
Непрямой путь обработки двигательного сигнала базальными ганглиями
Роль базальных ганглиев в двигательной регуляции
Базальные ганглии (анатомически обособленная группа парных подкорковых структур) - это глубинные ядра головного мозга . Вместе с другими ядрами промежуточного и среднего мозга они оказывают на двигательную активность иное влияние, чем мозжечок . В отличие от последнего у базальных ганглий нет входа от спинного мозга , но есть прямой вход от коры больших полушарий . Их основное влияние направлено посредством таламуса на двигательные области коры мозга . Кроме того, базальные ганглии участвуют в эмоциональных (аффективных) функциях и познавательных (когнитивных) функциях .
Базальные ганглии оказывают влияние прежде всего на кору больших полушарий .
Поражения базальных ядер обусловливают нарушения движений, поскольку они играют важную роль в деятельности латеральной системы двигательных путей . Это подтверждается характером симптомов при заболеваниях базальных ганглиев . Однако эти ганглии связаны и с медиальной системой двигательных путей ; их заболевания сопровождаются нарушениями позы и тонуса проксимальной мускулатуры .
Базальные ядра облегчают запускаемые корой движения и подавляют лишние сопутствующие движения. Стриатум получает соматотопически организованные проекции почти от всех зон коры. Эти проекции организованы в виде параллельных путей, которые начинаются от лобных областей , постцентральных (соматосенсорных) областей , прецентральных (моторных) областей и теменно-височно-затылочных областей . В то же время прямое отношение к регуляции движений имеют только глутаматергические, возбуждающие проекции от моторной и соматосенсорной коры .
Нейроны стриатума посылают волокна к структурам, формирующим главный эфферентный путь базальных ядер - к сетчатой части черной субстанции и медиальному бледному шару. Эти структуры, в свою очередь, образуют тормозные ГАМК ергические синапсы на переднем и латеральном вентральных ядрах таламуса , которые направляют к коре возбуждающие глутаматергические волокна .
Следовательно, при торможении нейронов сетчатой части черной субстанции и медиального бледного шара возбуждающее влияние таламуса на кору усиливается - что и способствует облегчению запускаемого корой движения.
Напротив, при возбуждении нейронов сетчатой части черной субстанции и медиального бледного шара возбуждающее влияние таламуса на кору подавляется - что приводит к торможению ненужного движения.
Прямого выхода на спинной мозг базальные ядра не имеют. Прямой путь от стриатума к сетчатой части черной субстанции и медиальному бледному шару образован тормозными ГАМКергическими волокнами. Функция их состоит в усилении возбуждающего влияния ядер таламуса на те отделы моторной коры, которые отвечают за нужное движение.
Непрямой путь организован сложнее. Функция его заключается в подавлении возбуждающего влияния таламуса на другие отделы моторной коры. Первое звено этого пути - тормозные ГАМКергические проекции стриатума на латеральный бледный шар. Латеральный бледный шар посылает тормозные ГАМКергические волокна к субталамическому ядру. Выходы субталамического ядра представлены возбуждающими глутаматергическими волокнами - часть их возвращается к латеральному бледному шару, другие направляются к сетчатой части черной субстанции и медиальному бледному шару.
Таким образом, если активация прямого пути от стриатума усиливает возбуждение моторной коры, то активация непрямого пути - ослабляет.
Активность этих двух путей регулирует компактная часть черной субстанции, посылающая к стриатуму дофаминергические волокна. Эти волокна возбуждают прямой эфферентный путь стриатума (через нейроны с D1-рецепторами ) и тормозят - непрямой (через нейроны с D2-рецепторами ). Стриатум посылает также тормозные ГАМКергические волокна к компактной части черной субстанции, образуя тем самым с ней отрицательную обратную связь. Наконец, активность стриатума модулируют его холинергические вставочные нейроны - антагонисты дофаминергических нейронов.
К базальным ганглиям относят комплекс нейронных узлов серого вещества, которые располагаются в белом веществе больших полушарий головного мозга. Эти образования называют стриополитарной системой. Относится хвостатое ядро, скорлупа – вместе они образуют полосатое тело. Бледный шар на разрезе состоит из 2х сегментов – наружного и внутреннего. Наружный сегмент бледного шара имеет общее происхождение с полосатым телом. Внутренний сегмент развивается из серого вещества промежуточного мозга. Эти образования имеют тесную связь с субталамическими ядрами промежуточного мозга, с черной субстанцией среднего мозга, которая состоит из двух частей – вентральной части(сетчатой) и дорсальной(компактная).
Нейроны компактной части вырабатывают дофамин. А сетчатая часть черной субстанции по строению и функциям напоминает нейроны внутреннего сегмента бледного шара.
Черная субстанция образует связи с передним вентральным ядром зрительного бугра, бугорками четверохолмия, с ядрами моста и двухсторонние связи с полосатым телом. Эти образования получают афферентные сигналы и сами формируют эфферентные пути. Чувствительные пути к базальным ганглиям идут от коры больших полушарий и главный афферентный путь начинается от моторной и премоторной зоны коры.
Корковое поля 2,4,6,8. Эти пути идут к полосатому телу и бледному шару. Имеется определенная топография проекции мышц дорсальной части скорлупы представлены мышцы ног, рук, а в вентральной части – рта и лица. От сегментов бледного шара идут пути к зрительному бугру переднем вентральному и вентролатеральному ядрам, от которых информация будет возвращаться в кору.
Большое значение играют пути к базальным ядрам от зрительных бугров. Обеспечивают получение сенсорной информации. К базальным ядрам также через зрительный бугор передаются влияния от мозжечка. Также имеются чувствительные пути к полосатому телу от черной субстанции. Эфферентные пути представлены связями полосатого тела с бледными шарами, с черной субстанцией, ретикулярной формацией ствола мозга, от бледного шара идут пути к красному ядру, к субталамическим ядрам, к ядрам гипоталамуса и зрительных бугров. На подкорковом уровне сложные кольцевые взаимодействия.
Связи коры больших полушарий, зрительного бугра базальные ганглии и снова кора формируют два пути: прямой(обеспечивает облегчение прохождения импульсов) и непрямой(тормозной)
Непрямой путь. Оказывает тормозящее действие. Этот путь тормозной идет от полосатого тела к наружному сегменту бледного шара и полосатое тело тормозит наружный сегмент бледного шара. Наружный сегмент бледного шара тормозит Люисово тело, которое в норме оказывает возбуждающие действие на внутренний сегмент бледного шара. В этой цепочке есть два последовательных торможения.
При прямом пути кора больших полушарий оказывает на полосатое тело, тормозящее действие на внутренний сегмент бледно шара, происходит растормаживание.
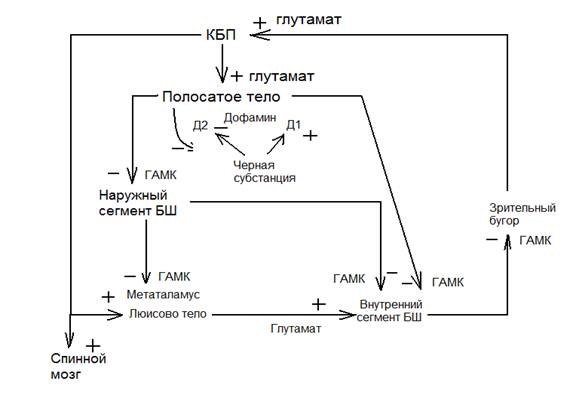
Черная субстанция(вырабатывает дофамин) В полосатом теле есть 2 вида рецепторов Д1- возбуждающие, Д2 – тормозящие. Полосатое тело с черной субстанцией два тормозящих пути. Черная субстанция тормозит полосатое тело дофамином, а полосатое тело черную субстанцию ГАМК. Высокое содержание меди в черной субстанции, синем пятне ствола мозга. Возникновение стриополитарной системы было необходимо для совершения перемещения тело в пространстве - плаванье, ползанье, полет. Эта система образует связь с подкорковыми двигательными ядрами(красное ядро, покрышка среднего мозга, ядра ретикулярной формации, вестибулярные ядра) От этих образований – нисходящие пути в спинной мозг. Все это вместе образует экстрапирамидную систему.
Двигательная активность реализуется через пирамидную систему – нисходящие пути. Каждое полушарие связано с противоположной половиной тела. В спинном мозге с альфа моторными нейронами. Через пирамидную систему реализуются все наши желания. Она работает с мозжечком, экстрапирамидной системы и выстраивается несколько контуров – кора мозжечка, кора, экстрапирамидная система. Зарождение мысли возникает в коре. Для того, чтобы его совершить необходим план движения. Который включает в себя несколько компонентов. Они связываются в один образ. Для этого нужны программы. Программы быстрых движений – в мозжечке. Медленных – в базальных ганглиях. Кора выбирает необходимые программы. Она создает единственную общую программу, которая будет реализовано через спинальные пути. Чтобы сделать бросок мяча в кольцо нам нужно принять определенную позу, распределить тонус мышц – это все на подсознательном уровне – экстрапирамидная система. Когда все будет готово произойдет само движение. Стриополитарная система может обеспечивать стереотипные заученные движения – ходьба, плаванье, езда на велосипеде, но только когда они заучены. При выполнение движение стриополитарная система определяет масштаб движений – амплитуда движений. Масштаб определяется стриополитарной системой. Гипотония-пониженный тонус с гиперкинезом - повышенная двигательная активность.
Базальные ганглии(Стрио - Паллидарная система)
Симптомы поражения базальных ганглиев
К числу гиперкинезов (сопровождаются снижением тонуса мышц) относятся
-Хорея - связана с дегеративными поражениями хвостатого ядра и проявляется в возникновении быстрых танцующих движений. Возникает богатая мимика, непрерывная игра пальцами рук, причмокивание, развивается в результате ревматического поражения. Все движения непроизвольные
-Атетоз - обусловлен поражением скорлупы и бледного шара и характеризуется медленными, извивающимися движениями – червеобразными движениями, которые начинаются с дистальных отделов конечностей и постепенно пермещаются на проксимальные.
-Баллизм - размашистые движения верхних и нижних конечностей
Гиперкинез в сочетании с гипертонией – Болезнь Паркинсона(уменьшение выработки дофамина в нейронах компактной части черной субстанции. Черная субстанция оказывает тормозящее действие на полосатое тело. Таким образом снижается содержание дофамина в полосатом теле. Симптомы – снижение дофамина до 50 % от нормы. Одновременно снижается содержание и норадреналина в гипоталамусе.). Симптомы – мелкие движения пальцев рук, мимика, гипертония(повышается тонус мышц, в основном сгибателей. Поза – руки приведены к туловищу, колени согнуты, голова прижата. Дрожание в покое – Тренор, маскообразное лицо, замедленная речь). Симптом складного ножа –попытка согнуть руку в локтевом суставе – сначала большое сопротивление, а потом легко. Симптом зубчатого колеса – периодическая смена повышения и снижения тонуса.
Вводят препараты Эльдофа – могут проникать через гемоэнцефалический барьер и превращаются в дофамин. Помогают блокаторы, которые разрушает норажреналин и дофамин. Есть попытки вживления клеток, взятых у мертвых новорожденных из черной субстанции
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Структурно-функциональная организация базальных ганглиев.Базальные ганглии (стриопаллидарная система)– это совокупность трех парных образований, расположенных в конечном мозге на основании больших полушарий. Включает в себя 3 парных образования (см.рис 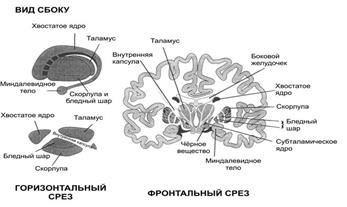
.12):
Рис.12. Анатомия базальных ядер.
бледный шар – состоит из наружного и внутреннего сегмента, является наиболее филогенетически древней частью;
- полосатое тело – состоит из хвостатого ядра и скорлупы;
- ограда – расположена между скорлупой и островковой корой, является наиболее молодой частью.
Установлены внутренние связи между базальными ганглиями. За их счет компоненты базальных ганглиев тесно взаимодействуют и образуют единую стриопаллидарную систему. Кроме того, базальные ганглии имеют многочисленные афферентные и эфферентные связи с другими структурами мозга.
Афферентные связи поступают (см.рис.13) в базальные ядра от всех областей коры прямо и через таламус, от неспецифических ядер таламуса, от черного вещества.
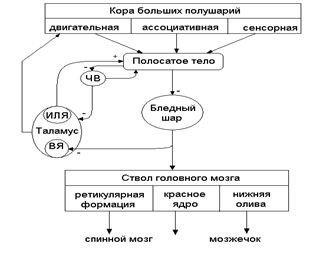
Рис. 13. Схема основных афферентных и эфферентных связей базальных ядер:
ЧВ – черное вещество; ВЯ – вентральные ядра; ИЛЯ – интраламинарное ядро;
Имеются три эфферентных выхода:
1. от полосатого тела тормозящие пути идут к бледному шару, от бледного шара к двигательным ядрам таламуса и от них к двигательной коре (поля 4 и 6);
2. часть эфферентных волокон из бледного шара и полосатого тела идет к ретикулярной формации и красному ядру ствола мозга и далее в спинной мозг, а также через нижнюю оливу в мозжечок;
3. от полосатого тела тормозящие пути идут также к черному веществу и далее к ядрам таламуса.
В последнее время было выявлено, что связи базальных ганглий образуют параллельные функциональные петли (см.рис.14): скелетно-моторную и глазодвигательную петли.
Скелетно-моторная петля служит для регуляции таких параметров движения как амплитуда, сила и направление. Она соединяет премоторную, двигательную и соматосенсорную области коры со скорлупой базальных ядер, далее импульсация идет в бледный шар и черное вещество среднего мозга и через двигательное ядро (вентролатеральное ядро) таламуса возвращается в премоторную кору.
Глазодвигательная петля участвует в регуляции движений глаз (например, скачкообразных движений глаз – саккад). Она соединяет области коры, контролирующие направление взгляда (поле 8 лобной коры и поле 7 теменной коры), с хвостатым ядром. Оттуда импульсы поступают в бледный шар и черное вещество из которых она поступает в ассоциативное медиадорсальное и переднее релейное вентральное ядро таламуса и возвращается в лобное глазодвигательное поле 8.
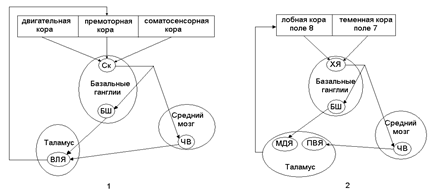
Рис.14. Схема функциональных петель, проходящих через
базальные ганглии (по Р.Шмидту, 1996, с изменениями)
1 – скелетно-моторная петля; 2 – глазодвигательная петля;Ск – скорлупа; БШ – бледный шар; ЧВ – черное вещество; ВЛЯ – вентролатеральное ядро; ХЯ - хвостатое ядро; МДЯ – медиодорсальное ядро; ПВЯ – переднее вентральное ядро
Функции базальных ядер.Основными функциями базальных ядер являются:
1. Регуляция двигательной активности. Важную роль базальные ядра играют при переходе от замысла движения (фазы подготовки к движению) к выбранной программе действия (фазе выполнения).
2. Участие в интеграции (объединении) тонических рефлексов и в обеспечении позы.
3. Участие, наряду с мозжечком, в выработке сложных двигательных программ.
4. Участие в контроле таких параметров движения как сила, амплитуда, скорость и направление движения.
Функции полосатого тела.Полосатое тело состоит из более крупных нейронов с длинными отростками, которые выходят за пределы стриапаллидарной системы. Полосатое тело участвует в регуляции мышечного тонуса, уменьшая его, а также в организации и регуляции движений. Стимуляция полосатого тела вызывает:
- простые двигательные реакции;
- торможение активности коры, подкорковых образований, безусловных и условных рефлексов;
- торможение поведенческих реакций (пищедобывательной и ориентировочной).
Полосатое тело оказывает на бледный шар тормозящее влияние (медиатор ГАМК), снижая активность бледного шара, и тем самым, снижая двигательную активность.
Полосатое тело имеет двухсторонние связи с черным веществом среднего мозга. Нейроны полосатого тела оказывают тормозное влияние (медиатор ГАМК) на нейроны черного вещества. Нейроны черного вещества секретируют медиатор дофамин, который посредством аксонного транспорта поступает к базальным ганглиям. При резком снижении концентрации дофамина в хвостатом ядре развивается болезнь Паркинсона, которая характеризуется сильным дрожанием пальцев, губ, головы и других частей тела, ригидностью мышц и т.д.
Между полосатым телом и таламусом также имеются двухсторонние связи. Через черное вещество полосатое тело оказывает на таламус тормозящее влияние и получает возбуждающие афферентные импульсы от субталамического ядра.
На двигательную кору полосатое тело также оказывает тормозное влияние, сдерживая ненужную в данный момент степень свободы движения, и тем самым обеспечивая четкость и направленность двигательных реакций.
Поражение полосатого тела сопровождается возникновением так называемых гиперкинезов. Гиперкинезы – это избыточные, насильственные, непроизвольные движения. Движения у больных становятся размашистыми, избыточным, широко амплитудными. Больные не могут усидеть на одном месте, все время вертятся, меняют положение тела и т.д. Кроме того, могут наблюдаться следующие симптомы:
· Тремор – подергивание конечностей;
· Ригидность мышц – повышение мышечного тонуса;
· Баллизм – крупноразмашистый гиперкинез конечностей;
· Атетоз – больше наблюдается в дистальных отделах конечностей – в кистях и пальцах рук; проявляется медленными, извивающимися, червеобразными движениями;
Скорлупа отвечает за организацию пищевого поведения. При раздражении скорлупы появляется слюноотделение и изменение дыхания, при поражении наблюдаются трофические изменения в коже.
Функции бледного шара. Бледный шар оказывает модулирующее влияние на двигательную кору, мозжечок, ретикулярную формацию и красное ядро. Его функцией является провоцирование ориентировочной реакции, движения конечностей, активация пищевого поведения (жевание, глотание).
Ограда – функция мало изучена. Имеет двухсторонние связи с лобной, затылочной, височной корой, обонятельной луковицей, таламусом и другими базальными ядрами. При раздражении ограды вызываются моторные реакции со стороны пищеварительного тракта (жевание, глотание, рвотные движения), ориентировочные реакции. При повреждении – теряется способность говорить.
Дата добавления: 2015-09-15 ; просмотров: 8668 . Нарушение авторских прав
Функция мозжечка состоит, в первую очередь, в формировании двигательных программ, необходимых для поддержания равновесия тела, регуляции силы мышц и объема совершаемых с их участием движений. Он не является составной частью иерархической сети двигательных систем, но имеет связи со всеми ее уровнями: моторной корой, двигательными центрами ствола и спинным мозгом. Благодаря этим связям мозжечок может сравнивать моторные программы коры с их исполнением, а затем корректировать движения, не соответствующие намеченной программе. Мозжечок способен накапливать опыт двигательной активности, сохраняя его в долговременной моторной памяти.
В мозжечке имеются три области, различающиеся спецификой своих соединений с головным и спинным мозгом и выполняемыми функциями:
1) вестибулоцеребеллум (червь с примыкающими к нему областями старой коры);
2) спиноцеребеллум (примыкающие к червю медиальные области полушарий мозжечка);
3) цереброцеребеллум, включающий в себя латеральные области полушарий мозжечка.
Вестибулоцеребеллум (функция сохранения равновесия и управления движениями глаз) получает зрительную и слуховую информацию, входные сигналы от вестибулярных ядер, ядер тройничного нерва и спинного мозга. Основной эфферентный выход к вестибулярным ядрам ствола.
Спиноцеребеллум (управление движениями конечностей) получает афферентную информацию от проприоцепторов посредством спиномозжечковых трактов; эфферентный выход к первичной и вторичной моторной коре.
Цереброцеребеллум (формирование двигательных программ) получает афферентную информацию о планирующемся движении при посредничестве ядер моста от сенсомоторных областей коры; основной эфферентный выход к первичной и вторичной моторной коре.
При выполнении запрограммированных произвольных движений выходная активность нейронов зубчатых ядер мозжечка регистрируется приблизительно на 10 мс раньше, чем она обнаружится в моторной коре. Это опережение имеет особенное значение при выполнении быстрых движений, когда из-за дефицита времени кора не может исправить ошибку. Мозжечок не может инициировать движение, он лишь обеспечивает необходимый баланс действующих мышц, причем его участие особенно необходимо в заключительной стадии движений, когда замысел сопоставляется с результатом. При мозжечковом поражении пациент, например, промахивается, когда его просят быстро прикоснуться к кончику своего носа указательным пальцем.
Клетки ядер мозжечка представляют собой обычные переключательные нейроны: в ответ на поступающую по коллатералям моховидных и лазающих волокон афферентную информацию они возбуждаются и отправляют эфферентные сигналы двигательным ядрам ствола. Клетки Пуркинье (кора мозжечка, медиатор ГАМК) ограничивают их активность таким образом, что выполнение одних моторных программ допускается, а других — отменяется в соответствии с характером афферентной информации и организацией связей между корой мозжечка и ядрами.
Эфферентное влияние зубчатых и промежуточных ядер мозжечка на сенсомоторную кору больших полушарий осуществляется при участии моторных вентролатеральных ядер таламуса, которые служат узловым пунктом нейронной сети, связывающей сенсомоторную кору не только с мозжечком, но и с базальными ганглиями. Аксоны таламических нейронов способны изменять активность пирамидных клеток, образующих кортикоспинальный путь, что позволяет мозжечку корректировать двигательные программы, создаваемые в моторных областях коры.
Еще одним пунктом переключения моторных систем мозга являются красные ядра, служащие источником образования руброспинальных путей. Здесь оканчиваются аксоны возбуждающих нейронов промежуточных ядер мозжечка и моторной коры, оказывающей тормозное влияние. От ядер шатра эфферентные волокна идут к вестибулярным ядрам и моторным ядрам ретикулярной формации ствола мозга, что позволяет мозжечку контролировать нисходящую активность вестибулоспинальных и ретикулоспинальных путей. Действие мозжечка на стволовые двигательные центры позволяет исправлять ошибки уже запущенной двигательной программы, если они возникают в процессе ее осуществления.
46. Участие базальных ганглиев в регуляции движений: прямой и непрямой пути. Роль дофамина и ацетилхолина в регуляции активности прямого и непрямого пути.
Полосатое тело (хвостатое ядро и скорлупа) функционируют как входные ворота в базальные ганглии: они получают возбуждающие (глутаматергические) сигналы от коры. Скорлупа получает входы от сенсорных и моторных областей, окружающих центральную борозду, поэтому её активность прямо связана с двигательной системой. Хвостатое ядро иннервируется фронтальной корой, и поэтому оно вовлечено в когнитивные процессы высокого порядка. Параллельное поступление той и другой информации обеспечивает роль базальных ганглиев в сознательной деятельности, эмоциях, а также в выполнении двигательных команд.
Прямой путь. ГАМК-ергические (главные) нейроны полосатого тела направляют свои аксоны во внутренний сегмент бледного шара, снижая её активность. Нейроны бледного шара относятся к числу тормозных, они освобождают ГАМК в синапсах на нейронах таламуса. Нейроны бледного шара тонически активны, что приводит к постоянному торможению прохождения сигнала от таламуса к коре. При поступлении возбуждающего сигнала от коры к полосатому телу увеличивается активность его главных тормозных нейронов, что, в свою очередь, приводит к снижению активности тормозных нейронов внутреннего сегмента бледного шара и растормаживанию ядер таламуса. В результате облегчается прохождение сигнала по таламокортикальному пути (облегчается таламическая активация коры).
Непрямой путь. ГАМК-ергические (главные) нейроны полосатого тела направляют свои аксоны к тормозным нейронам наружного сегмента бледного шара. В результате ограничения активности последних растормаживаются затормаживаемые ими нейроны субталамического ядра, и усиливается активация тормозных нейронов внутреннего сегмента бледного шара. Таким образом, в результате активации непрямого пути усиливается тормозное влияние внутреннего сегмента бледного шара на таламические ядра, и ограничивается прохождение сигнала по таламокортикальному пути (ограничивается таламическая активация коры).
Дофаминергические нейроны компактной дорсальной части ретикулярной формации образуют диффузную нигростриатную проекцию на нейроны полосатого тела. Эта проекция топографически не организована и предназначена для модуляции кортикостриатных глутаматергических переключений. При дегенерации дофаминергических нейронов черной субстанции нарушается необходимый баланс активности нейронов полосатого тела, что приводит к характерным двигательным нарушениям.
Дисфункция, вызванная потерей дофаминергических нейронов, проявляется в виде болезни Паркинсона и характеризуется мышечной ригидностью (значительным повышением тонуса мышц), резким ограничением сопутствующих движений, или акинезией, а также появлением крупноразмашистого тремора — дрожания конечностей во время покоя. Указанные нарушения двигательной функции (гипокинез) возникают в связи с уменьшением влияния дофаминергических нейронов и изменением баланса активности в пользу остальных нейронов полосатого тела.
Если же этот баланс нарушается вследствие поражения холинергических или ГАМКергических нейронов полосатого тела, то возникает другая разновидность двигательных расстройств, проявляющихся появлением непроизвольных и нежелательных для человека насильственных движений — гиперкинезов (н-р, хорея).
Базальными ганглиями называют скопления нейронов в базальных отделах переднего и среднего мозга, которые участвуют в регуляции движения. К базальным ганглиям относят следующие структуры:
• Полосатое тело (хвостатое ядро, скорлупа, прилежащее ядро).
• Бледный шар (часть чечевицеобразного ядра), который имеет наружную (латеральную) и внутреннюю (медиальную) части. Внутренняя часть продолжается к среднему мозгу в виде сетчатой части черной субстанции.
• Субталамическое ядро (СТЯ).
• Пигментная компактная часть черной субстанции.
• Чечевицеобразное ядро, которое образуют скорлупа и бледный шар.
Вставочные нейроны оказывают прямой модулирующий эффект на обе группы проекционных нейронов за счет влияния на пресинаптические терминали и изменения выброса глутамата (корково-стриарный путь) и дофамина (нигростриарный путь). В полосатом теле выделяют отдельные ядра, которые получают нервные волокна от нейронов коры или от ядер таламуса. Полосатое тело также подразделяют на функциональные области: стриосому и матрикс. Нейроны прямого и непрямого путей расположены в матриксе. Нейроны, находящиеся внутри стриосомы, получают нервные импульсы от лимбической коры и отдают проекции к компактной части черной субстанции; эти нейроны образуют путь, через который базальные ганглии оказывают свое влияние на лимбическую систему.
Можно выделить, как минимум, четыре пути, которые формируются в коре, направляются к базальным ганглиям, а затем возвращаются обратно в кору:
1. Двигательная петля — отвечает за выученные двигательные акты.
2. Когнитивная петля — отвечает за намерения и планирование движения.
3. Лимбическая петля — отвечает за эмоциональную окраску движения.
4. Глазодвигательная петля — отвечает за произвольные саккады.
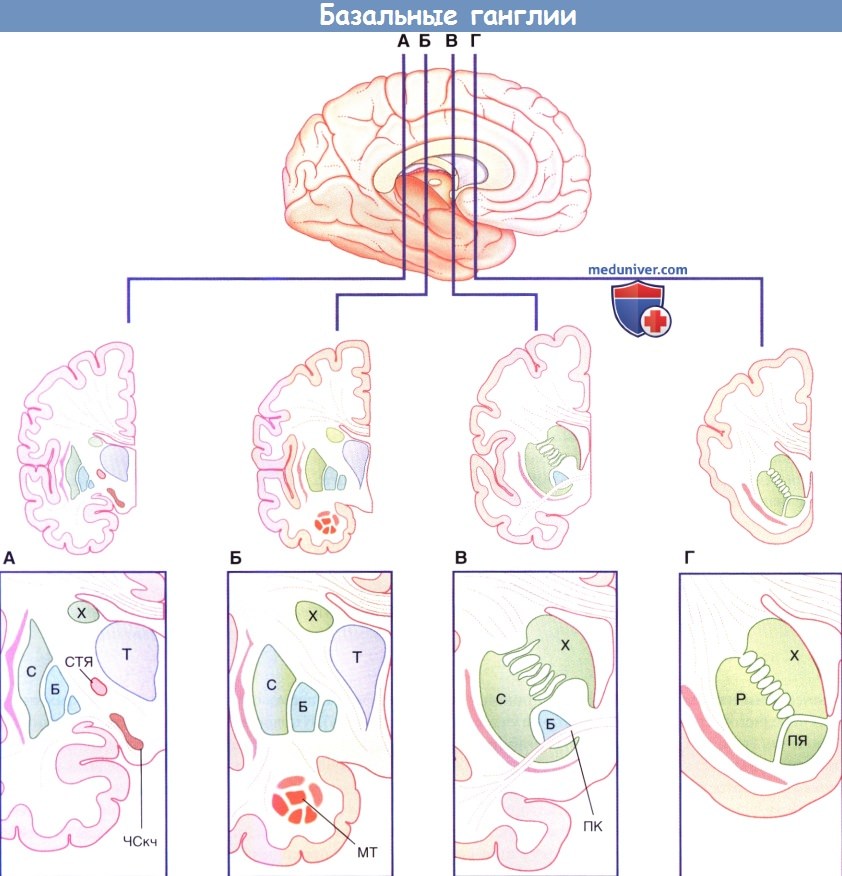
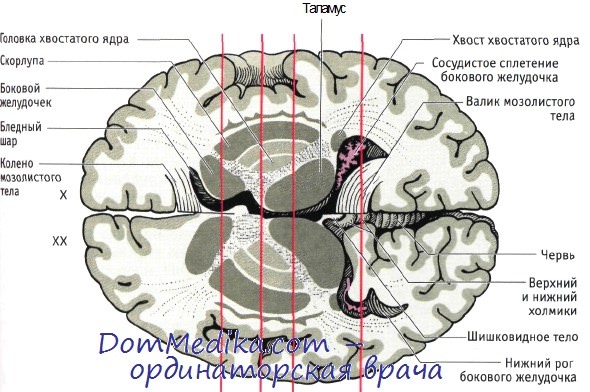
(А-Г). Четыре фронтальных среза головного мозга, вид сзади.
Вентральные отделы на рисунках снизу представлены в увеличенном виде.
ПЯ — прилежащее ядро; ПК — передняя комиссура; МТ — миндалевидное тело; X — хвостатое ядро; Б — бледный шар;
С — скорлупа; ЧСкч — компактная часть черной субстанции; СТЯ — субтапамическое ядро; Т — таламус.
а) Двигательная петля. Двигательная петля начинается в сенсомоторной коре, а возвращается обратно через полосатое тело, таламус и дополнительную моторную область (ДМО).
На рисунке ниже схематично изображено строение проводящих путей двигательной петли, в том числе задней части полосатого тела. Существует два пути. Прямой путь образован ядрами базальных ганглиев, таламуса и проходит через пять последовательных групп нейронов. Непрямой путь также включает СТЯ и состоит из семи групп нейронов. Отдельно от них проходят проекции от ВБШ к таламусу (чечевицеобразная петля и чечевицеобразный пучок), которые показаны на рисунке ниже.
Все проекции, начинающиеся от коры, исходят от пирамидных клеток и являются возбуждающими (глутаматергическими). Это же касается волокон, идущих от таламуса к ДМО. Волокна, начинающиеся от полосатого тела и обеих частей бледного шара, исходят от средних шипиковых нейронов и оказывают тормозящее влияние. Они являются ГАМК-ергическими, а также содержат нейропептиды с неустановленной в настоящее время функцией.
Нигростриарный путь идет от компактной части черной субстанции до полосатого тела, где на проекционных нейронах он образует два вида синапсов. Волокна, образующие синапсы с нейронами прямого пути,— стимулирующие, на дендритных шипиках расположены дофаминергические рецепторы типа 1 (D1). Волокна, образующие синапсы с нейронами непрямого пути,— тормозящие, их действие опосредовано до-фаминергическими рецепторами типа 2 (D2). Холинергические вставочные нейроны полосатого тела возбуждают проекционные нейроны; активность этих вставочных нейронов тормозит допамин.
У здорового человека черная субстанция находится в активном состоянии, постоянно стимулируя работу прямого пути. Стимуляция прямого пути необходима для активации ДМО перед движением и во время него. Возбуждение ДМО, которое возникает сразу перед началом двигательного акта, можно зарегистрировать при помощи электродов, прикрепленных к волосистой части головы. Эту электрическую активность называют (электрическим) потенциалом готовности (происхождение этого потенциала описано и показано на рисунке ниже). Импульсы от ДМО направляются к двигательной коре, откуда мозжечково-та-ламо-корковые волокна избирательно стимулируют пирамидные и корково-сетчатые нейроны за миллисекунды до генерации потенциала действия.
Скорлупа и бледный шар организованы соматотопически. За счет этого возможно селективно стимулировать нейроны, отвечающие, например, за движения рукой (прямой путь), и при этом одновременно тормозить ненужные движения, например ногой (непрямой путь). Для подавления ненужных в данный момент двигательных актов особенно важна работа СТЯ, которое может воздействовать на определенный сегмент ВБШ. Известно, что разрушение СТЯ приводит к тому, что движения конечностей противоположной половины тела становятся размашистыми и неконтролируемыми (см. далее).
Постепенное снижение выработки дофамина в компактном веществе черной субстанции — один из предрасполагающих факторов болезни Паркинсона (БП).
в) Лимбическая петля. На рисунке ниже изображена лимбическая петля базальных ганглиев. Она идет от нижней префронтальной коры через прилежащее ядро (передний конец полосатого тела) и вентральный отдел бледного шара, а затем возвращается в нижнюю префронтальную кору через дорсальное медиальное ядро (ДМЯ) таламуса. Прилежащее ядро и расположенный рядом обонятельный бугорок называют вентральным полосатым телом (вентральный стриатум).
Вероятнее всего, благодаря лимбической петле мы можем выражать свои эмоции в различных движениях, например улыбкой, жестами, принятием агрессивной или покорной позы. У этой петли имеется множество дофаминергических окончаний, которые разрушаются при БП. Из-за этого у таких больных лицо становится маскообразным, исчезает произвольная жестикуляция, через несколько лет развивается деменция.
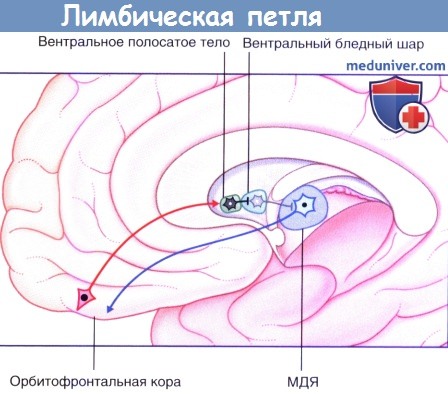
Лимбическая петля базальных ганглиев, правое полушарие.
Активация медиального дорсального ядра (МДЯ) таламуса происходит за счет его дезингибиции.
г) Глазодвигательная петля. Глазодвигательная петля начинается в лобном глазодвигательном поле (поле 8) и задней теменной коре (поле 7). Она проходит через хвостатое ядро и сетчатую часть черной субстанции. Далее через ПВЯ таламуса она возвращается в лобное глазодвигательное поле и префронтальную кору. От СЧЧС отходят тормозящие ГАМК-ергические волокна к верхним холмикам, где они образуют синапсы с нейронами, контролирующими непроизвольные саккады (Глава 23). Эти нейроны непосредственно иннервирует лобное глазодвигательное поле.
При фиксации взора СЧЧС находится в активном состоянии. Когда человек планирует перевести взгляд на какой-либо объект, происходят активация глазодвигательной петли и дезингибиция верхних холмиков (устранение торможения). При отсутствии нормального ингибирования со стороны базальных ганглиев происходит возбуждение верхних холмиков, в результате чего повышается активность прямого пути. Глазные яблоки с максимально высокой скоростью (22 м/с) перемещаются в сторону цели, после чего СЧЧС продолжает выполнять свою контролирующую функцию.
Для выявления глазодвигательной гипокинезии при болезни Паркинсона (БП) существует ряд специальных тестов. Движения глаз в сторону объекта, находящегося на периферическом поле зрения, обычно замедленны и иногда недостаточны. Объяснением этого симптома может служить нарушение дезингибиции верхних холмиков вследствие разрушения нейронов СЧЧС.
д) Резюме. Базальные ганглии представляют собой группу ядер, отвечающих за контроль над двигательной активностью. К ним относят полосатое тело (в том числе прилежащее ядро), бледный шар, СТЯ, черную субстанцию, а также вентральное латеральное (ВЛЯ), ПВЯ и ДМЯ таламуса. Бледный шар имеет наружную (НБШ) и внутреннюю (ВБШ) части, последняя продолжается в средний мозг в виде СЧЧС. Компактная часть черной субстанции лежит вне основных нейронных связей, но влияет на них за счет нигростриарного пути.
Кора больших полушарий возбуждает нейроны полосатого тела и СТЯ. Полосатое тело, в свою очередь, тормозит нейроны бледного шара. Влияние бледного шара на СТЯ и таламус также тормозящее. СТЯ возбуждает нейроны ВБШ.
Прямой путь, полосатое тело => ВБШ, находится в активированном состоянии за счет нормальной тонической активности нигростриарных дофаминергических нейронов. Непрямой путь, полосатое тело => НБШ => СТЯ => ВБШ, ингибирован. Для активации ДМО перед началом движения и во время него в двигательной петле происходит усиление передачи нервных импульсов по прямому пути. Активность ДМО, которую наблюдают непосредственно перед началом двигательного акта, можно зарегистрировать в виде потенциала готовности. Его появление обусловлено торможением нейронов ВБШ с последующим высвобождением (дезингибицией) таламо-корковых нейронов, посылающих нервные импульсы в ДМО. Затем сигнал поступает в двигательную кору, и начинается двигательный акт.
Полосатое тело и бледный шар организованы соматотопически, за счет чего становится возможным избирательный контроль той или иной части тела. Для подавления нежелательных движений особенно важно СТЯ.
Основной функцией двигательной петли, по всей видимости, служит организация правильного порядка выполнения отдельных движений. Это необходимо для реализации выученных двигательных программ. У пациентов с БП разрушение нигростриарных дофаминергических нейронов приводит к тому, что начинает преобладать активность непрямого пути. За этим следует подавление активности ВЛЯ таламуса и ДМО, что клинически проявляется брадикинезией. К основным симптомам БП относят ригидность, тремор и расстройство постуральных рефлексов.
Когнитивная петля начинается в ассоциативной коре, далее через ПВЯ таламуса она возвращается в премоторную и префронтальную кору. Активация когнитивной петли происходит во время изучения новых двигательных программ. Эта петля также отвечает за планирование двигательной активности.
Лимбическая петля начинается в поясной коре и миндалевидном теле, проходит через прилежащее ядро и направляется к ДМО. Вероятно, она отвечает за физическое выражение эмоций.
Глазодвигательная петля дезингибирует СЧЧС и таким образом активирует верхние холмики, которые инициируют саккады.
Гиперкинезы наблюдают при хорее Хантингтона, гемибаллизме и церебральном параличе (ЦП).
Редактор: Искандер Милевски. Дата публикации: 23.11.2018
Читайте также:
