспрессируемых этими вставочными нейронами биомаркеров: парвальбумин, соматостатин и серотонин (5-гидрокситриптамин, 5НТ) 3а-рецептор (5HT3aR).
• Парвальбумин-экспрессирующие вставочные нейроны не имеют шигшков на дендритах. К ним подходят возбуждающие волокна от таламуса и коры, а тормозные — от других вставочных нейронов того же типа. Считают, что они играют роль в стабилизации активности корковых нейронных сетей. Как и в коре мозжечка, эти нейроны обладают фокусирующим действием на кору больших полушарий, тормозя слабо возбужденные колонны клеток. Клетки-канделябры (названы так из-за канделяброподобных групп аксоаксональных синаптических окончаний) наиболее многочисленны в слое II, образуют контакты с начальным сегментом аксона пирамидной клетки и играют ключевую роль в корково-корковых взаимодействиях.
• Соматостатин-экспрессирующие вставочные нейроны. Примером служат клетки Мартинотти, расположенные в пластинках V и VI, аксоны которых направляются к пластинке I. Получая сигналы от пирамидных клеток, они способны ограничивать их периферическое возбуждение и объединять несенсорную информацию, осуществляя, таким образом, регулирование обработки сигналов в дендритах их пирамидных клеток в зависимости от внешних обстоятельств.
• 5НТ3а-экспрессирующие вставочные нейроны — гетерогенная группа, к которой относят большую часть вставочных нейронов поверхностных корковых слоев. Получая информацию от корковых и таламических нейронов, они могут участвовать в процессах обучения, осуществляя влияние на корковые нейронные цепи. Он нейроглиеформных клеток (паутинообразных клеток)—одного из главных типов вставочных нейронов слоев II и III — веерообразно расходятся дендриты, обладающие уникальным свойством образования синапсов друг с другом и другими типами вставочных нейронов; этот факт указывает на их важнейшую роль в синхронизации корковых нейронных цепей. Другая морфологически гетерогенная группа вставочных нейронов, помимо ГАМК, экспрессирует вазоактивный интестинальный полипептид; другие типы вставочных нейронов этой группы экспрессируют также холецистокинин и другие пептидные рецепторы.
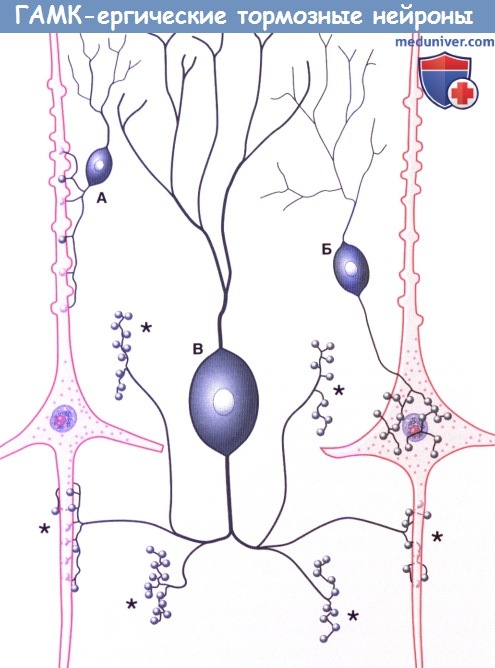
Три морфологических типа ГАМК-ергических тормозных нейронов:
А. Аксодендритическая клетка, образующая синапс с апикальным отделом пирамидного нейрона.
Б. Корзинчатая клетка, образующая аксоаксональные синапсы с пирамидными клетками.
В. Клетка-канделябр, формирующая аксоаксональные синапсы (*) с начальными сегментами аксонов двух пирамидных клеток, показанных здесь, и с начальными сегментами четырех других клеток, не показанных на рисунке.
(На основе DeFelipe, 1999, с разрешения автора.)
г) Афферентные волокна. Восходящие волокна к любому отделу коры могут иметь четыре различных источника (в первую очередь, корковые) и оканчиваются в разных отделах.
1. Длинные и короткие ассоциативные волокна от мелких и средних пирамидных клеток в слоях II и III из других корковых зон того же полушария.
2. Комиссуральные волокна от средних пирамидных клеток слоев II и III проходят через мозолистое тело от аналогичных или топографически идентичных участков противоположного полушария.
3. Таламо-корковые волокна от соответствующего специфического или ассоциативного ядра (например, волокна от вентрального заднего таламического ядра к соматосенсорной коре) и от дорсомедиального таламического ядра к предлобной (префронтальной) коре (описана ниже) оканчиваются в пластинке IV. Неспецифические таламо-корковые волокна от внутрипластинчатых ядер оканчиваются во всех пластинках.
д) Эфферентные волокна. Единственные эфферентные волокна, покидающие кору больших полушарий, представлены возбуждающими аксонами пирамидных клеток. Одна часть аксонов пирамидных клеток соединяется с длинными и короткими ассоциативными волокнами, другая образует комиссуральные или проекционные пути. Ассоциативные и комиссуральные пути составляют большой объем белого вещества полушарий головного мозга.
• Примерами коротких ассоциативных путей (проходят между соседними зонами коры в составе поверхностного белого вещества в виде U-волокон) служат пути, направляющиеся в моторную кору от сенсорной коры и обратно. Примерами длинных ассоциативных путей служат волокна между префронтальной корой (лежащей кпереди от моторных зон коры) и ассоциативными сенсорными зонами. Источник этих волокон — пирамидные клетки, расположенные преимущественно в слоях II и III.
• Комиссуральные волокна мозга представлены исключительно аксонами пирамидных клеток, проходящих через мозолистое тело, переднюю и заднюю спайки (и другие мелкие спайки) к аналогичным участкам противоположного полушария (например, волокна от первичной корковой зоны направляются к соответствующей ассоциативной области другого полушария), а также к несоответствующим зонам [подобные комиссуральные связи отсутствуют между первичной зрительной корой, первичной соматосенсорной и моторной корой, иннервирующими дистальный отдел верхней конечности]. Волокна начинаются от пирамидных клеток, расположенных преимущественно в слоях II и III.
• Основная часть проекционных волокон от первичной сенсорной и моторной коры подходит к базальным ганглиям. К таламусу направляются волокна от всех отделов коры. К другим основным проводящим путям относят корково-мостовой (к ипсилатеральным ядрам моста), корково-ядерный (к двигательным и чувствительным ядрам черепных нервов моста и продолговатого мозга на противоположной стороне) и корково-спинномозговой. Источником этих волокон служат пирамидные клетки, расположенные преимущественно в слоях V и VI (отдающие волокна преимущественно к специфическим релейным ядрам таламуса).
Редактор: Искандер Милевски. Дата публикации: 22.11.2018
15.1. Общие сведения о головном мозге

I - стадия трёх мозговых пузырей;
II -стадия образования пяти отделов мозга (II).
15.1.2. Cоставные части головного мозга
а) Согласно предыдущей схеме, в головном мозгу различают 5 следующих друг за другом отделов.
б) Перечислим их ещё раз, рассмотрев эти отделы (и составляющие их образования)
в окончательно сформированном мозгу.
2. а) Собственно задний мозг включает две части :
3. а) Средний мозг тоже имеет две части ; его составляют
Схема - сагиттальный срез головного мозга человека.
15.1.3. Рефлекторные дуги,
образованные с участием головного мозга
15.1.3.1. Введение
б) Поэтому кору больших полушарий называют также плащом или мантией .
б) Надо отметить, что нижеследующее,
15.1.3.2. Сознаваемая мышечно-суставная (проприоцептивная) и кожная
чувствительность: рефлекторный ответ
б) Если в реакцию вовлечены мышцы головы, то эффекторные нейроны располагаются
15.1.3.3. Бессознательная проприоцептивная
чувствительность и координация движений
а) Одни импульс ы к мозжечку (12-13) поступают от соответствующих рецепторов мышц и суставов -
б) Другие импульсы идут
Осуществив анализ поступающих сигналов, кора мозжечка (12) передаёт импульсы
а ) От указанных структур идут нисходящие пути
б ) Однако существует и более короткий путь - непосредственно
15.1.3.4. Рефлекторный ответ на слуховые и зрительные раздражения
а) Слуховое или зрительное возбуждение передаётся
а ) Помимо такой бессознательной реакции, возможен и сознательный ответ на слуховые и зрительные воздействия -
б) Кроме того, нейроны сетчатки идут
а) Подробней строение и проводящие пути головного мозга изучаются в курсе нормальной анатомии.
б) Мы же теперь остановимся на гистологической структуре некоторых частей заднего и переднего мозга.
15.2. Задний мозг
15.2.1. Продолговатый мозг
а) На поперечном срезе продолговатого мозга выявляется серое и белое вещество, т.е.
б) Многие из них уже упоминались нами при описании проводящих путей.
15.2.1.1. Компоненты рефлекторной дуги, связанной
с сознаваемой кожной и мышечно-суставной чувствительностью (п. 15.1.3.2)
| Рисунок - продолговатый мозг: I - вид сзади; II - поперечный срез на уровне нижней оливы, вид спереди. | |
 |  |
I. Компоненты восходящих путей
б) На задней поверхности продолговатого мозга им соответствуют одноимённые бугорки (1 -2).
б) Эти волокна и образуют медиальную петлю (3) , которая
II. Компоненты нисходящих путей
3. На вентральной поверхности продолговатого мозга этим путям соответствуют два продолговатых тяжа - пирамиды (5), переходящие далее в передние канатики спинного мозга.
Таким образом, в продолговатом мозгу имеются перекрёсты двух длинных проводящих путей - чувствительных и двигательных.
15.2.1.2. Компоненты рефлекторной дуги, связанной
с координацией движения (п. 15.1.3.3)
б) Напомним: этот путь
15.2.1.3. Другие образования продолговатого мозга
5. Видимо, ретикулярная формация отвечает
б) А. С этими ядрами связаны центры дыхания и кровообращения, задающие ритм дыхания и сердцебиения.
15.2.2.1. Общий вид
I. Составные части
2. а) Поверхность мозжечка образует узкие извилины ,
отделённые друг от друга бороздками .
II. Вид на разрезе
2. При этом в извилинах обнаруживаются:
3. Эти структуры придают мозжечк у характерный вид, обозначаемый как "древо жизни" .
(На самом деле, повреждение мозжечка не создаёт прямой опасности для жизни).
4. Помимо коры , серое вещество находится в составе нескольких
1,а. Препарат - мозжечок собаки. Импрегнация азотнокислым серебром.

15.2.2.2. Кора мозжечка. Клеточный состав
I. Три слоя коры
б) Следовательно, кору можно видеть как справа, так и слева от борозды.
2. В коре выделяют 3 слоя (перечисляем, начиная с поверхностного) :

II. Характеристика слоёв коры
б) Содержит клетки двух видов.
1. Звёздчатые клетки - в поверхностной части слоя; небольшие, имеют много отростков.
2 . Корзинчатые клетки - в нижней трети слоя, более крупные;
б) Среди них - клетки трёх видов.
1. Клетки-зёрна - наиболее многочисленные и мелкие; их аксоны
III. Грушевидные нейроны
15.2.2.3. Основные рефлекторные дуги в коре мозжечка
1. Между клетками коры мозжечка имеются строго определённые связи, и сами клетки выполняют определённые функции.
2. При этом можно выделить
3. Рассмотрим вначале первые из них , т.е. основные способы прохождения сигнала .
I. Кратчайшая рефлекторная дуга

II. Более длинная дуга
б) В том числе они возбуждают грушевидные клетки .
3. Дальнейшее - как в предыдущей дуге:
15.2.2.4. Корректировка сигнала в коре мозжечка
Теперь остановимся на дополнительных рефлекторных дугах и их функциональной роли.
I. Корректировка (ограничение) входного сигнала
1. Так, клетки-зёрна контактируют своими аксонами (в молекулярном слое) не только с грушевидными клетками, но и
II. Корректировка (ограничение) ответа
2. Те и другие своими аксонами
3. Торможение торможения , реализуемое в дуге, приводит
из пяти типов клеток коры мозжечка (фигурирующих в этих дугах)
15.2.2.5. Ещё одна схема
2. а) Более длинная рефлекторная дуга:
В нём происходит
15.3. Передний мозг
15.3.1.1. Строение
I. Общие сведения
II. Ядра гипоталамуса
Вот наиболее известные из ядер гипоталамуса .
паравентрикулярные ядра (2) - в боковой стенке III желудочка.
2. а) Обе пар ы ядер вырабатывают нейрогормоны пептидной природы :
Более детально взаимоотношения между гипоталамусом и гипофизом мы будем рассматривать при изучении эндокринной системы (тема 22) .
15.3.1.2. Просмотр препарата
1. Этот препарат нам уже встречался в п. 12.2.4.4.
а) (Малое увеличение) 
б) Последние окрашиваются в фиолетовый цвет и обуславливают вышеотмеченную неоднородность цитоплазмы.
4 . Поскольку нейроциты принадлежат супраоптическому ядру,
б) (Большое увеличение) 
15.3.2. Кора больших полушарий
15.3.2.1. Общие сведения
I. Поверхность больших полушарий мозга
б) Э то значительно увеличивает её площадь.
2. И вновь, как в мозжечке, на разрезе выявляется кора-

4 . а) Как те, так и другие располагаются в определённом порядке , отчего говорят о
II. Клетки коры: пирамидные нейроны
Основными клетками коры больших полушарий являются пирамидные нейроны различного размера.
2. Характерна ориентация пирамидных нейронов:
либо вниз (в белое вещество),
либо поворачивает к поверхности коры.
III. Прочие нейроны коры
2. Б ольшинство из последних тормозят пирамидные клетки,
15.3.2.2. Подразделение коры на слои
I. Перечень слоёв
2. а) Однако эти с лои выявляются не так чётко, как в мозжечке.
б) Поэтому вначале их продемонстрируем не на препарате, а на приведённой схеме.
3. Вот перечень слоёв коры (начиная с самого поверхностного):
II. Характеристика слоёв
2. Нейронов в нём очень мало;
б) Все клетки - небольшого размера, что и придаёт слою зернистый вид.
б) Имеется также небольшое количество тормозных нейронов.
2. а) Поскольку содержание непирамидных клеток невелико,
б) Как уже отмечалось, аксоны этих клеток образуют пирамидные пути, идущие к мотонейронам спинного мозга.
2. Кроме этих вертикальных (проекционных) путей, в данном слое, как и в предыдущем, имеется много горизонтальных нервных волокон.
2. а) Его клеточный состав:
б) Концентрация клеток по направлению к белому веществу убывает.
3. Через слой проходят
III. Резюмирующие замечания
2. В связи с этим, различают 2 типа коры больших полушарий:
15.3.2.3. Просмотр препарата
Теперь посмотрим, как выглядит многослойная структура коры на гистологическом препарате.
I. Среднее увеличение
I. Молекулярный слой:
II. Наружный зернистый слой:

IV. Внутренний зернистый слой :
V. Ганглионарный слой: крупные пирамидные клетки.
VI Слой полиморфных клеток:
II. Большое увеличение
б) Один из отростков, отходящих от основания нейронов, - аксон ( 3 ) .

15.3.2.4. Нейроглия в сером веществе головного мозга
I. Астроцитарная глия
б) Астроциты имеют небольшое тело и многочисленные отростки.
в) Причём, преобладают протоплазматические астроциты (1) - с толстыми и короткими отростками.
II. Микроглия
б) Но, кроме того, в коре больших полушарий содержатся и микроглиальные клетки (1) - мелкие, с ветвящимися отростками.
2. Этот препарат тоже уже встречался - в п. 12.3.5.
3. Клетки макро- и микроглии выполняют в коре разнообразные функции -

III. Олигодендроглия
б) Как правило, эти волокна являются миелиновыми.
15.3.2.5. Нервные связи в коре больших полушарий
I. Сопоставление клеток коры мозжечка и коры больших полушарий
(тогда как интенсивность принимаемого ими сигнала и формируемого ответа регулируется другими клетками).
2. Но имеются, по крайней мере, 3 отличия. –
а) Пирамидные клетки располагаются не в один ряд, а более диффузно.
б) Аксоны этих клеток направляются не в подкорковые ядра (как в мозжечке), а
2. Причём, подобно ситуации в мозжечке,
сигнал от афферентных волокон может передаваться на пирамидные клетки
Нейроны:
пирамидные - синие,
звёздчатые (возбуждающие) - жёлтые,
тормозные - чёрные.
Волокна:
афферентные - красные,
эфферентные - синие .

аналоги корзинчатых и звёздчатых нейронов мозжечка.
б) О ни образуют тормозные синапсы , соответственно, на телах и аксонах пирамидных клеток.
2. Последние тормозят практически все виды тормозных нейронов и тем самым
Мультиполярные нейроны коры ГМ весьма разнообразны по форме. Среди них можно выделить: пирамидные; звёздчатые; веретенообразные; паукообразные; горизонтальные.
Пирамидные нейроны составляют основную и наиболее специфическую для коры ГМ форму (80—90 % всех нейронов). Пирамидные клетки различных слоёв коры отличаются размерами и имеют разное функциональное значение. Мелкие клетки представляют собой вставочные нейроны, аксоны которых связывают отдельные участки коры одного полушария (ассоциативные нейроны) или двух полушарий (комиссуральные нейроны).
Эти клетки встречаются в разных количествах во всех слоях коры. Особенно богата ими кора ГМ человека. Аксоны крупных пирамидных нейронов принимают участие в образовании пирамидных путей, проецирующих импульсы в соответствующие центры мозгового ствола и СМ.
Нейроны коры расположены нерезко отграниченными слоями. Каждый слой характеризуется преобладанием какого-либо одного вида клеток. В двигательной зоне коры различают 6 основных слоёв: Молекулярный; Наружный зернистый; Пирамидальных нейронов; Внутренний зернистый; Ганглионарный (слой клеток Беца); Слой мультиформных (полиморфных) клеток.
Кора полушарий ГМ также содержит мощный нейроглиальный аппарат, выполняющий трофическую, защитную, опорную и разграничительную функции. На медиальной и нижней поверхности полушарий сохранились участки старой, древней коры, которые имеют двухслойное и трехслойное строение.
Молекулярный слой. Молекулярный слой коры содержит небольшое количество мелких ассоциативных клеток веретеновидной формы. Их аксоны проходят параллельно поверхности мозга в составе тангенциального сплетения нервных волокон молекулярного слоя. Основная масса волокон этого сплетения представлена ветвлениями дендритов нейронов нижележащих слоёв.
В новой коре больших полушарий нейроны сгруппированы в шесть-семь корковых слоёв.
В слое I (молекулярном) Сравнительно широкий слой. Мало клеточных тел, в нем находятся преимущественно терминали аксонов и их синапсы на дендритах. Выполняет несколько функций: пограничную, защитную, трофическую, газообменную. В слое II (наружном гранулярном (Наружный зернистый слой)) Сравнительно узкий, характеризуется высокой частотой расположения нервных клеток, преобладают мелкие пирамидные нейроны. Дендриты этих клеток уходят в молекулярный слой, а аксоны в КГМ этого же полушария. Клетки обеспечивают связь с другими участками коры этого же полушария. Слой III (наружный пирамидный) Наиболее широкий слой, содержит пирамидные нейроны—мелкие, средние (преимущественно), крупные, которые образуют 3 подслоя. Дендриты этих клеток достигают молекулярного слоя, аксоны части клеток заканчиваются в других участках коры этого же полушария или противоположного полушария. Они образуют ассоциативные нервные пути. Выполняют ассоциативные функции. Часть нервных клеток—аксоны крупных пирамидных нейронов уходят в БВ и участвуют в образовании нисходящих проекционных двигательных путей. Этот слой выполняет наиболее мощные ассоциативные функции. В слое IV (внутреннем гранулярном (Внутренний зернистый слой)) Узкий, больше всего звездчатых клеток, в том числе шипиковых. Обычно густоклеточный и варьирует по ширине и чёткости границ с 3 и 5 пограничными слоями, при этом в некоторых полях развит очень сильно (например, в зрительной зоне коры), а иногда отсутствует (например, в прецентральной извилине). Выполняет ассоциативные функции. В слое V (внутреннем пирамидном (ганглиозный)) Довольно широкий, содержит крупные и средние пирамидные нейроны. В нем располагаются гигантские нейроны (клетки Беца). Дендриты уходят в вышележащие слои, достигают молекулярного слоя. Аксоны уходят в БВ и образуют нисходящие двигательные пути. Слой VI (полиформный (Мультиформный)) Уже, чем ганглиозный. Содержит клетки разнообразные по форме, но преобладают веретеновидные нейроны. Их дендриты также уходят в вышележащие слои, достигают молекулярного слоя, а аксоны поступают в БВ и участвуют в образовании нисходящих нервных двигательных путей.
1-4 слои являются ассоциативными. 5-6 слои являются проекционными.
кора головного мозга или кора головного мозга - это нервная ткань, которая покрывает поверхность полушарий головного мозга. Упомянутая другая форма, это составляет самую превосходящую область мозга.
Эта структура мозга достигает максимального развития у приматов, менее развита у других животных и связана с более сложным развитием познавательной и интеллектуальной деятельности..
Кора головного мозга является основной областью мозга для функционирования человека. В этом регионе выполняются такие функции, как восприятие, воображение, мысль, суждение или решение..
Анатомически он состоит из серии тонких слоев, состоящих из серого вещества, которые расположены над широким скоплением путей белого вещества..
Кора головного мозга принимает извилистую форму, поэтому, если она будет расширяться, она будет иметь очень обширную массу. В частности, исследования показывают, что общая площадь коры головного мозга может составлять около 2500 квадратных сантиметров..
Аналогично, эта большая масса мозга характеризуется наличием огромного количества нейронов внутри. В общем, постулируется, что в коре головного мозга имеется около 10 миллиардов нейронов, что составляет около 50 триллионов синапсов.
Основные характеристики коры головного мозга объясняются ниже. Его слои, его нейроны и его функциональная организация определены, и функции, которые выполняются в этой области мозга, рассматриваются.
- 1 Характеристика коры головного мозга
- 2 слоя
- 2.1 Молекулярный слой
- 2.2 Внешний зернистый слой
- 2.3 Внешний пирамидальный слой
- 2.4 Внутренний зернистый слой
- 2.5 Слой лимфатического узла
- 2.6 Мультиформный слой
- 3 Функциональная организация
- 3.1 Чувствительные зоны
- 3.2 Моторные зоны
- 3.3 Области ассоциации
- 4 нервных клетки
- 4.1 Пирамидальные клетки
- 4.2 Звездные клетки
- 4.3 Шпиндельные клетки
- 4.4 Кахал горизонтальные клетки
- 4.5 клеток Мартинотти
- 5 ссылок
Характеристика коры головного мозга
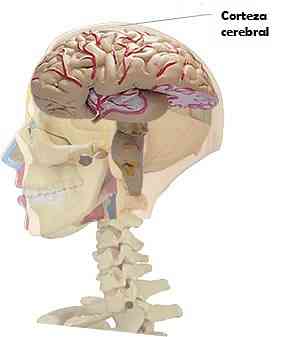
Кора головного мозга животных млекопитающих представлена слоем серого вещества, которое покрывает два полушария головного мозга.
Он состоит из очень сложной структуры, в которой различные сенсорные органы представлены в определенных областях или областях, которые называются первичными сенсорными областями..
Каждое из пяти чувств, которыми обладают люди (зрение, осязание, обоняние, вкус и осязание), развивается в определенной области коры. То есть каждая сенсорная модальность имеет ограниченную территорию внутри коры головного мозга.
Помимо сенсорных областей кора головного мозга также имеет множество вторичных соматических, ассоциативных и моторных областей. В этих областях развиваются корковые и ассоциативно-афферентные системы, что дает начало обучению, памяти и поведению..
В этом смысле кора головного мозга считается особенно важной областью при развитии превосходной деятельности человеческого мозга..
Наиболее продвинутые и сложные процессы человека, такие как мышление, планирование, организация или ассоциация, осуществляются в разных областях коры головного мозга..
По этой причине кора головного мозга представляет собой структуру, которая приобретает максимальную сложность с человеческой точки зрения. Кора головного мозга является результатом медленного эволюционного процесса, который мог начаться более 150 миллионов лет назад.
слои
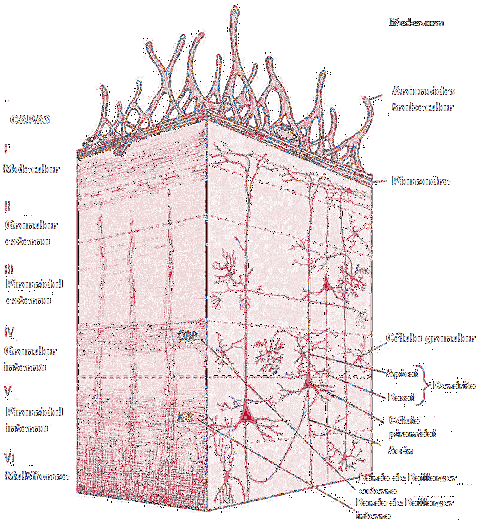
Основная характеристика коры головного мозга состоит в том, что она состоит из различных слоев серого вещества. Эти слои составляют структуру коры и определяют ее структурную и функциональную организацию..
Кроме того, слои коры головного мозга характеризуются не только определением со структурной точки зрения, но и с филогенетической точки зрения..
То есть каждому из слоев коры головного мозга соответствует свой эволюционный момент. В начале человеческого вида мозг был менее развит, а кора была меньше слоев.
В ходе эволюции видов эти слои увеличивались, что связано с увеличением познавательных и интеллектуальных способностей человека с течением времени..
Молекулярный слой, также известный как плексиформный слой, является наиболее поверхностным участком коры головного мозга и, следовательно, новейшим началом.
Он состоит из плотной сети нервных волокон, которые тангенциально ориентированы. Эти волокна происходят от дендритов пирамидных и веретенообразных клеток, аксонов звездчатых и мартиноттиевых клеток..
Афферентные волокна, которые возникают в таламусе, ассоциации и комиссурах, также могут быть обнаружены в молекулярном слое. Будучи самой поверхностной областью коры, в молекулярном слое между нейронами установлено большое количество синапсов..
Внешний зернистый слой является вторым наиболее поверхностным участком коры и лежит ниже молекулярного слоя. Содержит большое количество мелких пирамидных и звездчатых клеток.
Дендриты клеток внешнего зернистого слоя оказываются в молекулярном слое, а аксоны входят в более глубокие слои коры головного мозга. По этой причине внешний зернистый слой связан с различными областями коры..
Внешний пирамидальный слой, как следует из его названия, состоит из пирамидальных клеток. Он характеризуется неправильной формой, то есть размер слоя увеличивается от поверхностного предела до самого глубокого предела.
Дендриты нейронов пирамидального слоя переходят в молекулярный слой, а аксоны перемещаются в виде проекционных, ассоциативных или комиссуральных волокон к белому веществу, расположенному между слоями коры головного мозга..
Внутренний зернистый слой состоит из звездчатых ячеек, которые расположены в очень компактной форме. У этого есть высокая концентрация горизонтально расположенных волокон, известных как внешняя полоса Baillarger.
Ганглиозный слой или внутренний пирамидальный слой содержит очень большие и средние по размеру пирамидальные клетки. Кроме того, они содержат большое количество волокон, расположенных горизонтально, которые образуют внутреннюю полосу Baillarger.
Наконец, мультиформный слой, также известный как слой полиморфных клеток, в основном содержит веретенообразные клетки. Кроме того, он содержит модифицированные пирамидальные клетки, которые содержат треугольные или яйцевидные клетки тела.
Многие из нервных волокон мультиформного слоя проникают в нижележащее белое вещество и связывают слой с промежуточными областями..
Функциональная организация
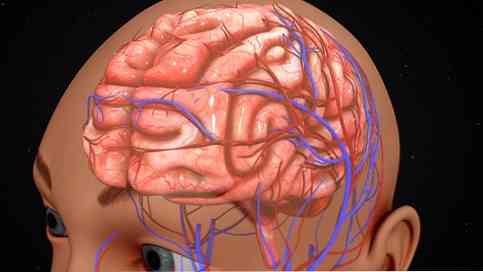
Кора головного мозга также может быть организована в соответствии с мероприятиями, проводимыми в каждом регионе. В этом смысле определенные участки коры головного мозга обрабатывают специфические сигналы чувствительного, моторного и ассоциативного характера..
Сенсорные области - это области коры головного мозга, которые получают информацию чувствительного характера и тесно связаны с восприятием..
Информация поступает в кору головного мозга в основном через заднюю половину обоих полушарий головного мозга. Первичные области содержат наиболее прямые связи с периферическими сенсорными рецепторами.
С другой стороны, вторичные сенсорные и ассоциативные области обычно соседствуют с первичными областями. Как правило, они получают информацию как из первичных областей ассоциации, так и из нижних областей мозга..
Основная задача областей ассоциации и второстепенных областей состоит в том, чтобы интегрировать чувствительный опыт для создания моделей распознавания и поведения. Основными чувствительными областями коры головного мозга являются:
- Первичная соматосенсорная зона (зоны 1, 2 и 3).
- Основная зрительная зона (зона 17).
- Основная слуховая зона (зоны 41 и 42).
- Основная зона вкуса (область 43).
- Первичная обонятельная зона (зона 28).
Моторные зоны находятся в передней части полушарий. Они несут ответственность за инициирование мозговых процессов, связанных с движением и начало такой деятельности.
Наиболее важные моторные зоны:
- Основная моторная зона (зона 4).
- Область бурения языка (области 44 и 45).
Области ассоциации коры головного мозга коррелируют с более сложными функциями интеграции. Эти регионы выполняют такие действия, как процессы памяти и познания, управление эмоциями и развитие рассуждений, воли или суждения..
Кроме того, области ассоциации играют особенно важную роль в развитии личности и черт характера людей. Кроме того, это область мозга, необходимая для определения интеллекта.
Области ассоциации включают определенные двигательные области, а также определенные сенсорные области.
Нервные клетки

Кора головного мозга имеет большое разнообразие клеток внутри. В частности, пять различных типов нейронов были определены в этой области мозга.
Пирамидальные клетки - это нейроны, которые характеризуются наличием пирамидальной формы. Большинство из этих клеток имеют диаметр от 10 до 50 микрометров.
Однако есть и крупные пирамидальные клетки. Они известны как ячейки Бетца и могут иметь диаметр до 120 микрометров..
Как мелкие пирамидальные клетки, так и крупные пирамидальные клетки обнаруживаются при моторно-прецентральной цирконвуляции и в основном выполняют действия, связанные с движением..
Звездчатые клетки, также известные как гранулезные клетки, представляют собой маленькие нейроны. Они обычно имеют диаметр около 8 микрометров и имеют многоугольную форму.
Веретенообразные клетки - это нейроны, у которых вертикальная продольная ось на поверхности. Они сосредоточены в основном в более глубоких корковых слоях мозга.
Аксон этих нейронов происходит в нижней части тела клетки и направлен к белому веществу в виде проекции, ассоциации или комиссурального волокна..
Горизонтальные клетки Кахала представляют собой маленькие веретенообразные клетки, которые ориентированы горизонтально. Они находятся в самых поверхностных слоях коры головного мозга и играют решающую роль в развитии этой области мозга.
Этот тип нейронов был открыт и описан Рамоном и Кахалем в конце 19-го века, и последующие исследования показали, как важные клетки должны координировать нейронную активность.
Чтобы достичь своего положения в коре головного мозга, горизонтальные клетки кахала должны скоординированно мигрировать во время эмбриогенеза головного мозга. То есть эти нейроны путешествуют от места рождения к поверхности коры головного мозга.
Что касается молекулярной структуры этих нейронов, Виктор Боррелл и Оскар Марин из Института нейробиологии Аликанте показали, что горизонтальные клетки Кахала представляют собой ориентацию нейронных слоев коры во время эмбрионального развития..
На самом деле, дисперсия этих клеток происходит на начальных стадиях эмбрионального развития. Клетки рождаются в разных областях мозга и мигрируют на поверхность мозга, чтобы полностью покрыть его.
Наконец, недавно было продемонстрировано, что менингеальные мембраны выполняют иные функции, чем защитные, которые предполагались в начале. Менинги служат субстратом или путем горизонтальных клеток кахала для их тангенциальной миграции вдоль поверхности коры..
Последние нейроны, которые составляют нейронную активность коры головного мозга, являются хорошо известными клетками Martinotti. Они состоят из небольших многоформных нейронов, присутствующих на всех уровнях коры головного мозга..
Эти нейроны обязаны своим именем Карло Мартинотти, студенту-исследователю Камило Гольджи, который обнаружил существование этих клеток коры головного мозга..
Клетки Martinotti характеризуются тем, что они являются многополярными нейронами с короткими дендритными дендритами. Они распространяются через несколько слоев коры головного мозга и отправляют свои аксоны в молекулярный слой, где образуются аксональные древовидные образования..
Недавние исследования этих нейронов показали, что клетки Martinotti участвуют в тормозном механизме мозга..
В частности, когда пирамидальный нейрон (который является наиболее распространенным типом нейрона в коре головного мозга) начинает перевозбуждать, клетки Martinotti начинают передавать ингибирующие сигналы на окружающие нервные клетки..
В этом смысле делается вывод, что эпилепсия может быть тесно связана с дефицитом клеток Martinotti или дефицитом активности этих нейронов. В эти моменты нервная передача мозга этими клетками больше не регулируется, что вызывает дисбаланс в функционировании коры..
Читайте также:
