
Нейрональные и мембранные рецепторы автономной нервной системы
Вегетативная нервная система поддерживает гомеостаз. ВНС управляет такими висцеральными функциями, как кровообращение, пищеварение и выделение, главным образом без условного или сознательного контроля. ВНС также модулирует функцию эндокринных желез, регулирующих метаболизм. ВНС имеет сенсорные и моторные компоненты и разделяется на симпатическую и парасимпатическую системы. Первые нейроны симпатической системы расположены в промежуточных рогах тораколюмбального отдела спинного мозга; синапс со вторым набором нейронов находится в параили превертебральном симпатическом ганглии. В парасимпатической системе первые нейроны расположены либо в черепно-мозговом нерве, в автономных ядрах, либо в промежуточном роге сакрального отдела спинного мозга; синапс со вторым набором нейронов находится или в автономном ганглии (в случае черепно-мозговых нервов), или в эффекторной ткани непосредственно. ВНС имеет три главных компонента:
• афферентный (центростремительный, чувствительный);
• центральный объединяющий;
• эфферентный.
Афферентный компонент несет информацию от нейрональных физиологических рецепторов, расположенных в концах центростремительных нервов, к спинному мозгу и более высоким областям ЦНС. Большая часть этой информации обрабатывается в пределах гипоталамуса и других нижележащих областей мозга. После обработки соответствующий сигнал посылается от ЦНС вниз по эфферентным нервам к исполнительным органам (см. рис. 8.1, 8.9), названным так потому, что они отвечают на деятельность в ЦНС.
На основе различий анатомии и медиаторов эфферентную часть ВНС подразделяют на три системы:
• парасимпатическую (холинергическую);
• симпатическую (адренергическую);
• неадренергическую нехолинергическую (НАНХ).
Ацетилхолин — нейромедиатор холинергической системы. Ацетилхолин — нейромедиатор, высвобождаемый из пресинаптического окончания в автономном ганглии и в окончаниях нервов в исполнительном органе. Рецепторами для ацетилхолина служат холинорецепторы, которые подразделяют на мускариновые и никотиновые.
Норадреналин — нейромедиатор адренергической системы. Другая важная составляющая ВНС — адренергическая система. До сих пор неизвестно, какой нейромедиатор использовался в этой системе первоначально — эпинефрин или норэпинефрин. Сейчас известно, что за исключением надпочечников, которые секретируют эпинефрин (адреналин), нейроме-диатором в адренергической системе является норэпинефрин.
Ацетилхолин — ганглионарный медиатор для холинергической и адренергической систем. Эфферентные нервы и для холинергической, и для адренергической систем происходят из соответствующих частей ствола мозга и спинного мозга. Эфферентные нервы образуют синапс в ганглии, расположенном вне органа, где основным нейромедиатором является АХ:
• в адренергической системе ганглии находятся в цепочке вблизи спинного мозга, известной как паравертебральная симпатическая цепочка;
• в холинергической системе ганглий обычно располагается внутри или вблизи эффекторного органа.
Несмотря на явное анатомическое различие, оба типа ганглиев используют АХ как основной ганглионарный нейромедиатор, активирующий никотиновые рецепторы.
Нейромедиаторы могут модулировать собственное высвобождение. Нейромедиаторы могут модулировать собственное высвобождение. Нейромедиаторы могут активировать пресинаптические рецепторы на нейроне, что ингибирует высвобождение самих нейромедиаторов.
Учебное видео анатомии вегетативной нервной системы (ВНС)
- Вернуться в оглавление раздела "фармакология"

Нервная система
Раздражимость или чувствительность – характерная черта всех живых организмов, означающая их способность реагировать на сигналы или раздражители.
Сигнал воспринимается рецептором и передается с помощью нервов и (или) гормонов к эффектору, который осуществляет специфическую реакцию или ответ.
Животные имеют две взаимосвязанные системы координации функций – нервную и гуморальную (см. таблицу).
Нервная регуляция
Гуморальная регуляция
Электрическое и химическое проведение (нервные импульсы и нейромедиаторы в синапсах)
Химическое проведение (гормоны) по КС
Быстрое проведение и ответ
Более медленное проведение и отстроченный ответ (исключение - адреналин)
В основном кратковременные изменения
В основном долговременные изменения
Специфический путь распространения сигнала
Неспецифический путь сигнала (с кровью по всему телу)к специфической мишени
Ответ часто узко локализован (например, один мускул)
Ответ может быть крайне генерализованным (например, рост)
Нервная система состоит из высокоспециализированных клеток со следующими функциями:
- восприятие сигналов – рецепторы;
- преобразование сигналов в электрические импульсы (трансдукция);
- проведение импульсов к другим специализированным клеткам – эффекторам, которые получив сигнал, дают ответ;
Связь между рецепторами и эффекторами осуществляют нейроны .
Нейрон – это структурно – функциональная единица НС.

Нейрон — электрически возбудимая клетка, которая обрабатывает, хранит и передает информацию с помощью электрических и химических сигналов. Нейрон имеет сложное строение и узкую специализацию. Нервная клетка содержит ядро, тело клетки и отростки (аксоны и дендриты).
В головном мозге человека насчитывается около 90—95 миллиардов нейронов. Нейроны могут соединяться друг с другом, образуя биологические нейронные сети.
Нейроны разделяют на рецепторные, эффекторные и вставочные.
Тело нейрона: ядро (с большим количеством ядерных пор) и органеллы (ЭПС, рибосомы, аппарат Гольджи, микротрубочки), а также из отростков (дендриты и аксоны).
Нейроглия – совокупность вспомогательных клеток НС; составляет 40% общего объема ЦНС.
- Аксон – длинный отросток нейрона; проводит импульс от тела клетки; покрыт миелиновой оболочкой (образует белое вещество мозга)
- Дендриты - короткие и сильно разветвлённые отростки нейрона; проводит импульс к телу клетки; не имеют оболочки

Важно! Нейрон может иметь несколько дендритов и обычно только один аксон.
Важно! Один нейрон может иметь связи со многими (до 20 тысяч) другими нейронами.
- чувствительные – передают возбуждение от органов чувств в спинной и головной мозг
- двигательные – передают возбуждение от головного и спинного мозга к мышцам и внутренним органам
- вставочные – осуществляют связь между чувствительными и двигательным нейронами, в спинном и головном мозге
Нервные отростки образуют нервные волокна.
Пучки нервных волокон образуют нервы.
Нервы – чувствительные (образованы дендритами), двигательные (образованы аксонами), смешанные (большинство нервов).
Синапс – это специализированный функциональный контакт между двумя возбудимыми клетками, служащий для передачи возбуждения

У нейронов синапс находится между аксоном одной клетки и дендритом другой; при этом физического контакта не происходит – они разделены пространством - синаптической щель.
Нервная система:
- периферическая (нервы и нервные узлы) – соматическая и автономная
- центральная (головной и спинной мозг)
В зависимости от характера иннервации НС:
- Соматическая – управляет деятельностью скелетной мускулатуры, подчиняется воле человека
- Вегетативная (автономная) – управляет деятельностью внутренних органов, желез, гладкой мускулатуры, не подчиняется воле человека
Соматическая нервная система – часть нервной системы человека, представляющая собой совокупность чувствительных и двигательных нервных волокон, иннервирующих мышцы (у позвоночных — скелетные), кожу, суставы.
Она представляет часть периферической нервной системы, которая занимается доставкой моторной (двигательной) и сенсорной (чувственной) информации до центральной нервной системы и обратно. Эта система состоит из нервов, прикрепленных к коже, органам чувств и всем мышцам скелета.
- спинномозговые нервы – 31 пара; связаны со спинным мозгом; содержат как двигательные, так и сенсорные нейроны, поэтому смешанные;
- черепномозговые нервы – 12 пар; отходят от головного мозга, иннервируют рецепторы головы (за исключением блуждающего нерва – иннервирует сердце, дыхание, пищеварительный тракт); бывают сенсорными, моторными (двигательными) и смешанными
Рефлекс – это быстрый автоматический ответ на раздражитель, осуществляемый без осознанного контроля головного мозга.
Рефлекторная дуга – путь, проходимый нервными импульсами от рецептора до рабочего органа.
- в ЦНС – по чувствительному пути;
- от ЦНС – к рабочему органу – по двигательному пути
- рецептор (окончание дендрита чувствительного нейрона) – воспринимает раздражение
- чувствительное (центростремительное) нервное волокно – передает возбуждение от рецептора к ЦНС
- нервный центр – группа вставочных нейронов, расположены на разных уровнях ЦНС; передает нервные импульсы с чувствительных нейронов на двигательные
- двигательное (центробежное) нервное волокно – передает возбуждение от ЦНС к исполнительному органу

Простая рефлекторная дуга: два нейрона – чувствительный и двигательный (пример – коленный рефлекс)
Сложная рефлекторная дуга: три нейрона – чувствительный, вставочный, двигательный (благодаря вставочным нейронам происходит обратная связь между рабочим органом и ЦНС, что позволяет вносить изменения в работу исполнительных органов)
Вегетативная (автономная) нервная система – управляет деятельностью внутренних органов, желез, гладкой мускулатуры, не подчиняется воле человека.
Делится на симпатическую и парасимпатическую.

Обе состоят из вегетативных ядер (скопления нейронов, лежащих в спинном и головном мозге), вегетативных узлов (скопления нейронов, нейронов, за пределами НС), нервных окончаний (в стенках рабочих органов)
Путь от центра до иннервируемого органа состоит из двух нейронов (в соматической - один).
Место выхода из ЦНС
От спинного мозга – в шейный, поясничный, грудной отделы
От ствола головного мозга и ствола крестцового отдела спинного мозга
Местоположение нервного узла (ганглия)
По обе стороны спинного мозга, за исключением нервных сплетений (непосредственно в этих сплетениях)
В иннервируемых органах или вблизи них
Медиаторы рефлекторной дуги
В предузловом волокне –
в послеузловом - норадреналин
В обоих волокнах - ацетилхолин
Названия основных узлов или нервов
Солнечное, легочное, сердечное сплетения, брыжеечный узел
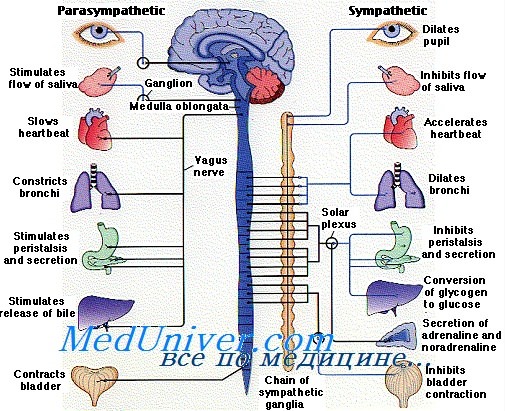
Общие эффекты симпатической и парасимпатической НС на органы:
- Симпатическая НС – расширяет зрачки, угнетает слюноотделение, повышает частоту сокращений, расширяет сосуды сердца, расширяет бронхи, усиливает вентиляцию легких, угнетает перистальтику кишечника, угнетает секрецию пищеварительных соков усиливает потоотделение, удаляет с мочой лишний сахар; общий эффект – возбуждающий, повышает интенсивность обмена, снижает порог чувствительности; активизирует во время опасности, стресса, контролирует реакции на стресс
- Парасимпатическая НС – сужает зрачки, стимулирует слезотечение, уменьшает частоту сердечных сокращений, поддерживает тонус артериол кишечника, скелетных мышц, снижает кровяное давление, уменьшает вентиляцию легких, усиливает перистальтику кишечника, расширяет артериолы в коже лица, увеличивает выделение с мочой хлоридов; общий эффект – тормозящий, снижает или не влияет на интенсивность обмена, восстанавливает порог чувствительности; доминирует в состоянии покоя, контролирует функции в повседневных условиях
Центральная нервная система (ЦНС) – обеспечивает взаимосвязь всех частей НС и их координированную работу
У позвоночных ЦНС развивается из эктодермы (наружного зародышевого листка)
ЦНС – 3 оболочки:
- твердая мозговая (dura mater) - снаружи;
- мягкая мозговая оболочка (pia mater) – прилегает непосредственно к мозгу.
Головной мозг расположен в мозговом отделе черепа; содержит
- белое вещество - проводящие пути между головным мозгом и спинным, между отделами головного мозга
- серое вещество - в виде ядер внутри белого вещества; кора покрывающая большие полушария и мозжечок
Масса головного мозга – 1400-1600 грамм.

5 отделов:
- продолговатый мозг– продолжение спинного мозга; центры пищеварения, дыхания, сердечной деятельности, рвота, кашель, чихание, глотание, слюноотделение, проводящая функция
- задний мозг – состоит из варолиевого моста и мозжечка; варолиев мост связывает мозжечок и продолговатый мозг с большими полушариями; мозжечок регулирует двигательные акты (равновесие, координация движений, поддержание позы)
- промежуточный мозг– регуляция сложных двигательных рефлексов; координация работы внутренних органов; осуществление гуморальной регуляции;
- средний мозг – поддержание тонуса мыщц, ориентировочные, сторожевые, оборонительные рефлексы на зрительные и звуковые раздражители;
- передний мозг (большие полушария) – осуществление психической деятельности (память, речь, мышление).
Промежуточный мозг включает таламус, гипоталамус, эпиталамус
Таламус – подкорковый центр всех видов чувствительности (кроме обонятельного), регулирует внешнее проявление эмоций (мимика, жесты, изменение пульса, дыхания)
Гипоталамус – центры вегетативной НС, обеспечивают постоянство внутренней среды, регулируют обмен веществ, температуру тела, чувство жажды, голода, насыщения, сна, бодрствования; гипоталамус контролирует работу гипофиза
Эпиталамус – участие в работе обонятельного анализатора
Передний мозг имеет два больших полушария: левое и правое
- Серое вещество (кора) находится сверху полушарий, белое – внутри
- Белое вещество – это проводящие пути полушарий; среди него – ядра серого вещества (подкорковые структуры)
Кора больших полушарий – слой серого вещества, 2-4 мм в толщину; имеет многочисленные складки, извилины
Каждое полушарие разделено бороздами на доли:
- лобная – вкусовая, обонятельная, двигательная, кожно- мускульная зоны;
- теменная – двигательная, кожно- мускульная зоны;
- височная – слуховая зона;
- затылочная – зрительная зона.
Важно! Каждое полушарие отвечает за противоположную сторону тела.
- Левое полушарие – аналитическое; отвечает за абстрактное мышление, письменную и устную речь;
- Правое полушарие – синтетическое; отвечает за образное мышление.
Спинной мозг расположен в костном позвоночном канале; имеет вид белого шнура, длина 1м; на передней и задней сторонах есть глубокие продольные борозды
В самом центре спинного мозга – центральный канал, заполненный спинномозговой жидкостью.
Канал окружен серым веществом (имеет вид бабочки), который окружен белым веществом.
- В белом веществе – восходящие (аксоны нейронов спинного мозга) и нисходящие пути (аксоны нейронов головного мозга)
- Серое вещество напоминает контур бабочки, имеет три вида рогов.
- передние рога – в них расположены двигательные нейроны (мотонейроны) – их аксоны иннервируют скелетные мышцы
- задние рога – содержат вставочные нейроны – связывают чувствительные и двигательные нейроны
- боковые рога – содержат вегетативные нейроны – их аксоны идут на периферию к вегетативным узлам
Спинной мозг – 31 сегмент; от каждого сегмента отходит 1 пара смешанных спинномозговых нервов, имеющих по паре корешков:
- передний (аксоны двигательных нейронов);
- задний (аксоны чувствительных нейронов.
Функции спинного мозга:
- рефлекторная – осуществление простых рефлексов (сосудодвигательных, дыхательных, дефекации, мочеиспускания, половых);
- проводниковая – проводит нервные импульсы от и к головному мозгу.

Повреждение спинного мозга приводит к нарушению проводниковых функций, вследствие чего – паралич.
Цитоархитектоника головного мозга человека организована таким образом, что более чем 10 млрд. нервных клеток, занимая относительно небольшое пространство и будучи сформированными в специализированные структуры, обеспечивают специфические функции мозга, связанные с восприятием, переработкой и проведением информации, в соответствии с которой осуществляется взаимодействие организма с внешней средой на основе высокой нейрональной специфичности и пластичности.
Основной структурной единицей нервной системы является нейрон. Различные типы нейронов дифференцируются по величине и форме тела клетки, а также по длине и степени ветвистости ее отростков.
Клеточное тело по своим размерам варьирует очень широко — от 5 до 100 мкм в диаметре. Оно содержит следующие органеллы: ядро, митохондрии, эндоплазматический ретикулум (гладкий и шероховатый), расположенные на цистернах эндоплазматического ретикулума и в свободном пространстве рибосомы и полисомы, комплекс Гольджи и различные внутриклеточные включения (гранулы гликогена, липидные капли, скопления частиц пигмента в особых нейронах и др.), везикулы, а также лизосомы. Группы параллельно расположенных цистерн шероховатого эндоплазматического ретикулума в виде ограниченных мембраной удлиненных цистерн с прикрепленными к ним рибосомами образуют субстанцию (тельца) Ниссля (тигроидное вещество). В цитоплазме имеются также нейрофиламенты и нейротрубочки (рис. 3).
Все перечисленные ультраструктурные органеллы клетки несут определенные функции. Ядро является субстратом основных генетических процессов в клетке. Митохондрии обеспечивают энергетический обмен — в них происходит окислительное фосфорилирование, приводящее к продукции энергии в виде молекул АТФ. Эндоплазматический ретикулум с прикрепленными на его цистернах рибосомами, а также свободно расположенные рибосомы и их комплексы (полисомы) имеют отношение к белковому обмену и синтетическим процессам в клетке. Лизосомам приписывается обменно-выделительная роль. Нейротрубочки и нейрофиламенты обеспечивают транспорт внутриклеточных веществ, имеющих отношение к проведению нервного импульса. Долгое время считали, что комплекс Гольджи, состоящий из параллельно расположенных цистерн и скоплений пузырьков на их концах, выполняет неопределенные обменно-выделительные функции. Хотя об этом комплексе известно далеко не все, привлекают к себе накопленные многими исследователями данные, свидетельствующие о том, что он играет главную роль в процессах обновления клеточной мембраны и ее генетически обусловленной специализации. Известно, что в комплексе Гольджи может происходить первичная сборка специализированных участков мембраны (рецепторов), которые в виде пузырьков транспортируются к наружной клеточной оболочке и встраиваются в нее. Такие исследования были обобщены А.А.Милохиным (1983).
От тела нейрона отходят основной отросток — аксон и многочисленные ветвящиеся отростки — дендриты. Длина аксонов различных нейронов колеблется от 1 мм до почти 1 м (нервное волокно). Вблизи окончания аксон разделяется на терминали, на которых расположены синапсы, контактирующие с телом и дендритами других нейронов. Синапсы вместе с нейрофиламентами и нейротрубочками являются субстратом проведения нервного импульса.

Рис. 3. Основные ультраструктурные компоненты нейрона.
Л — лизосомы; ШЭР — шероховатый эндоплазматический ретикулум (цистерны с прикрепленными рибосомами); М — митохондрии; НФ — нейрофиламенты; НТ — нейротрубочки; P — рибосомы; П — полисомы (комплексы рибосом); КГ — комплекс Гольджи; Я — ядро; ЦЭР — цистерны эндоплазматического ретикулума; ЛГ — липидные гранулы; ЛФ — липофусцин.
Кроме нейронов, в ткани мозга имеются различные виды глиальных клеток — астроглия, олигодендроглия, микроглия. Астроглия играет большую роль в обеспечении функции нейрона и формировании реакции мозговой ткани на вредоносные воздействия (инфекция, интоксикация и др.) — принимает участие в воспалительных процессах и ликвидации их последствий (заместительный глиоз). Олигодендроглия, как известно, обеспечивает миелинизацию нервного волокна и регулирует водный обмен (дренажная глия). Функции микроглии не до конца изучены, но ее значение подчеркивается размножением этих клеток при некоторых специфических процессах (участие в формировании сенильных бляшек; существует предположение о выработке микроглиальными клетками амилоидных фибрилл и т.п.).
Особые клеточные структуры характерны для желудочковых поверхностей головного мозга и его сосудистого сплетения. Желудочковая поверхность мозга покрыта клетками эпендимы с многочисленными микроворсинками и ресничками, принимающими участие в ликворообращении; сосудистое сплетение представлено "гроздьями" ворсинок, состоящих из капилляров, покрытых эпителиальными клетками. Их основная функция связана с обменом веществ между кровью и цереброспинальной жидкостью.

Типичный синапс состоит из пресинаптической терминали, постсинаптической области и расположенной между ними синаптической щели. Пресинаптическая терминаль является окончанием аксона. Она содержит нейрофиламенты, нейротрубочки, митохондрии и синаптические пузырьки, скопления которых видны около пресинаптической мембраны. Через последнюю переносятся содержащиеся в пузырьках нейротрансмиттеры. Постсинапс характеризуется наличием постсинаптического утолщения. Постсинаптическое утолщение представлено мембраной клетки с расположенными на ней рецепторами, входящими в структуру самой мембраны. Синапс представлен на рис. 4, а его электронно-микроскопическая картина на рис. 5.

Синапс может быть расположен на теле (соме) клетки — аксосоматический синапс, на дендрите — аксодендритный, на шипике дендрита — аксошипиковый (рис. 6) и на аксоне другой клетки — аксо-аксональный. Аксошипиковые синапсы несколько отличаются по своему строению от типичного синапса, что определяется строением шипика, имеющего в составе постсинапса особый шипиковый аппарат.
Взаимодействие пресинапса и постсинапса обеспечивается благодаря переносу нейротрансмиттера через синаптическую щель. Выделяясь из пресинапса, нейротрансмиттер (медиатор) может связываться с рецептором постсинаптической мембраны, инактивироваться в синаптической щели и частично вновь захватываться пресинаптической мембраной (процесс обратного захвата — reuptake ). Если рецептор постсинаптической мембраны заблокирован, то возможны оба последних процесса, а также избыточное накопление медиатора и связанное с этим развитие гиперчувствительности рецепторов (см. рис. 4).
Более подробно эти процессы рассматриваются в разделе "Нейрохимические системы мозга".
Рецепторы нейронов — это белковые структуры, расположенные на внешней поверхности мембраны клеток. Они способны "распознавать" и связывать биологически активные вещества — нейротрансмиттеры, различные эндогенные вещества, а также экзогенные соединения, в том числе психофармакологические средства. Соединения, которые могут связывать рецепторы, называются лигандами. Лиганды бывают эндогенными и экзогенными.
Распознавание лиганда рецептором обеспечивается специальными структурными элементами, или сайтами. Специфичность связывания лиганда происходит благодаря структурному соответствию молекул лиганда и рецептора, когда они подходят друг к другу по типу "ключ к замку". Реакция связывания является моментом запуска каскада внутриклеточных реакций, приводящих к изменению функционального состояния нейрона. В зависимости от "силы" и "прочности" связывания лиганда с рецептором употребляют понятие аффинности (сродства) лиганда по отношению к рецептору.
При связывании рецептора с лигандом может происходить как активация, так и блокада рецептора. В связи с этим говорят об агонистах и антагонистах рецепторов, а также о частичных агонистах (рис. 7).
Максимальную эффективность в отношении активации рецептора имеет полный агонист, минимальную (практически нулевую) — антагонист. Между ними находятся вещества, называемые частичными агонистами. Последние действуют значительно мягче, чем полные агонисты. Частичные агонисты, кроме того, занимая определенное пространственное положение в молекуле рецептора, могут предотвращать избыточное действие полного агониста, т.е. действуют частично как антагонисты. В этом случае употребляют понятие агонист/антагонист.
Высокой аффинностью могут обладать как агонисты, так и антагонисты рецептора. Агонист активирует рецептор, вызывая соответствующий физиологический эффект, в то время как антагонист, связываясь с рецептором, блокирует его и предотвращает развитие физиологического эффекта, выявляемого агонистами. Примером антагонистов могут служить нейролептики, которые предотвращают эффекты дофамина на уровне дофаминового рецептора.
При связывании лиганда с рецептором происходит изменение конфигурации последнего (рис. 7).
Многие вещества, как эндогенные, так и экзогенные, реагируют не с одним, а с несколькими типами рецепторов — "семейством" их, которое подразделяется на отдельные типы. Примером могут служить многие нейротрансмиттеры, реагирующие с несколькими типами специфических рецепторов (например, Д1—Д5-типы дофаминовых рецепторов). Существование нескольких рецепторов к одному лиганду носит название гетерогенности рецепторов.
Представление о функции рецепторов было бы неполным, если не представить внутриклеточные процессы, развивающиеся после связывания рецептора соответствующим веществом, и механизмы, обеспечивающие трансформацию внешнего сигнала в процессы, приводящие к появлению нервного импульса. Связывание лиганда с рецептором может приводить либо непосредственно к открытию (или закрытию) соответствующих ионных каналов (см. рис. 7), либо к активации вторичных мессенджерных систем (в качестве первичного мессенджера рассматривается вещество, реагирующее с рецептором).
Первые упоминания о вторичных мессенджерных системах появились в связи с работами E . Sutherland и соавт. (1950), которые показали, что адреналин стимулирует гликогенез путем увеличения концентрации циклического аденозинмонофосфата (цАМФ) в клетке. Оказалось, что этот вторичный мессенджер опосредует и другие клеточные реакции. В дальнейшем была выявлена связь действия цАМФ с активацией белковых киназ — ферментов, фосфорилирующих белки, что приводит к изменению их структуры и активности.


Позднее были открыты и другие вторичные мессенджеры. Сейчас выделяют среди них 3 класса: 1) циклические нуклеотиды (цАМФ, циклический гуанозинмонофосфат — цГМФ); 2) ионы кальция (Са 2+ ); 3) метаболиты фосфолипидов — инозитол-1,4,5-трифосфат (1Р 3 ), диглицерин (ДАГ), арахидоновую кислоту. В отличие от других вторичных мессенджеров Са 2+ транспортируется в нейрон из внутриклеточного пространства.
Мембраны нейрона содержат специализированные трансмембранные белки, которые формируют ионные каналы не только для Са 2+ , но и для других ионов, концентрация которых по обе стороны мембраны влияет на изменение мембранного потенциала. Происходят поляризация и деполяризация мембраны, т.е. изменение трансмембранного потенциала. Наибольшее значение в этих процессах имеют ионные каналы для натрия ( Na + ), калия (К + ), хлора (С1 - ) и кальция (Са 2+ ) .
Ацетилхолин является первым биологически активным веществом, которое было идентифицировано как нейромедиатор. Он высвобождается в окончаниях холинергических парасимпатических и симпатических волокон. Процесс освобождения медиатора является кальцийзависимым. Инактивация медиатора происходит с помощью фермента ацетилхолинэстеразы. Ацетилхолин оказывает свое воздействие на органы и ткани посредством специфических холинорецепторов. Действие ацетилхолина на пост-синаптическую мембрану постганглионарных нейронов может быть воспроизведено никотином, а действие ацетилхолина на исполнительные органы — мускарином (токсин гриба мухомора). На этом основании холинорецепторы разделили на Н-холинорецепторы (никотиновые) и М-холинорецепторы (мускариновые). Однако и эти виды холинорецепторов не однородны.
Н-холинорецепторы в периферических отделах вегетативной нервной системы расположены в ганглионарных синапсах симпатического и парасимпатического отделов, в каротидных клубочках и хромаффинных клетках мозгового слоя надпочечников. Возбуждение этих холинорецепторов сопровождается соответственно облегчением проведения возбуждения через ганглии, что ведет к повышению тонуса симпатического и парасимпатического отделов вегетативной нервной системы; повышением рефлекторного возбуждения дыхательного центра, в результате чего углубляется дыхание; повышением секреции адреналина. Вышеперечисленные Н-холинорецепторы блокируются веществами типа бензогексония (ганглиоблокаторами), но не реагируют на курареподобные вещества (d-тубокурарин), которые блокируют Н-холинорецепторы, локализованные на клетках скелетных мышц (в нервно-мышечном синапсе). В свою очередь, Н-холинорецепторы скелетных мышц не чувствительны к ганглиоблокаторам. В связи с этим Н-холинорецепторы подразделили на Н-холинорецепторы ганглионарного типа (Нн-холинорецепторы) и мышечного типа (Нм-холинорецепторы).
М-холинорецепторы также подразделяются на несколько типов: М1-, М2- и М3-холинорецепторы. Но все они блокируются атропином. М1-холинорецепторы находятся на обкладочных клетках желудочных желез и их возбуждение приводит к усилению секреции соляной кислоты. М2-холинорецепторы располагаются в проводящей системе сердца. Возбуждение этих рецепторов приводит к понижению концентрации цАМФ, открытию каливых каналов и увеличению тока К + , что приводит к гиперполяризации и тормозным эффектам: брадикардии, замедлению атриовентрикулярной проводимости, ослаблению сокращений сердца, понижению потребности сердечной мышцы в кислороде. М3-холинорецепторы локализованы в основном в гладких мышцах некоторых внутренних органов и экзокринных железах. Взаимодействие ацетилхолина с этими рецепторами приводит к активации натриевых каналов, деполяризации, формированию ВПСП, вследствие чего клетки возбуждаются и происходит сокращение гладких мышц и выделение соответствующих секретов. Возбуждение этих рецепторов в гладких мышцах бронхов, кишечника, мочевого пузыря, матки, круговой и цилиарной мышцах глаза приводит соответственно к бронхоспазму, усилению перистальтики кишечника, желудка при расслаблении сфинктеров, сокращению мочевого пузыря, матки, сужению зрачка и спазму аккомодации. Возбуждение М3-холинорецепторов экзокринных желез вызывает слезотечение, усиление потоотделения, выделение обильной бедной белком слюны, бронхорею, выделение желудочного сока. Имеются также внесинаптические М3-холинорецепторы, которые располагаются в эндотелии сосудов, где они ассоциированы с сосудорасширяющим фактором — окисью азота. Их возбуждение приводит к расширению сосудов и понижению артериального давления.
Норадреналин обеспечивает химическую передачу нервного импульса в норадренергических синапсах вегетативной нервной системы. Норадреналин относится к катехоламинам. Он синтезируется из аминокислоты тирозина в области пресинаптической мембраны адренергического синапса. В хромаффинных клетках надпочечников этот процесс продолжается, в результате чего образуется адреналин (тирозин-ДОФА-дофамин-норадреналин-адреналин). Инактивация норадреналина происходит с помощью ферментов катехол-о-метилтрасферазы (КОМТ) и моноаминоксидазы (МАО), а также путем обратного захвата нервными окончаниями с последующим повторным использованием. Частично Норадреналин диффундирует в кровеносные сосуды.
α -Адренорецепторы (постсинаптические) в основном локализованы в гладких мышцах сосудов кожи, слизистых и органов брюшной полости, а также в радиальной мышце глаза, гладких мышцах кишечника, матки, семявыносящих протоков, семенных пузырьках, капсуле селезенки, сфинктерах пищеварительного тракта и мочевого пузыря, пиломоторах. Возбуждение α1-адренорецепторов приводит к сужению радиальной мышцы глаза и расширению зрачка (мидриаз), сужению соответствующих сосудов и повышению АД, сокращению капсулы селезенки и выбросу депонированной крови, сокращению сфинктеров пищеварительного тракта и мочевого пузыря, расслаблению гладких мышц кишечника и снижению его перистальтики и т.д.
Среди α2-адренорецепторов выделяют пре-, пост- и внесинаптические. Возбуждение пресинаптических α2-адренорецепторов по механизму отрицательной обратной связи уменьшает выделение норадреналина при его избытке в синаптической щели. Постсинаптические α2-адренорецепторы находятся в бета-клетках поджелудочной железы. Их возбуждение вызывает угнетение выброса инсулина в кровь. Внесинаптические α2-адренорецепторы обнаружены преимущественно на мембране тромбоцитов, эндотелии некоторых сосудов, в жировых клетках. Возбуждение этих рецепторов вызывает сужение сосудов, агрегацию тромбоцитов, угнетение липолиза.
Β1-адренорецепторы (постсинаптические) выявлены в основном в проводящей системе сердца и гладкой мышце кишечника. Их возбуждение приводит к увеличению частоты сердечных сокращений, повышению проводимости и сократимости сердечной мышцы, увеличению потребности сердца в кислороде, понижению тонуса и моторной активности кишечника.
Стимуляция пресинаптических β2-адренорецепторов по механизму положительной обратной связи вызывает выделение норадреналина при его недостатке в синаптической щели. Постсинаптические β2-адренорецепторы расположены в основном в эндотелии сосудов скелетных мышц, головного мозга, легких, коронаров, а также в гладкой мускулатуре бронхов, матки и на гепатоцитах. Их возбуждение вызывает расширение соответствующих сосудов и понижение АД, расслабление бронхов и матки, усиление в печени гликогенолиза за счет активации цАМФ-зависимой фосфорилазы и повышение в крови сахара. Β3-Адренорецепторы находятся в жировых клетках. Их стимуляция приводит к активации липолиза.
В гладкой мышце артерий скелетных мышц содержатся α1- и β2-адренорецепторы. Возбуждение α1- адренорецепторов приводит к сужению артериол, а возбуждение β2-адренорецепторов — к их расширению. В гладких мышцах кишечника находятся α1- и β1-адренорецепторы, возбуждение и тех и других приводит к расслаблению мышц. Эффекты норадреналина реализуются через α- и β-адренорецепторы, однако норадреналин обладает большим сродством к а-адренорецепторам. Адреналин выполняет гормональную функцию в организме, его эффекты реализуются через кровь посредством возбуждения внесинаптических α- и β-адренорецепторов, но большим сродством он обладает к β-адренорецепторам. Реакция органа на норадреналин и адреналин зависит от преобладания α- или β-адренергического действия.
Дофамин осуществляет химическую передачу нервных импульсов не только в дофаминергических синапсах ЦНС, но и во вставочных нейронах симпатических ганглиев и во внутриорганном отделе вегетативной нервной системы. В дофаминергических нейронах биосинтез катехоламинов заканчивается на дофамине. Инактивация дофамина осуществляется ферментами КОМТ и МАО, а также путем обратного нейронального захвата. Периферические дофаминовые рецепторы (Д-рецепторы) изучены недостаточно. Д-рецепторы выявлены на гладкомышечных клетках кишечника, сосудов почек, аорты, паращиторидных железах, канальцах почек. Возбуждение этих рецепторов приводит к расслаблению гладких мышц, понижению тонуса кишечника, расширению соответствующих сосудов, повышению высвобождения паратгормона, усилению выделения натрия и воды. Дофаминовые рецепторы выявлены также в надпочечниках и поджелудочной железе, но пока не идентифицированы. Эти рецепторы регулируют секрецию панкреатического полипептида, бикарбонатов и альдостерона.
АТФ может играть роль не только макроергического соединения, но и медиатора. Местом его локализации является пресинап-тические терминали эффекторных нейронов внутриорганного отдела вегетативной нервной системы. Эта передача получила название пуринергической, так как при стимуляции этих окончаний выделяются пуриновые продукты распада — аденозин и инозин. Действие АТФ проявляется в основном в расслаблении гладкой мускулатуры. Пуринергические нейроны являются, по-видимому, главной антагонистической тормозной системой по отношению к холинергической возбуждающей системе. Пуриноре-Цепторы представлены двумя группами: Р1, и Р2 Р1-рецепторы более чувствительны к продукту распада АТФ — аденозину, Р2-рецепторы — к самому АТФ. Р1-рецепторы преобладают в сердечнососудистой системе, трахее, мозге. Специфичными антагонистами Р1-рецепторов являются метилксантины, например, алкалоиды кофе и чая — кофеин и теофиллин. Р2-рецепторы располагаются в основном в органах желудочно-кишечного тракта и мочеполовой системы. Специфическим блокатором Р2-рецепторов служит хинидин.
Одним из медиаторов внутриорганного отдела вегетативной нервной системы является серотонин, или 5-окситриптамин, который выполняет также медиаторную функцию в центральных образованиях. Серотонин оказывает свое воздействие путем взаимодействия со специфическими серотониновыми рецепторами. Периферические S1-рецепторы (или 5-НТ1) в основном обнаружены в гладких мышцах желудочно-кишечного тракта, сосудах скелетных мышц и сердца, проводящей системе сердца. Их возбуждение сопровождается спазмом гладких мышц кишечника, вазодилатацией, тахикардией. S2-рецепторы (5-НТ2) находятся в гладких мышцах стенок сосудов, бронхов, на тромбоцитах. При их стимуляции возникает спазм сосудов, за исключением сосудов скелетных мышц и сердца, и повышается АД, увеличивается агрегация тромбоцитов. S3-рецепторы (5-НТ3) локализуются в гладких мышцах, вегетативных ганглиях. Посредством взаимодействия с этими рецепторами серотонин осуществляет регуляцию сократительной способности гладких мышц и усиление освобождения ацетилхолина в терминалях вегетативных нервов.
Роль медиатора в вегетативной нервной системе может играть гистамин. Наибольшее количество его находится в постганглионарных симпатических волокнах. Инактивация гистамина осуществляется ферментом диаминоксидазой. Периферические гистаминовые рецепторы встречаются во всех органах и тканях организма. Известно два класса гистаминовых рецепторов: Н1 и H2. H1-рецепторы локализуются в гладкой мускулатуре бронхов, желудочно-кишечного тракта, сосудов, в сердце (атриовентрикулярный узел). Возбуждение Н1-рецепторов сопровождается спазмом бронхов, повышением тонуса и перистальтики кишечника, сужением крупных сосудов, но расширением артериол, венул и развитием, в общем итоге, гипотензии, повышением сосудистой проницаемости, уменьшением времени проведения по атриовентрикулярному узлу, тахикардией, увеличением образования простагландинов. Н2-рецепторы обнаружены преимущественно в слизистой желудка и 12-перстной кишки, слизистой бронхов, на базофилах, на Т-супрессорах. Возбуждение Н2-рецепторов приводит к повышению секреции кислоты в желудке и секреции бронхиальных желез, уменьшению высвобождения гистамина базофилами, стимуляции Т-супрессоров.
Функцию медиаторов синаптической передачи во внутриорганном отделе вегетативной нервной системы выполняют и некоторые аминокислоты, регуляторные нейропептиды, простагландины и другие биологические активные вещества. Аспарагиновая и глутаминовая кислоты являются медиаторами возбуждающего типа, гамма-аминомасляная кислота (ГАМК) — медиатором тормозного типа. В результате взаимодействия ГАМК с ГАМК-рецепторами происходит открытие каналов для ионов хлора, что обусловливает гиперполяризующее действие ГАМК. Периферическое действие ГАМК практически не исследовано, хотя имеются сведения о способности ГАМК изменять состояние внутренних органов. Например, ГАМК-рецепторы обнаружены в кишечнике. различают ГАМК-а и ГАМК-в-рецепторы. Возбуждение ГАМК-а-рецепторов приводит к сокращению гладких мышц кишечника, возбуждение ГАМК-в, наоборот, — расслаблению. ГАМК-в-рецепторы выявлены также в предсердиях и сосудах. Их возбуждение приводит к снижению сократительной активности предсердий и тонуса сосудов. ГАМК способна влиять на освобождение других медиаторов из пресинаптических окончаний: ацетилхолина, дофамина, серотонина.
Представителем регуляторных нейропептидов является субстанция Р. Периферические рецепторы к субстанции Р обнаружены на гладкомышечных клетках кишки (SP-P-рецепторы) и на клетках мочевого пузыря, семявыносящих протоков (SP-E-рецепторы).
В волокнах блуждающего нерва содержится большое количество простагландинов.
Вегетативные (автономные) рефлексы
Процессы в вегетативной и соматической нервных системах тесно связаны.
Различают висцеро-висцеральные, висцеросоматические, висцеросенсорные рефлексы. Классическим примером висцеро-висцерального рефлекса является рефлекс Гольца, показывающий, что механическое раздражение брыжейки вызывает замедление частоты сердечных сокращений. Разновидностью висцеро-висцерального рефлекса является аксонрефлекс, например, возникновение сосудистой реакции при раздражении кожных болевых рецепторов.
К висцеросоматическим рефлексам относятся торможение общей двигательной активности организма при раздражении хемо- и механорецепторов каротидной зоны, а также сокращение мышц брюшного пресса или подергивание конечностей при раздражении рецепторов пищеварительного тракта.
При висцеросенсорных рефлексах в ответ на раздражение вегетативных чувствительных волокон возникают не только реакции во внутренних органах, но и изменяется соматическая чувствительность. Для их вызова необходимо продолжительное и сильое воздействие. Зона повышенного восприятия обычно огранивается участком кожи, иннервируемым сегментом, к которому поступают импульсы от раздражаемого висцерального органа. В клинике имеют определенное значение висцеродермальные рефлексы. Вследствие сегментарной организации вегетативной и соматической иннервации при заболеваниях внутренних органов на ограниченных участках кожи возникает повышение тактильной и болевой чувствительности. Эти боли называются отраженными, а области, в которых они появляются, — зонами Захарьина—Геда.
При раздражении некоторых областей поверхности тела возникают сосудистые реакции и изменения функций определенных висцеральных органов. Это дермовисцеральный рефлекс, который является разновидностью соматовисцерального рефлекса. На этом основана рефлексотерапия.
Читайте также:
