
Морфогенез нервной системы это
Не нужно доказывать, что особенности поведения теснейшим образом связаны со структурной и функциональной организацией нервной системы. Поэтому раннее развитие, в том числе и эмбриональное, когда происходит дифференциация нервных клеток и образование связей между ними и прочими системами организма, является одним из важнейших этапов, на котором формируются многие индивидуальные особенности поведения человека. Что же известно об этом современной науке?
Центральная нервная система состоит из огромного числа клеток, отличающихся высочайшей степенью дифференциации по сравнению со всеми другими тканями организма. Эти клетки образуют сложные функционально значимые связи между собой и с остальными органами и тканями. Клетки образуют скопления, популяции, обладающие специфической морфологией и функциями. То есть по мере развития складывается сложнейшая функциональная система организма. И вместе с тем все клетки, слагающие эту систему, обладают одним и тем же набором генов.
Мы уже упоминали, что количество ДНК в клеточном ядре мыши составляет 6х10-6 микрограмм. Это число для дрозофилы в 25 раз меньше. Однако если мы захотим сравнить в этом отношении мышь и человека, то получим поразительный результат. Оказывается, количество ДНК приблизительно одинаково у мыши, шимпанзе и человека. Это кажется парадоксом, если учесть, насколько возрастает сложность организации мозга при переходе от мыши к человеку. Число мозговых клеток увеличивается с 5-6 миллионов до нескольких десятков миллиардов. Вместе с тем количество ДНК в ядре оплодотворенной клетки существенно не меняется. Еще больше поражает то, что максимальное количество генов на такое количество ДНК (пусть это будет даже 200 000 или миллион) абсолютно несопоставимо с разнообразием нейронов, не говоря уже о синапсах. Таким образом, эволюционное усложнение мозга не сопровождается, по всей видимости, параллельным усложнением организации генома. Какой же механизм позволяет сформировать сложнейший орган человека - его мозг - на основе столь малого числа генетических детерминант, как происходит дифференциация нейронов, как складываются их популяции, как они образуют функционально значимые связи между собой и с иннервируемыми органами?
На все эти вопросы пытается ответить интенсивно развивающаяся область науки - нейробиология развития, однако она еще очень далека от того, чтобы дать полное объяснение процессам морфогенеза нервной системы.
Анатомические изменения в нервной системе координированы с подобными изменениями в остальном теле и с появлением функции. Эта координация осуществляется путем реципрокного взаимодействия между растущей нервной системой и остальными органами и тканями. В результате создается соответствие между структурой и функцией. В основе развития нервной системы лежат динамические процессы, идущие при участии внутренних и внешних по отношению к нейрону или нейронной популяции событий.
События, связанные с развитием отдельной клетки, включают в себя рождение клетки, миграцию, дифференциацию, образование связей и возможную гибель на ранних этапах онтогенеза.
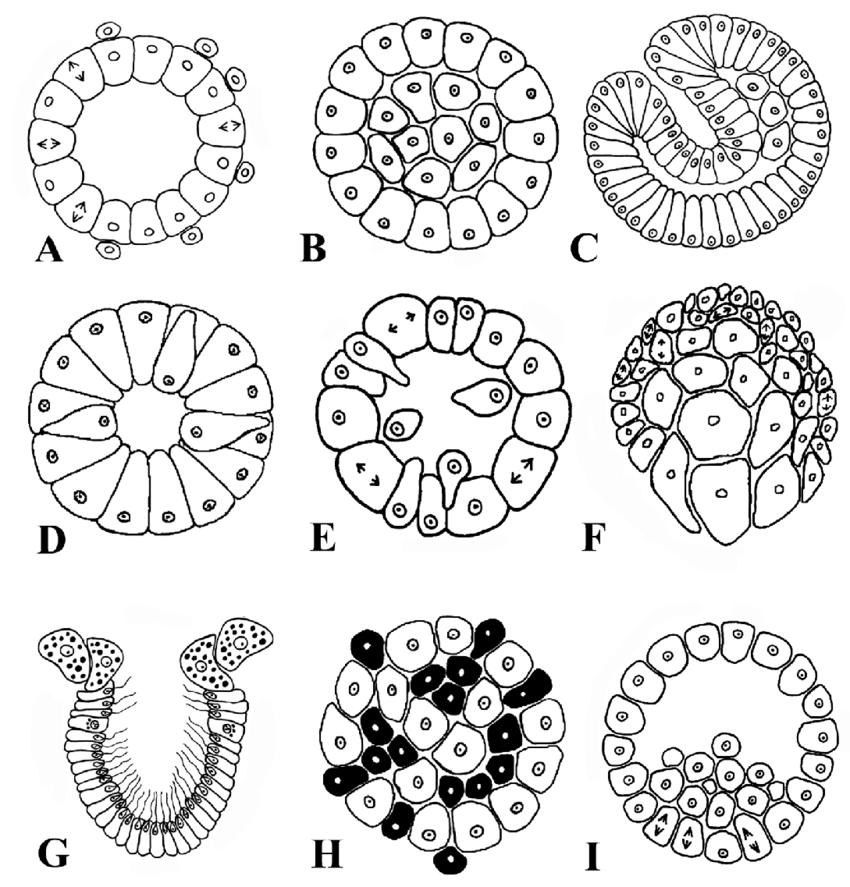
Рождение. Увеличение числа нейронов происходит в результате митоза, который приводит к появлению новых клеток из уже существующих. У позвоночных большинство нейронов рождается на ранних стадиях развития в результате деления клеток неврального эпителия, который тянется вдоль нервной трубки. На 25-й день эмбриона человека нервная трубка еще открыта с обоих концов, но позже она замыкается, а в передней ее части образуются три пузырька, которые будут формировать головной мозг. Между 4 и 6 неделями жизни нервная система эмбриона человека напоминает нервную систему рыбы. Митотическое деление нервных клеток в этот период происходит настолько интенсивно, что через несколько месяцев их количество уже составляет несколько десятков миллиардов. Временами скорость деления достигает 250 000 клеток в минуту. Через 16-20 недель после оплодотворения деление нервных клеток, по-видимому, прекращается. Таким образом, количество нервных клеток достигает своего максимума задолго до рождения.
Образование новых нейронов возможно и в постэмбриональный период, но это не является правилом. Конечное же число нейронов определяется не только процессами их рождения, но и процессами естественной гибели, которые являются важным элементом развития нервной ткани. Естественная гибель нейронов играет особенно существенную роль в морфогенезе нервной ткани у птиц и млекопитающих. По мере роста, дифференцировки и установления функциональных связей сверхпроизведенные нейроны, которые как бы не находят себе места в складывающейся функциональной системе, погибают. Гибель нейронов регулируется локальными условиями и является относительно поздним эмбриональным событием, обычно происходящим после формирования синапсов. В промежутке между своим рождением и решающим моментом, "жить ему или погибнуть", нейрон мигрирует к месту своей окончательной локализации, проходит дифференциацию и устанавливает свои первичные афферентные и эфферентные связи (рис. 9.5 а, б).
Миграция. Большинство нейронов имеют место своей локализации на некотором расстоянии от места своего рождения. Так что миграция нейронов является обязательным и важным этапом нейронального развития. До сих пор является загадкой, как нейроны находят место своего окончательного назначения. Очевидно, главным источником информации по поводу миграции являются поверхности, окружающие клетку. Эти поверхности отличаются по своей адгезивности (то есть вязкости, способности склеивать клетки между собой), и именно это свойство, по-видимому, является наиболее важным для установления путей миграции. Градиент адгезивности заставляет нейрон передвигаться. Большая часть адгезивного субстрата для нейронов связана с другим важным типом клеток, представленных в нервной ткани, а именно с клетками глии. Например, при формировании коры большую роль играет радиальная глия, которая простирает отростки сквозь все развивающиеся слои коры вплоть до ее поверхности. По этим "дорогам" нейроны, образующиеся в глубинных слоях вблизи мозгового желудочка, скользят в более поверхностные слои коры (рис. 9.6 а, б). Однако существуют и другие "маршруты" для передвижения нейронов.
Известно, что клетки коры мозга образуют шесть слоев. V взрослого слой I является самым поверхностным, а слой VI - самым глубоким. Зона деления нервных клеток находится еще глубже. Было бы естественно предположить, что по мере деления слои клеток накапливаются подобно осадочным породам, и более молодые клетки оказываются ближе к зоне деления. На самом деле случается обратное. Слой VI - самый глубокий - соответствует наиболее рано сформировавшимся клеткам, тогда как все остальные слои занимают свои места, проходя через все уже установившиеся слои, так что слой I соответствует самым молодым клеткам (рис. 9.7). Принципы умножения, миграции и размещения клеток в слоях, по-видимому, являются общими для всей коры, что позволяет предположить участие в этом процессе небольшого числа генов.
Дифференциация. Другой загадкой нейроэмбриогенеза являются процессы дифференциации нейронов. Как клетки с идентичным генным составом преобразуются в столь разнообразные формы, которые характерны для нейронов зрелого мозга? Во-первых, клетки нервной ткани дифференцируются на два крупных класса. Клетка становится либо нейрональной, либо глиальной. Внутри каждого из этих классов также идет дифференцировка, особенно характерная для нейронов. Нейроны отличаются по форме, по размерам, по характеру своих отростков и по нейротрансмиттерам, которые они используют (адренэргические, холинэргические). Причины возникновения этих различий до конца не поняты. Ясно только, что они включают как процессы взаимодействия клетки с ее окружением, так и дифференциальную активацию и подавление генов на определенных стадиях развития. Проходя последовательные стадии развития, нейрон приобретает свойства, которые могут влиять на прохождение последующих стадий. Скорее всего, решающие воздействия на окончательное формирование нейрона оказывают процессы взаимодействия с другими клетками. Например, нейротрансмиттерная специфичность нейронов в автономной нервной системе зависит от клеточного окружения. Вещества, продуцируемые теми тканями, которые иннервирует данный нейрон, способны изменять его специфичность. Например, вещества из клеток сердца побуждают нейрон становиться холинэргическим.
Выживание и гибель. Для выживания нейронов, по-видимому, решающим является воздействие трофических факторов. К трофическим факторам относят вещества, продуцируемые другими клетками, которые способны поддерживать рост, функционирование и выживание данной клетки. Число нейронов, образующихся в результате митоза на ранних эмбриональных стадиях, примерно в два или три раза превышает то количество нейронов, которое выживает на более поздних стадиях. Естественная нейрональная гибель происходит вскоре за установлением афферентных и эфферентных связей с другими клетками. Предполагается, что естественная гибель нейронов происходит в процессе конкуренции за ограниченный ресурс трофических факторов, поставляемых теми тканями, с которыми нейроны устанавливают функциональные связи. Однако точная природа этой конкуренции пока неясна. Нейроны, которые успешно взаимодействуют с клетками-мишенями, получают необходимые трофические факторы, а остальные, которые не могут успешно взаимодействовать, отмирают. Идет своего рода естественный отбор. Наиболее известным из трофических факторов является фактор роста нервов (ФРН), который был описан Р. Леви-Монтальчини (Levi-Montalcini R.) в 1975 г. Оказалось, что слюнная железа мыши является превосходным источником ФРН, что позволило вести интенсивные исследования с его применением. ФРН продуцируется естественным путем внутри нервной системы и ее периферических мишеней. Некоторым популяциям нейронов ФРН требуется для выживания и роста. Инъекция ФРН в развивающийся эмбрион увеличивает выживаемость нейронов в период естественной гибели клеток.
Образование связей. Аксоны и дендриты корковых нейронов начинают появляться намного раньше, чем устанавливаются все шесть слоев коры. В это же время формируются и первые синапсы. Так, предшественники пирамидных клеток посылают свои аксоны к соответствующим ядрам таламуса, а аксоны таламических нейронов растут по направлению к коре. Связь между корой и таламусом начинает формироваться реципрокно и синхронно еще до того, как образуются II и III слои. Главные пути связей в мозговой коре человека закладываются еще до рождения.
Многие процессы, лежащие в основе миграции нейронов, имеют место и при образовании отростков нервных клеток - аксонов и дендритов. Аксоны и дендриты движутся по направлению к клеткам-мишеням, постепенно отрастая из тела клетки. Скорость роста может составлять 15-20 микрон в час. На ведущем конце нейрита (аксона или дендрита) имеется специальная структура, называемая конусом роста (рис. 9.8).
Человек - сложнейшая система, отдельные части которой находятся в постоянном взаимодействии между собой и окружающей средой. Каждый орган, ткань, клетка выполняют свою строго определенную функцию, что делает наш организм максимально устойчивым к внешним раздражителям. Как это стало возможным, как наш организм развивался со временем, чтобы стать таким совершенным, изучает отдельный раздел генетики под названием "морфогенетика".
Что такое морфогенез?
Морфогенез - это последовательность определенных этапов, которые проходит в своем развитии отдельный орган или система органов. Она включает в себя два основных подвида: онтогенез (индивидуальное развитие органов и систем в пределах одного организма) и филогенез (эволюционное, или историческое, развитие систем и отдельных органов).
В основном в изучении морфогенеза человека принимает участие генетика. Но есть и другие науки, которые также вносят вклад в накопление знаний о развитии органов человека:
- молекулярная биология;
- физиология;
- биохимия;
- сравнительная анатомия;
- палеонтология;
- теория эволюции.
Особенности и история
Онтогенез характеризует собой развитие тканей, органов и систем в эмбриональный период развития. Помимо зрелого человеческого организма, процесс морфогенеза может также происходить в культурах клеток и новообразованиях. Также данный процесс происходит у неклеточных форм, хотя у них и отсутствует эмбриональная стадия.
Впервые предположения о формировании органов и о том, как условия внешней и внутренней среды могут на него повлиять, были высказаны в 1952 году Аланом Тьюрингом в его работе под названием "Химические основы морфогенеза". В этой работе был описан математический базис самоорганизации клеток и тканей.
Однако знания о морфогенезе значительно увеличились при открытии ДНК и ее структуры, развитии таких наук, как молекулярная биология и биохимия. Немаловажным стало открытие молекулярных механизмов работы генов.
Основные процессы
В эмбриогенезе, то есть внутриутробном развитии, выделяют следующие основные процессы морфогенеза:
- Клеточное размножение - увеличение их количества, при котором набор хромосом должен быть строго распределен среди последующих клеток, иначе могут произойти аномалии развития. Это возможно путем двух процессов: дробления и митоза.
- Детерминация - означает, что развитие каждой клетки предопределено. Какие-то из них могут быть только половыми, другие - лишь эпителиальными, третьи - нервными и т.д.
- Дифференциация - клетка становится более специфической и выполняет строго предназначенную ей функцию.
- Индукция - одни клетки способны влиять на развитие других. Таким образом, наличие индукции способно активировать процесс дифференциации благодаря наличию специальных факторов регуляции морфогенеза.
- Интеграция - объединение структур, занимающих более низкую нишу эмбриогенеза, в более сложные. Например, клетки образуют ткани. Ткани - органы. Органы - системы.
- Адгезия - способность клеток образовывать контакты для обмена информацией
- Миграция клеток - способность клеток перемещаться в пределах эмбриона.
- Апоптоз - запрограммированная гибель клетки.
Основные уровни
Морфогенез - это сложная, высокоточная система, которая происходит на многих уровнях организма. Условно выделяют два основных уровня регуляции развития всего организма:
- молекулярный;
- клеточный.

Молекулярный уровень
Структуры, которые имеют возможность влиять на морфогенез, получили название морфогенов. Их подразделяют на следующие классы:
- Факторы транскрипции - могут взаимодействовать с ДНК, имеют способность регулировать экспрессию генов через катализацию, то есть ускорение, их транскрипции.
- Структуры, способные регулировать межклеточные контакты. К примеру, на одном из этапов внутриутробного развития, гаструляции, клетки теряют связь друг с другом, мигрируют в новое место, где вновь обретают способность образовывать новые контакты.
Клеточный уровень
Что он собой представляет? Морфогенез - это последовательность реакций, которые невозможны без постоянного взаимодействия клеток друг с другом. Ниша стволовой клетки - это именно то связующее звено между одной клеткой и всем организмом. Существуют такие типы клеток, как эпителиальные и мезенхимальные. В процессе эмбриогенеза они могут переходить друг в друга. Вследствие такого процесса как клеточная дифференциация. При этом возможно также их перемещение из одного места в эмбрионе в другое.
Этапы развития организма
Человеческий организм в своем развитии проходит определенные этапы. И хотя морфогенез - это механизм, который наибольшее свое значение обретает на этапе внутриутробного развития, и после рождения наши органы меняются вплоть до самой смерти. Множество патологий развития органов происходят именно в эмбриональном периоде. Однако стоит выделить и другие стадии морфогенеза:
- Пренатальный - тот самый внутриутробный, от зачатия и до родов. Он делится, в свою очередь, на эмбриональный и фетальный периоды. Их сроки: 8 недель после зачатия и после окончания эмбрионального периода до родов соответственно.
- Перинатальный - период с 22 недель внутриутробного развития до 7 первых дней жизни.
- Интернатальный - период всей родовой деятельности.
- Неонатальный - первые 28 дней жизни.
- Постнатальный - 15 лет после рождения.
Прогенез, фетогенез, киматогенез: что это такое?
Однако можно выделить еще один период, который начался еще до зачатия - это прогенез. Это этап, который подразумевает под собой закладку половых клеток в организме будущих родителей. Именно прогенез можно назвать этапом начала морфогенеза.
Отдельно также выделяют такой период, как фетогенез. В нем есть еще два периода:
- ранний фетальный - с 76 по 180 день беременности;
- поздний фетальный - с 181 по 280 день беременности.
Киматогенез, в свою очередь, включает все перечисленные выше процессы: эмбриогенез, прогенез и фетогенез. А изучает его отдельная наука, которая получила название "киматология". Изучением развития эмбриона и плода занимается эмбриология.
Филогенез
Выше подробно была рассмотрена такая часть морфогенеза как онтогенез. Однако стоит уделить внимание также филогенезу - развитию отдельных видов в процессе эволюции. Как и в онтогенезе, в филогенезе также есть свои принципы, на которых он построен:
- Дифференциация - в процессе формирования организма определенная часть клеток становится обособленной, обретая собственные функции и уникальное строение. Эта совокупность клеток образует отдельную ткань или орган.
- Интеграция - укрепление связей между отдельными клетками и целыми системами органов.
- Каждый орган имеет как основную функцию, так и множество второстепенных, которые дают возможность преобразовываться и адаптироваться к новым условиям.
- Основная функция может как утрачиваться, так и обретать большее значение в зависимости от новых условий.
- Развитие органов может быть как в сторону большего совершенствования и усложнения, то есть прогрессирования, так и в сторону упрощения (регрессирования).
- Корреляция - все части организма находятся в тесной взаимосвязи друг с другом, изменение одной его части несет за собой изменение всего организма.
Таким образом, морфогенез и его биологическая сущность крайне комплексны и сложны. Постоянное взаимодействие отдельных этапов индивидуального и эволюционного развития организма обеспечивают возможность к его существованию в постоянно меняющейся внешней среде.
Источником развития нервной ткани и нейроглии является нервная пластинка 18-суточного зародыша человека. После нейруляции из нее образуются нервная трубка и ганглиозные пластинки. Эмбриональные зачатки состоят из малодифференцированных клеток — медуллобластов (матричных клеток). На ранних этапах гистогенеза происходит дифференцировка клеток, в результате чего возникают два направления их развития: нейробластическое и глиобластическое. В нейрогистогенезе различают стадии медуллобластов, нейробласта, молодого нейрона и зрелого нейрона. Стадия молодого нейрона характеризуется ростом отростков, увеличением объема клетки, образованием хроматофильной субстанции и появлением первых синапсов. Самой продолжительной стадией является стадия зрелого нейрона, на протяжении которой нейрон приобретает свою окончательную форму и специфическую гистохимическую организацию
Первыми созревают системы жизнеобеспечения, например сердечно-сосудистая система начинает функционировать на 4-ой неделе, несколько позже -гормональная система, но на этой стадии ее связи с центральными отделами (гипофизом) не выявляются. На втором месяце начинают созревать сенсорные системы: вестибулярная, кожная, причем фрагментарно, сначала в области губ, лица, потом пальцев рук, потом вкусовая. К третьему месяцу созревают системы управления мышцами лица, конечностей, причем сначала сгибателями верхних, а потом нижних, и позже – туловища. Формируются врожденные двигательные акты, как например – хватательный рефлекс. Второй триместр характеризуется формированием специфически человеческой в отличие от приматов пространственной организацией. В зрительном анализаторе формируются системы движения глаз, мигание и т.п, и к концу триместра возникают сложные системные взаимодействия: появляется общая и специфическая двигательная активность в связи изменением освещенности живота, резкими звуками или другими изменениями во внешней среде. К 20-ой неделе происходит становление основных ритмов биоэлектрической активности мозга.
3. Последовательность и сроки формирования структур ЦНС (а- эмбриона; б- плода).
Головной мозг развивается из ростральной части нервной трубки. Бо́льшая часть головного мозга (95 %) является производной крыловидной пластинки.
Эмбриогенез мозга проходит через несколько стадий.
Стадия трёх мозговых пузырей — у человека в начале четвёртой недели внутриутробного развития ростральный конец нервной трубки формирует три пузыря: Prosencephalon (передний мозг), Mesencephalon (средний мозг), Rhombencephalon (ромбовидный мозг, или первичный задний мозг).
Стадия пяти мозговых пузырей — у человека в начале девятой недели внутриутробного развития Prosencephalon окончательно делится на Telencephalon (конечный мозг) и Diencephalon (промежуточный мозг), Mesencephalon сохраняется, а Rhombencephalon делится на Metencephalon (задний мозг) и Myelencephalon (продолговатый мозг).
Последовательная смена основных этапов формирования мозга во II триместре беременности обусловлена с одной стороны генетической программой плода, с другой — факторами внешней среды через организм матери.
· ранняя стадия пренатального онтогенеза (5—6 нед) митоз нейронов в зоне мозговых пузырей и миграция нейронов к месту своей дальнейшей локализации.
· В 8 нед выявляется закладка новой коры. Закладка ее представляет собой однородное образование без какого-либо деления на отдельные формации. Клетки этого периода однородные, округлые, интенсивно окрашенные.
· С 16 нед развития плода происходит миграция клеток к месту новой коры. В этом периоде выделяются все основные области коры (прецентральная, постцентральная, височная, зрительная, лимбическая и др.).
· К 22 нед. гестации начинают прогрессировать процессы внутри коры, появляются цитоархитектонические слои, впервые определяются отдельные цитоархитектонические поля мозга.
· В период 22—28 нед внутриутробного развития мозга плода происходит яркая гетерохромность образования отдельных цитоархитектонических полей, свойственных человеку
· К 27—28-й неделе в коре больших полушарий мозга плода человека выделены все основные цитоархитектонические поля. Начинается активный процесс клеточной дифференцировки.
· 28 нед интенсивно увеличивается площадь поверхности коры.
4. Возрастные морфологические и функциональные особенности головного мозга человека.
У новорожденного головной мозг относительно большой, масса его в среднем 390 что составляет 12 —13 % массы тела (у взрослых — примерно 2,5%). К концу первого года жизни масса мозга удваивается, а к 3—4 годам утраивается. В дальнейшем (после 7 лет) масса головного мозга возрастает медленно и к 20 —29 годам достигает максимального значения (1 355 г — у мужчин и 1 220 г — у женщин). В последующие возрастные периоды, вплоть до 60 лет у мужчин и 55 лет у женщин, масса мозга существенно не изменяется, а после 55 — 60 лет отмечается некоторое уменьшение ее. Лобная доля большого мозга у новорожденного сильно выпуклая и относительно невелика. Височная доля высокая. Островковая доля (островок) расположена глубоко. До 4 лет жизни головной мозг ребенка растет равномерно в высоту, длину и ширину, в дальнейшем преобладает рост мозга в высоту. Наиболее быстро растут лобная и теменная доли. На поверхности полушарий большого мозга у новорожденного уже имеются борозды и извилины. Основные борозды (центральная, латеральная и др.) выражены хорошо, а ветви основных борозд и мелкие извилины обозначены слабо. В дальнейшем, по мере увеличения возраста ребенка, борозды становятся глубже, извилины между ними рельефнее. Миелинизация нервных волокон в онтогенетически более старых отделах (ствол мозга) начинается и заканчивается раньше, чем в более новых отделах. Взаимоотношения борозд и извилин с костями и швами крыши черепа у новорожденных несколько иные, чем у взрослого. Центральная борозда расположена на уровне теменной кости. Нижнелатеральная часть этой борозды находится на 1,0 — 1,5 см краниальнее чешуйчатого шва. Теменно-затылочная борозда лежит на 12 мм кпереди от ламбдовидного шва. Соотношения борозд, извилин мозга и швов, характерные для взрослого человека, устанавливаются у детей 6 — 8 лет. Мозолистое тело у новорожденного тонкое, короткое. Оно растет одновременно с развитием и увеличением полушарий большого мозга, преимущественно в краниальном и каудальном направлениях, располагаясь над полостью промежуточного мозга (над III желудочком). С развитием полушарий большого мозга увеличивается толщина ствола мозолистого тела (до 1 см у взрослого человека) и валика мозолистого тела (до 2 см), что обусловлено увеличением количества комиссуральных нервных волокон.
Морфо-функциональная организации спинного мозга и мозгового ствола.
1. Общее анатомическое описание спинного мозга.
Спинной мозг состоит из ряда элементов, имеющих сходное строение (взаиморасположение ядер серого вещества и волокон белого)- сегментов, из которых на периферию выходит по паре симметричных спиномозговых нервов, иннервирующих определенную область тела – метамер. Количество сегментов в спинном мозге:
· шейный отдел 8 позвонков,
· грудной отдел – 12 позвонков,
· поясничный отдел – 5 позвонков,
· крестцовый отдел – 5 позвонков,
· копчиковый отдел - 1- 2 позвонка (копчиковый отдел может быть редуцирован). Ядра сегментов, расположенные друг над другом, образуют столбы (передние, задние, а в грудном отделе дополнительные боковые). Столбы в центральной части, и по периферии - белое вещество, организованное в канатики (передние, задние и боковые).
Объем спинного мозга у человека составляет около 2% относительно массы головного мозга, диаметр около 1 см., а длина порядка 42 – 45 см ( у женщин короче), располагается в верхней части позвоночного канала. Соотношение моторных элементов, т.е. клеток которые обеспечивают иннервацию скелетной мускулатуры, и чувствительных клеток, которые обеспечивают рецепцию тела, составляет 1 к 5. Соответственно и проводящих путей примерно такое же соотношение: порядка 2 млн. - это восходящие чувствительные пути по которым в мозг поступает информация о состоянии мышц и кожи (болевая, тактильная. температурная, вибрационная и т.д.), а нисходящих чуть более 400 тыс. Остальные (около10 млн.) нейроны ассоциативные, и они обеспечивают внутрисегментарные интегративные процессы.
Морфологически спинной мозг разделен на симметричные правую и левую половину продольными щелями. В вентральной части это глубокая и широкая щель, а в дорзальной щель менее выражена. Внутри с.м. по центру, имеется цилиндрический канал, который на ранних стадиях развития (первые годы) заполнен ликвором, а затем замещается желеобразной массой. В местах выхода передних и задних корешков, вдоль спинного мозга, имеются соответствующие углубления – бороздки которые являются анатомическими границами между заднебоковыми и переднебоковыми отделами.
2. Морфофункциональная характеристика сегменто-ядерного апппарата спинного мозга.
В передних рогах имеется около 5-ти ядер. Клетки группируются в ядра в соответствии с функциональным значением и, в общем, можно сказать, что внутренние ядра иннервируют осевую мускулатуру, обеспечивающую регуляцию позы, а ядра, расположенные на периферии, иннервируют мышцы производящие более сложные, тонкие движения. Задние отделы серого вещества - задние столбы (рога) состоят из скоплений чувствительных и ассоциативных клеток, разнообразных по морфологии и более мелких, по сравнению с клетками передних рогов. Разнообразие морфологии, в какой-то степени, определяется их функциональной специализацией, поскольку они обеспечивают первичную обработку отдельных видов кожной и мышечно-суставной чувствительности.
3. Структурная организация проводящей системы (белого вещества) спинного мозга.
Белое вещество спинного мозга представлено отростками нервных клеток, которые составляет тракты, или проводящие пути спинного мозга:
1) короткие пучки ассоциативных волокон , связывающие сегменты спинного мозга, расположенные на различных уровнях;
2) восходящие (афферентные, чувствительные) пучки, направляющиеся к центрам большого мозга и мозжечка ; (дорсальная)
3) нисходящие (эфферентные, двигательные) пучки, идущие от головного мозга к клеткам передних рогов спинного мозга . (вентральная)
Белое вещество спинного мозга располагается по периферии серого вещества спинного мозга и представляет собой совокупность миелинизированных и отчасти маломиелинизированных нервных волокон , собранных в пучки. В белом веществе спинного мозга расположены нисходящие волокна (идущие из головного мозга) и восходящие волокна , которые начинаются от нейронов спинного мозга и проходят в головной мозг . По нисходящим волокнам передается преимущественно информация от моторных центров головного мозга к мотонейронам (двигательным клеткам) спинного мозга. По восходящим волокнам поступает информация как от соматических, так и от висцеральных чувствительных нейронов. Расположение восходящих и нисходящих волокон носит закономерный характер. На спинной (дорсальной) стороне расположены преимущественно восходящие волокна , а на брюшной (вентральной) - нисходящие волокна .
Межсегментные волокна образуют собственные пучки, расположенные тонким слоем по периферии серого вещества и осуществляющие связи между сегментами спинного мозга. Они присутствуют в переднем, заднем и боковом канатиках.
4. Морфофункциональная характеристика ядерных образований продолговатого мозга.
Структурно по разнообразию и строению ядер продолговатый мозг сложнее, чем спинной. В отличие от спинного мозга он не имеет метамерного, повторяемого строения, серое вещество в нем расположено не в центре, а ядрами к периферии. В продолговатом мозге находятся оливы, связанные со спинным мозгом, экстрапирамидной системой и мозжечком — это тонкое и клиновидное ядра проприоцептивной чувствительности (ядра Голля и Бурдаха). Продолговатый мозг за счет своих ядерных образований и ретикулярной формации участвует в реализации вегетативных, соматических, вкусовых, слуховых, вестибулярных рефлексов. Особенностью продолговатого мозга является то, что его ядра, возбуждаясь последовательно, обеспечивают выполнение сложных рефлексов, требующих последовательного включения разных мышечных групп, что наблюдается, например, при глотании. Продолговатый мозг регулирует ряд сенсорных функций: рецепцию кожной чувствительности лица — в сенсорном ядре тройничного нерва; первичный анализ рецепции вкуса — в ядре языкоглоточного нерва; рецепцию слуховых раздражений — в ядре улиткового нерва; рецепцию вестибулярных раздражений — в верхнем вестибулярном ядре. В задневерхних отделах продолговатого мозга проходят пути кожной, глубокой, висцеральной чувствительности, часть из которых переключается здесь на второй нейрон (тонкое и клиновидное ядра). На уровне продолготоватого мозга перечисленные сенсорные функции реализуют первичный анализ силы и качества раздражения, далее обработанная информация передается в подкорковые структуры для определения биологической значимости данного раздражения.
Кроме того, продолговатый мозг организует рефлексы поддержания позы. Эти рефлексы формируются за счет афферентации от рецепторов преддверия улитки и полукружных каналов в верхнее вестибулярное ядро; отсюда переработанная информация оценки необходимости изменения позы посылается к латеральному и медиальному вестибулярным ядрам. Эти ядра участвуют в определении того, какие мышечные системы, сегменты спинного мозга должны принять участие в изменении позы, поэтому от нейронов медиального и латерального ядра по вестибулоспинальному пути сигнал поступает к передним рогам соответствующих сегментов спинного мозга, иннервирующих мышцы, участие которых в изменении позы в данный момент необходимо. Большая часть автономных рефлексов продолговатого мозга реализуется через расположенные в нем ядра блуждающего нерва, которые получают информацию о состоянии деятельности сердца, сосудов, пищеварительного тракта, легких, пищеварительных желез и др. В ответ на эту информацию ядра организуют двигательную и секреторную реакции названных органов.
5. Сравнительная характеристика морфологии белого вещества продолговатого мозга и варолиева моста.
ПРОДОЛГОВАТНЫЙ МОЗГ
Белое вещество продолговатого мозга содержит длинные и короткие волокна. К длинным относятся проходящие транзитно в передние канатики спинного мозга нисходящие пирамидные пути, частью перекрещивающиеся в области пирамид. Кроме того, в ядрах задних канатиков находятся тела вторых нейронов восходящих чувствительных путей. Их отростки идут от продолговатого мозга к таламусу,. Волокна этого пучка образуют медиальную петлю, , которая в продолговатом мозге совершает перекрест, , и в виде пучка волокон, расположенных дорсальнее пирамид, между оливами — идет далее. Таким образом, в продолговатом мозге имеется два перекрестка длинных проводящих путей: вентральный двигательный, , и дорсальный чувствительный,
К коротким путям относятся пучки нервных волокон, соединяющие между собой отдельные ядра серого вещества, а также ядра продолговатого мозга с соседними отделами головного мозга.
МОСТ
. Белое вещество: тракты, проходящие транзитом в восходящем направлении (все виды кожной чувствительности) и нисходящем направлении (моторные тракты - пирамидной и экстропирамидной систем). Также здесь начинается новый восходящий чувствительный тракт слуховой и вестибулярной систем. Этот тракт мощным пучком белого вещества трапецевидной формы образует в средней части латеральную петлю. Эта часть моста носит название – покрышка. Кроме того, здесь располагается скопление нейронов образующих собственные моторные ядра моста, обеспечивающие переключение и копирование команд, идущих из моторной коры головного мозга, для их отправления в мозжечок. Аксоны этих нейронов образуют компактные жгуты белого вещества - средние мозжечковые ножки. Таким образом, в мозжечок поступают команды для коррекции произвольных движений. Волокна белого вещества, восходящих чувствительных трактов, имеют дугообразное направление и переходят на противоположную сторону, образуя спиномозговую и медиальную петли. Взаиморасположение волокон восходящих и нисходящих направлений аналогично взаиморасположению в спинном и продолговатом мозге, т.е., вентрально – эфферентные нисходящие тракты, дорзально – афферентные, чувствительные тракты, которые здесь перекрещиваются, переходя на противоположную сторону, образуя медиальную, латеральную, спиномозговую петли.
Читайте также:
