
Нервная цепочка червя от которой отходят нервы
Дождевой червь – это типичный представитель типа Кольчатые черви.
Особенности строения систем органов дождевых червей
Данный тип живых организмов обладает самой высокой степенью организации по сравнению с другими типами червей.
Это проявляется в следующем:
- у них впервые появляется вторичная полость тела, кровеносная система и высокого организованная нервная система;
- полость тела выстлана эластичными стенками из мезодермы.
Такую полость тела целесообразно сравнивать с подушками безопасности, расположенными в каждом сегменте тела. Поэтому каждый сегмент тела получает полноценную опору и поддержку.
Кольчатые черви обитают:
- в пресной воде;
- в морской воде.
Они не чувствительны к солености почв, но при этом обладают высокой степенью чувствительности к их увлажненности.
Дождевой червь обладает круглым в поперечном сечении телом, насчитывающим более ста сегментов. Тело красновато – коричневой окраски, осветленное на брюшной стороне. У дождевых червей развиваются настоящие ткани. Снаружи тело покрыто эктодермой, состоящий из эпителия, который богат железами. Также у кольчатого червя имеется прогрессивно развитая мускульная система. Она состоит из кольцевых, продольных мышц. Дождевой червь может существенно изменяться в размерах, поскольку он достаточно хорошо сжимается, благодаря сокращению мышц.
- Курсовая работа Как устроена нервная система дождевого червя 400 руб.
- Реферат Как устроена нервная система дождевого червя 230 руб.
- Контрольная работа Как устроена нервная система дождевого червя 230 руб.
Пищеварительная система начинается на переднем конце тела ротовым отверстием, затем пища поступает в глотку, пищевод, весьма расширенный зоб, мускульный желудок. Далее тянется кишка, которая способствует ферментативному перевариванию пищи и всасыванию питательных веществ обратно в организм. Задняя часть кишки весьма коротка и все переваренное выбрасывается через анальное отверстие. Его появление можно считать эволюционным ароморфозом.
Ароморфоз – это прогрессивное эволюционное приспособление организма, которое отличается расширением адаптационных возможностей организма.
Кровеносная система дождевого червя замкнута и состоит из кровеносных сосудов. Главных сосудов всего два: брюшной и грудной. В каждом сегменте присутствует кольцевой сосуд, который пульсирует ритмически, что способствует перегонке крови из спинного сосуда в брюшной. В плазме крови присутствует гемоглобин, благодаря чему она окрашивается в красный цвет.
Задай вопрос специалистам и получи
ответ уже через 15 минут!
Дыхание кольчатых червей называется кожным. Газообмен осуществляется через всю поверхность тела. Черви очень чувствительны к влажной почве. На сухих песчаных почвах они быстро теряют жизнеспособность. Поэтому определённую степень увлажненности почв можно признать лимитирующим фактором для существования многих кольчатых червей.
Органы выделения имеют вид петлеобразных изогнутых трубочек, которые одним концом открываются в полость тела, а другим наружу. Такая система называется воронкообразным метанефридием. Метанефридии способствуют выделению продуктов жизнедеятельности из тела организма кольчатого червя через его поверхность.
Размножение и развитие кольчатых червей происходит только половым путем. Эти организмы являются гермафродитами. Органы половой системы располагаются в сегментах передней части тела дождевого червя. Семенники, как правило, расположены впереди яичников. При спаривании сперматозоиды каждого из двух червей переносятся в семяприёмники (особые полости) другого. Оплодотворение червей перекрёстное.
Копуляция или спаривание сопровождается выделением слизи и образованием кокона (муфты) для откладки яиц. Внутри этой муфты расположено большое количество белка для питания последующего зародыша. Червь выползает задним концом вперед и откладывает яйца. Края муфты достаточно быстро слипаются и полученный кокон остается в земляной норке. Эмбрионально яйца развиваются внутри кокона и из него в дальнейшем выходят молодые особи червей. У данных живых организмов отсутствует стадия личинки, то есть их развитие можно назвать прямым. В одном коконе развивается достаточно много зародышей, выходящих из водянистой жидкости. яйца дождевого червя весьма богаты желтком, что позволяет ему развивать вполне полноценный организм. По спиральному типу происходит дробление яйца.
Нервная система дождевого червя
Но, функционирование каждой указанной системы становится возможным благодаря тому, что кольчатый червь обладает весьма прогрессивной нервной системой. Передний членик дождевого червя содержит в себе окологлоточное нервное кольцо или крупное скопление нервных клеток. От него отходит брюшная нервная цепочка и она обладает ганглиями в каждом сегменте.
Нервная система дождевого червя – это система ганглиев, отходящих от окологлоточного нервного кольца, которые соединены в цепочку на каждом сегменте тела
Эта нервная система узлового типа образовалась благодаря слиянию нервных тяжей на правой и левой стороне тела. Она обеспечивает самостоятельность члеников и слаженную работу всех органов. По сути, брюшная нервная цепочка, состоит из сближенных по отношению к друг другу продольных нервных тяжей и парными нервными узлами или своеобразными ганглиями. Только у примитивных форм отсутствует такая организация нервной системы. Каждый сегмент тела контролируется собственным нервном узлом, что позволяет мышечному волокну сохраняться весьма эффективно и обеспечивать движение червя в плотной среде. Нервная система кольчатых червей отражает тенденцию к сближению нервных клеток по отношению к друг другу и централизации функций нервных клеток.
Наряду с полноценным развитием нервной системы у кольчатых червей достаточно слабо развиваются органы чувств. Черви не обладают развитыми органами зрения. Но в их кожном покрове есть светочувствительные клетки. Там же помещаются рецепторы осязания, вкуса, обоняния. Дождевые черви способны к регенерации (легко восстанавливает заднюю часть).
Таким образом, нервная системы дождевых червей развита весьма прогрессивно, она в полной мере обладает развитым для данного типа организма устройством и может выполнять функцию координации и регуляции жизнедеятельности. Благодаря наличию брюшной нервной цепочке кольчатые черви обладают возможностью к быстрому реагированию на изменения окружающей среды.
Так и не нашли ответ
на свой вопрос?
Просто напиши с чем тебе
нужна помощь
Вдоль средней линии брюшной стороны тела у дождевых червей под мускулатурой тянется нервный ствол, называемый брюшной нервной цепочкой.
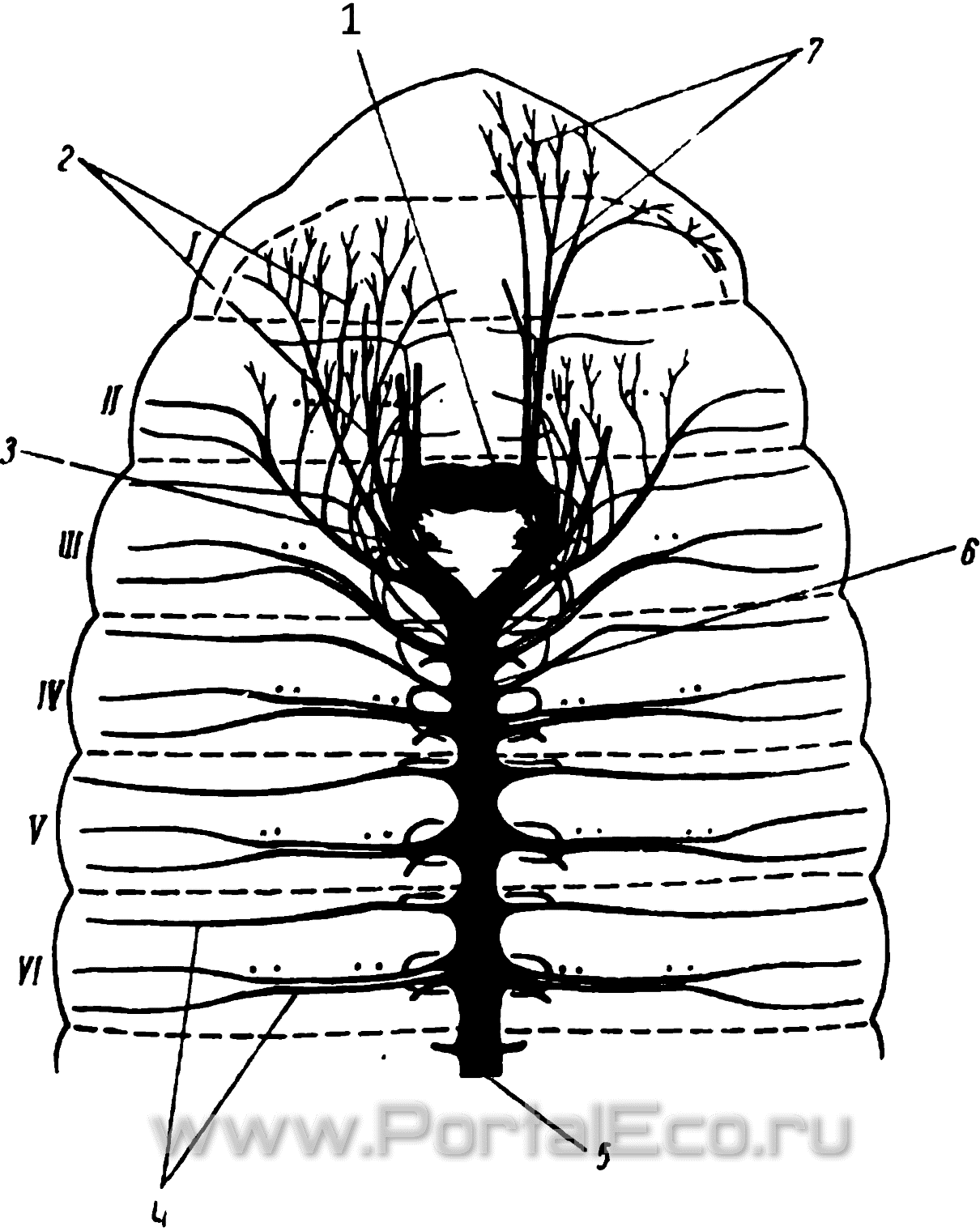
Рис. 17. Нервная система нередних сегментов дождевого червя. Вид со спинной стороны. (Из Штольте).
2 — надглоточный ганглий; 2 — нервы от окологлоточной комиссуры; з — окологлоточные комиссуры; 4 — сегментальные нервы; 6 — брюшная нервная цепочка; 6 — подглоточный ганглий; 7 — нервы головной лопасти; I—VI — 1-й—6-й сегменты.
В каждом сегменте тела имеется нервный узел, представляющий собой скопление нервных клеток и отдающий от себя 3 пары нервов. Ганглии соединены друг с другом перемычками, коннективами, которые, кроме нервных волокон, также содержат нервные клетки. В переднем конце тела, в 3-м сегменте, брюшная нервная цепочка разделяется на правую и левую глоточные коннективы, образующие окологлоточное нервное кольцо, соединяющееся с надглоточным, или головным ганглием (рис. 17). Этот ганглий — парный, состоит из правой и левой половин, плотно соединенных друг с другом.
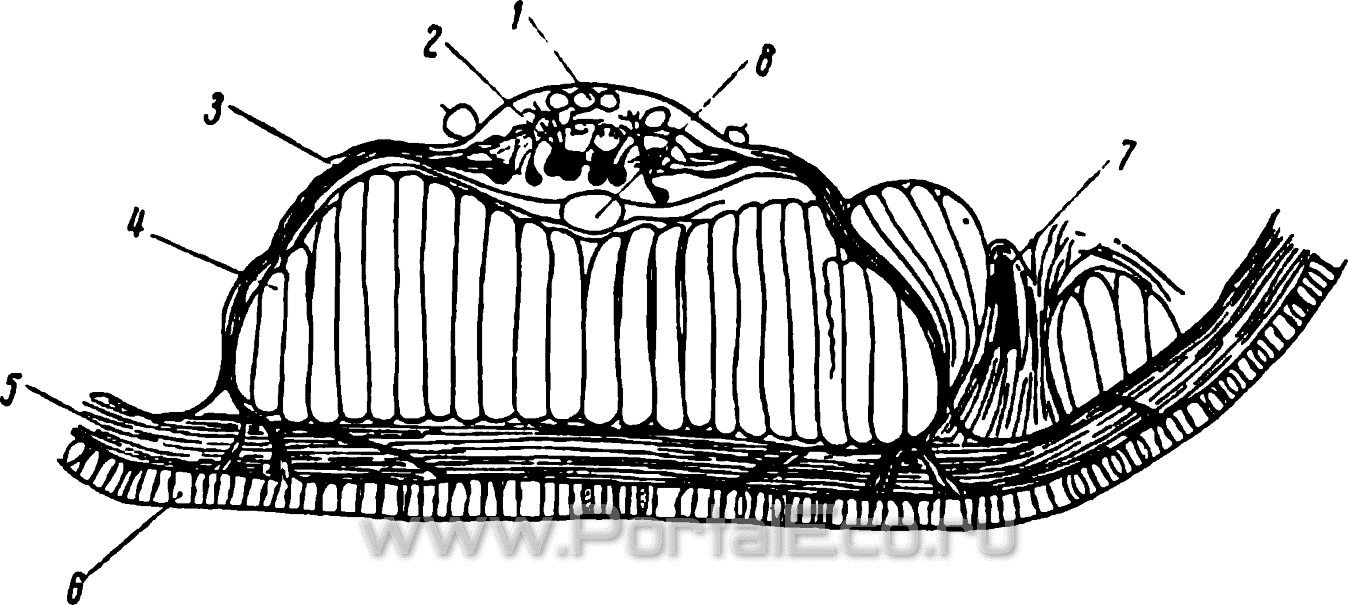
Рис. 18. Брюшная часть понеречного среза через дождевого червя.
1 — опорные волокна внутри брюшной нервной цепочки; 2 — нервные клетки в ганглии брюшной нервной цепочки; 3 — нерв; 4 — продольная мускулатура; 5 — кольцевая мускулатура; б — кожный эпителий с чувствительными нервными клетками; 7 — щетинка; 8 — брюшной кровеносный сосуд.
Но в (наличие от всех остальных нервных узлов, лежащих на брюшной стороне тела подкитечником, этот ганглий находится на спинной стороне тела и лежит над кишечником. Ганглий этот можно противопоставить всем прочим и вследствие того, что морфологически он сравним с головным мозгом вышестоящих форм (членистоногих). От него идут вперед многочисленные нервные стволы, обильно разветвляющиеся и образующие густые нервные сплетения в первых трех сегментах. Под глоткой, в месте расхождения глоточных коннектив, лежит подглоточный ганглий, представляющий собой результат слияния нескольких ганглиев брюшной нервной цепочки.
Как видно на поперечных разрезах, нервные клетки лежат по периферии ганглия, а средняя его часть занята сплетением отростков нервных клеток (рис. 18). В брюшной нервной цепочке обращают на себя внимание три очень толстых волокна, пробегающих но всей длине тела червя под соединительнотканной капсулой нервной цепочки на ее спинной стороне. Это так называемые нейро-хорды, которые еще совсем недавно принимали за гигантские нервные волокна. Однако теперь окончательно выяснено, что они являются своеобразными опорными образованиями. Эти образования и по строению, и по функции, и по положению между нервной системой и кишечником сходны с хордой позвоночных.
Нервы, отходящие от ганглиев брюшной цепочки, содержат двигательные волокна, заканчивающиеся в мускулатуре, и чувствительные, по которым раздражения поступают в нервную систему с периферии. Тела чувствительных нервных клеток находятся на периферии, в том числе в наружном эпителии (рис. 18). Нервные клетки здесь стоят в ряду эпителиальных клеток. Этот чрезвычайно древний тип соотношений элементов нервной системы сохранился у дождевых червей от нх далеких предков, примитивных многоклеточных животных. Очень интересно, что чувствительными нервными клетками здесь становятся не только клетки наружного эпителия, но, как это недавно установлено, также и клетки кишечника, происходящие из внутреннего зародышевого листка.
Чувствительные нервные клетки и их окончания имеются и в других местах тела
Ими богато снабжены также нефридии, щетинковые мешки и другие органы. Таким образом, у дождевых червей, как и у высших животных, работа внутренних органов протекает под контролем регулирующей и централизующей роли нервной системы.
Из рефлексов дождевых червей лучше всего известны наблюдаемые в акте ползания. При движении червя по всей длине тела, от переднего конца до заднего, пробегают перистальтические волны комбинированных сокращении мускулатуры. Они следуют друг за другом, и каждая следующая волна может возникнуть еще задолго до того, как первая дойдет до заднего конца тела. Казалось бы очевидным, по аналогии с высшими животными, что при- чиной этих волн сокращений служит последовательная передача раздражений вдоль по брюшной нервной цепочке. Однако, к удивлению исследователей, выяснилось, что
перерезание брюшного нервного ствола и даже вырезание из нее нескольких нервных узлов не останавливают бегущих волн сокращений мускулатуры: волна сокращения проходит через место повреждения так же, как это было и у нормального червя. С тем же результатом можно вдобавок к разрыву брюшной нервной цепочки удалить мускулатуру нескольких сегментов или повредить ее кислотой.
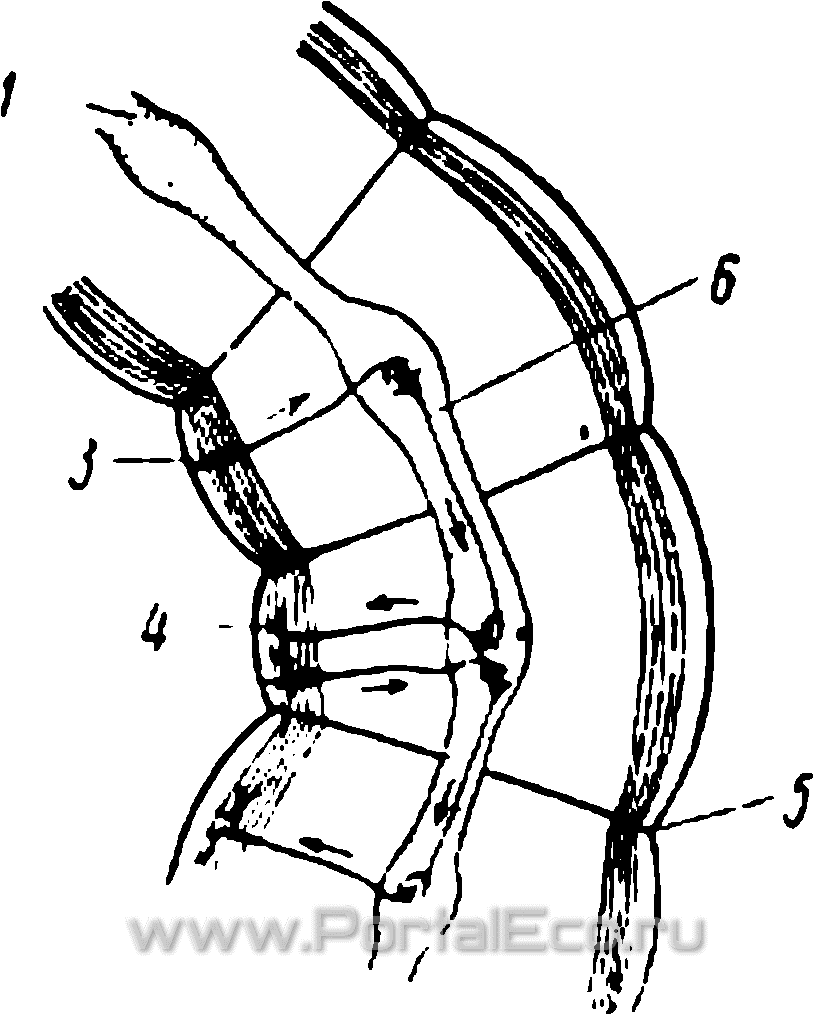
Рис. 19. Схема бисег-люптального рефлекса у дождевого червя.
/ — брюшная нервная цепочка; 2 — мускулатура стенки тела; з — чувствительная нервная клетка, воспринимающая импульс; 4 — оконч ние двигательной нервной клетки в мышцах; б — диссепимент; 6 — нервная клетка, передающая раздражение в соседний сегмент.
Анализ этих и аналогичных опытов показал, что поступательное движение червя представляет собой длинную цепь рефлекторных актов, в которой каждый сегмент является в значительной мере автономной физиологической единицей. Раздражения с периферии приводят к сокращению мускулатуры данного сегмента. Вследствие этого сокращения раздражаются периферические аппараты в соседнем сегменте, которые вызывают и в нем сокращения и т. д. Таким образом, комбинированные сокращения мышц в каждом сегменте могут представлять собой самостоятельный рефлекс, начинающийся с возбуждения чувствительных клеток на периферии и кончающийся эффектом сокращения мускулатуры этого сегмента. Это наиболее примитивный тип реакции на внешние воздействия. Некоторым его усложнением является передача полученного раздражения по нервной цепочке в соседний задний сегмент тела, в ответ на которое мускулатура этого сегмента сокращается. На рис. 19 изображена схема рефлекса при дугообразном сгибании червя, когда волна сокращений мускулатуры идет по одной стороне тела. Этот рефлекс и является основным при осуществлении поступательного движения червя вперед. Этот способ передачи раздражений по телу, как указано выше, говорит о слабой централизации его нервной системы.
Несколько большее значение имеет подглоточный ганглий, так как после его удаления червь лишается многих присущих ему способностей: очень страдают его вкусовые способности.
Было бы ошибочно думать, что слабая централизация нервной системы и относительная автономия отдельных сегментов, обнаруживаемые при поступательном движении червя, означают отсутствие реакций организма как целого. Заранее можно сказать, что таких реакций не может не быть, и, действительно, их очень легко обнаружить. При слабом раздражении заднего конца тела (при легком прикосновении) червь ползет вперед, при раздражении спереди он быстро весь сокращается и ползет в другую сторону, при сильном раздражении в любом месте червь начинает сильно сокращаться дугообразно, в разных направлениях (так называемые гимнастические движения); черви обнаруживают быстрые реакции на свет, на запахи и т. д. Таким образом, указанные выше несовершенство нервной системы и ее слабая централизация обнаруживаются только при внимательном наблюдении и в специально поставленных экспериментах.
Итак, нам известно, что червь располагает довольно богатым арсеналом возможностей для осуществления тех или иных реакций на изменении, происходящие и окружающей их среде.
Рассмотрим теперь, каким путем он может распознавать эти нзменепия. Средствами для этого являются органы чувств.
Как уже сказано, вся поверхность тела червя покрыта огромным количеством чувствительных первных клеток. Эти клетки служат органами осязания, развитого у червей очень сильно. Известно, что достаточно, осторожно приблизившись, слабо дунуть на червя, чтобы он ответил резким сокращением продольной мускулатуры; с помощью такого движепия он прячется в норку. Вдобавок к чувствительным нервным клеткам в наружном эпителии между клетками имеется очень большое количество свободных нервных окончаний, которые, скорее всего, также обеспечивают функцию осязания. Кроме изолированных светочувствительных клеток в наружном эпителии, имеются большие группы светочувствительных клеток, расположенные под кожей по ходу нервов, особенно в головной лопасти.

Рис. 20. Светочувствительная клетка из кожного эпителия дождевого червя. (Из Стефенсона).
1 — ядро; 2 — линза; 3 - нерв; 4 — сеть нейрофибрилл.

Рис. 22. Группа светочувствительных клеток, расположенных по ходу нерва, при большем увеличении. (Из Стефенсона).
1 — ядро; 2 — нейрофибрнллы; 3 — нерв; 4 — линза.
Дарвин тщательно исследовал восприятие света дождевыми червями. Он нашел, что если осторожно освещать их бесшумным фопарем, имеющим только узкий пучок света, интенсивность которого уменьшена красным или синим стеклом (цвет стекла безразличен), то только
Как известно уже более ста лет назад, дождевые черви, несмотря на отсутствие глаз, хорошо воспринимают свет. Восприятие света производится особыми светочувствительными клетками, которые по большей части располагаются поодиночке между клетками наружного эпителия (рис. 20). Внутри этих клеток, кроме ядра и густой сети тончайших волоконец — нейрофибрилл, имеется прозрачное светопреломляющее тельце бобовидной или удлиненной формы; его называют линзой по аналогии с хрусталиком глаза более высоко организованных животных. От тела клетки отходит нервный отросток, вступающий в подкожное нервное сплетение и связывающий ее с центральной нервной системой. Такая клетка несомненно представляет собой простейший глаз, как бы изолированную и автономную клетку сетчатки глаза высших животных. Светочувствительные клетки сосредоточены главным образом в передних сегментах тела; больше всего их в головной лопасти, где их может быть свыше 50 (рис. 21). В последующих сегментах число их быстро убывает, в середине тела их пе находят, а в последних трех сегментах их снова становится больше. У некоторых видов дождевых червей, очень немногие черви реагируют, а именно: уходят в свои норы.[1]
Способность ощущать свет играет очень большую роль в жизни червей, так как солнечный свет на них действует губительно (черви очень чувствительны к ультрафиолетовой части солнечного спектра). Реакция ухода в темноту спасает им жизнь.
В научной литературе издавна от времени до времени появлялись сообщения о звуках, издаваемых дождевыми червями. Действительно, при трении тела и щетинок о грунт, при поршневых движениях во влажных норках, при растирании пищи в глотке, при затаскивании листьев и камешков и т. д. могут возникать звуки. Они тем лучше ощутимы, чем червей больше и чем они крупнее. Но очень сомнительно, чтобы эти звуки имели какое-нибудь биологическое значение.
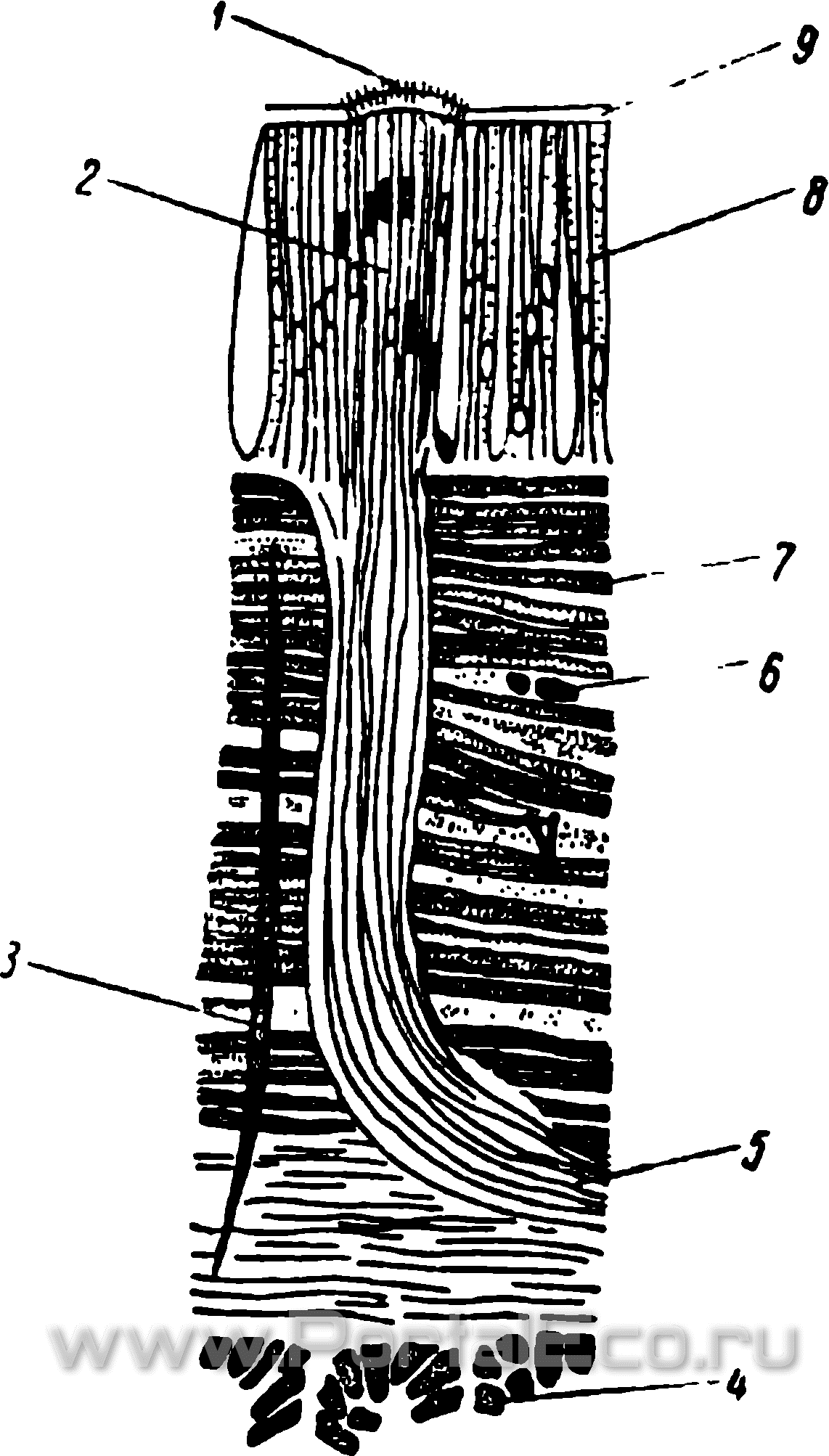
Рис. 23. Поперечный разрез через дождевого червя. Чувствительная почка в кожном эпителии. (По Гессе).
1 - чувствительные волоски; 2 - чувствительные клетки; 3 — спинно-брюшное мышечное волокно; 4 — продольные мышцы; 5 — чувствительный нерв; 6 — капилляр; 7 — кольцевые мышцы; 8 — эпителиальные клетки; 9 — кутикула.
Кроме чувствительных нервных клеток, нервных окончаний и светочувствительных клеток в наружном эпителии рассеяно большое количество органов, представленных комплексами клеток. Их называют иногда чувствительными почками. Несколько десятков чувствительных клеток образуют цилиндрический или яйцевидный комплекс (рис. 23). Это — чувствительные нервные клетки и длинные нервные отростки, которые идут в брюшную нервную цепочку. Поверхность кутикулы в области чувствительной точки несколько приподнята, а каждая клетка снабжена чувствительным волоском. Эти микроскопические органы распространены в большом количестве по всему телу, но особенно многочисленны они в 1-м сегменте и в его головной лопасти, где у крупных видов их насчитывается около 1800. Функция их точно не установлена. Как полагают некоторые исследователи, часть их может нести осязательную функцию. Но вряд ли можно сомневаться в том, что ими же осуществляются и функции обоняния и вкуса. В пользу такого заключения свидетельствует тот факт, что эти органы имеются в большом количестве в ротовой полости.
Чувство обоняния, т. е. способность к распознаванию различных веществ в газообразном состоянии (что и является способностью воспринимать запахи), развито у червей сравнительно, слабо. В опытах Дарвина черви не реагировали на запах табачного сока, духов, уксусной кислоты, но куски лука (который они очень любят) и листья капусты они находили по запаху. На эфир, поднесенный на близком расстоянии к переднему концу тела, черви реагировали отрицательно и немедленно устранялись от него.
Чувство вкуса, т. е. способность распознавать химические различия веществ при контакте с ними, развито у червей очень тонко и наряду с чувством осязания служит для них главным источником восприятия событий внешнего мира. Опытами Дарвина, развитыми в последнее время рядом исследователей, способность червей выбирать себе пищу доказана совершенно бесспорно и возражения, высказанные некоторыми авторами по этому поводу (например, Тарнани, 1928), несомненно основаны на ошибках.
На утро подсчитывают, сколько испытателей черви затащили в норки и при этом отмечают, за какой из концов испытателя ухватился червь. Нужно сказать, что черви, собирая корм, попадающийся им на поверхности земли, никогда его глубоко не вносили в норки, а оставляли его недалеко от наружного отверстия или же только пододвигали к нему. Поэтому вышеуказанный подсчет сделать нетрудно. Если червь не различает концов испытателя вкуса, то при достаточно большом числе повторений опыта должно оказаться, что как за тот, так и за другой его конец червь берется одинаково часто. Если он предпочитает испытуемое вещество чистому желатину, то пропитанный им конец должен оказываться чаще впереди при затаскивании. Наоборот, если вещество оказывается на вкус хуже, чем чистый желатин, то за него червь должен ухватываться реже. Этот опыт видоизменяется подкидыванием червям испытателей вкуса, пропитанных разными веществами, с последующим определением числа втянутых в норки тех и других. Результаты обрабатывались статистически.
Чистый желатин их привлекает более, чем высушенные листья. Гниющие листья разных растений могут быть выстроены в такой ряд в порядке убывающей склонности к ним червей: ива, сладкий лупин, грецкий орех, белая акация, тополь, дуб, горький лупин, липа, бук, клеи, конский каштан. Свежие листья располагаются совсем в другой последовательный ряд. От желатина с примесью хинина черви отказываются, причем чувствуют это вещество уже в концентрации 0.07%. От минеральных кислот они отказываются в любых концентрациях, но прибавка к желатину 1—2% лимонной и фосфорной кислот им нравится. К сахарам они относятся безразлично, но от очень крепких растворов
Сахара отказываются вовсе. К сахарину обнаруживается отрицательная реакция, уже начиная с ничтожных концентраций.
Способность определять форму тел у червей, по-видимому, отсутствует. Предпочтительное затаскивание ими в норки листьев за передний конец, а сосновой хвои за основание (факт, установленный Дарвином), дальнейшими исследованиями подтвержден. Однако опытами Мангольда установлено, что черви при этом руководствуются только чувством вкуса, позволяющим им отличать верхушку листа от черешка.
Как уже отмечено, аппарат, обеспечивающий наличие условных рефлексов, может быть локализован и в частях нервной системы, не соответствующих головному мозгу вышестоящих форм. Определить, где именно локализована эта функция у дождевых червей, — дело будущих исследований. Чтобы закончить рассмотрение рефлекторных реакций у дождевых червей, коснемся еще вопроса о болевых ощущениях у них.
[1] Однако имеются наблюдения, что черни более чувствительны к синему свету, чем к красному.
Существует несколько типов организации нервной системы, представленные у различных систематических групп животных.
Нервная система различных животных.
Царство животных разделяют на два подцарства : одноклеточные и многоклеточные, каждое из которых включает в себя по несколько типов.
Круглые черви (лат. Nemathelminthes) имеют нервную систему ортогонального типа. Нематоды составляют основной класс, в который входит большинство видов типа круглые черви. Нервная система у них состоит из центрального и периферического отделов. К центральному относится нервное кольцо, окружающее глотку, и отходящие от него нервные стволы. Периферический отдел представляет собой отходящие от центров нервные ветви и сплетения отростков нервных клеток. От окологлоточного кольца вперед отходят шесть коротких веточек, а назад шесть длинных, которые связаны между собой кольцевыми нервами. Наиболее хорошо развиты два ствола, проходящие в спинном и брюшном валиках гиподермы, первый иннервирует обе спинные мышечные ленты, а второй – обе брюшные. Для нематод характерно постоянное количество клеток в нервной системе.
Схема нервной системы аскариды с брюшной стороны (по Брауну):
1 - ротовые сосочки с осязательными окончаниями и иннервирующими их нервами,
2 - окологлоточное нервное кольцо,
3 - боковые головные ганглии,
4 - брюшной нервный ствол,
5 - боковые нервные стволы,
6 - кольцевые нервы,
7 - задний ганглий,
8 - чувствительные сосочки с соответствующими нервами,
9 - анальное отверстие,
10 - спинной нервный ствол
У членистоногих (лат. Arthropoda) нервная система организована по типу брюшной нервной цепочки, то есть как у кольчатых червей. При этом усиливается роль надглоточных ганглиев, которые сообща образуют головной мозг, состоящий из трех отделов: переднего – протоцеребрума, среднего – дейтоцеребрума и заднего – тритоцеребрума. Отмечается тенденция к олигомеризации ганглиев брюшной нервной цепочки, что выражается в уменьшении количества узлов за счет их слияния. Обычно очень хорошо развиты многочисленные органы чувств, обеспечивающие животному восприятие основных внешних раздражителей.
2 - нейросекреторные клетки,
3 - оптическая область мозга,
5 - антеннальный нерв,
7 - кардиальные тела,
8 - прилежащие тела,
9 - окологлоточные коннективы,
10 - подглоточный ганглий
, 11 - нервы, идущие к ротовым конечностям,
12 - ганглии грудных сегментов,
13 - ганглии брюшных сегментов,
14 - непарный нерв симпатической системы
Нервная система паукообразных отличается разнообразием строения. Общий план ее организации соответствует брюшной нервной цепочки, однако имеется ряд особенностей. В головном мозге отсутствует дейтоцеребрум, что связано с редукцией придатков акрона – антеннул, которые иннервируются этим отделом мозга у ракообразных, многоножек и насекомых. Сохраняются передний и задний отделы головного мозга. Ганглии брюшной нервной цепочки часто концентрируются, образуя более или менее выраженную ганглиозную массу. У сенокосцев и клещей все ганглии сливаются, образуя кольцо вокруг пищевода, однако у скорпионов сохраняется выраженная брюшная цепочка ганглиев.
Существует несколько типов организации нервной системы, представленные у различных систематических групп животных.
Нервная система различных животных.
Царство животных разделяют на два подцарства : одноклеточные и многоклеточные, каждое из которых включает в себя по несколько типов.
Круглые черви (лат. Nemathelminthes) имеют нервную систему ортогонального типа. Нематоды составляют основной класс, в который входит большинство видов типа круглые черви. Нервная система у них состоит из центрального и периферического отделов. К центральному относится нервное кольцо, окружающее глотку, и отходящие от него нервные стволы. Периферический отдел представляет собой отходящие от центров нервные ветви и сплетения отростков нервных клеток. От окологлоточного кольца вперед отходят шесть коротких веточек, а назад шесть длинных, которые связаны между собой кольцевыми нервами. Наиболее хорошо развиты два ствола, проходящие в спинном и брюшном валиках гиподермы, первый иннервирует обе спинные мышечные ленты, а второй – обе брюшные. Для нематод характерно постоянное количество клеток в нервной системе.
Схема нервной системы аскариды с брюшной стороны (по Брауну):
1 - ротовые сосочки с осязательными окончаниями и иннервирующими их нервами,
2 - окологлоточное нервное кольцо,
3 - боковые головные ганглии,
4 - брюшной нервный ствол,
5 - боковые нервные стволы,
6 - кольцевые нервы,
7 - задний ганглий,
8 - чувствительные сосочки с соответствующими нервами,
9 - анальное отверстие,
10 - спинной нервный ствол
У членистоногих (лат. Arthropoda) нервная система организована по типу брюшной нервной цепочки, то есть как у кольчатых червей. При этом усиливается роль надглоточных ганглиев, которые сообща образуют головной мозг, состоящий из трех отделов: переднего – протоцеребрума, среднего – дейтоцеребрума и заднего – тритоцеребрума. Отмечается тенденция к олигомеризации ганглиев брюшной нервной цепочки, что выражается в уменьшении количества узлов за счет их слияния. Обычно очень хорошо развиты многочисленные органы чувств, обеспечивающие животному восприятие основных внешних раздражителей.
2 - нейросекреторные клетки,
3 - оптическая область мозга,
5 - антеннальный нерв,
7 - кардиальные тела,
8 - прилежащие тела,
9 - окологлоточные коннективы,
10 - подглоточный ганглий
, 11 - нервы, идущие к ротовым конечностям,
12 - ганглии грудных сегментов,
13 - ганглии брюшных сегментов,
14 - непарный нерв симпатической системы
Нервная система паукообразных отличается разнообразием строения. Общий план ее организации соответствует брюшной нервной цепочки, однако имеется ряд особенностей. В головном мозге отсутствует дейтоцеребрум, что связано с редукцией придатков акрона – антеннул, которые иннервируются этим отделом мозга у ракообразных, многоножек и насекомых. Сохраняются передний и задний отделы головного мозга. Ганглии брюшной нервной цепочки часто концентрируются, образуя более или менее выраженную ганглиозную массу. У сенокосцев и клещей все ганглии сливаются, образуя кольцо вокруг пищевода, однако у скорпионов сохраняется выраженная брюшная цепочка ганглиев.
Сохранились животные, которые не имеют нервной системы. Это – простейшие одноклеточные организмы и примитивные многоклеточные. У одноклеточных животных функции восприятия выполняют особые участки мембраны клетки (хемочувствительные, фоточувствительные), а моторные функции – специальные органы движения (мембранеллы) типа ресничек, жгутиков и т. п.
У таких сложно организованных одноклеточных организмов, как инфузории, обнаружены внутриклеточные органеллы, выполняющие функции анализа поступающей информации (сенсориум) и организации согласованных движений эффекторов (моториум), а также специальные фибриллы (кинотодесмы), по которым передаются команды от моториума к эффекторам. Т.е. у одноклеточных организмов отсутствует необходимость в существовании специализированных клеток, поскольку сам организм является одноклеточным.
У многоклеточных организмов произошла дифференциация клеток тела по их расположению в теле (наружные, промежуточные и внутренние, выстилающие внутреннюю полость), так и по функции.
Часть поверхностных клеток специализировалась на восприятии внешних воздействий и превратилась в чувствительные (рецепторные). Другие клетки приобрели способность сокращаться и, объединившись, создали мышечные слои. И, наконец, появились клетки, специализированные на передаче возбуждения от рецепторов к мышечным клеткам. Это – нервные клетки. Поскольку мышечные клетки образовали мышечные слои, способные выполнять согласованные сокращения, нервные клетки тоже должны были объединиться в систему для их координации. Так развивается нервная система. Чем более совершенны движения животных, тем лучше развита их нервная система. Неподвижные животные (губки) имеют на поверхности вокруг пор отдельные эпителиально-мышечные клетки, которые способны и воспринимать раздражения, и реагировать на них сокращением.
Тем не менее, диффузная сеть с окологлоточным нервным кольцом была относительно медленно действующим устройством. Измеренная проводимость по нервной сети кишечнополостных составляет не более 5-20 см/с. Этого явно не хватает животным размером более 5 см, поэтому уже у актиний выделились участки нервной сети с высокой скоростью проведения (см. рис. 3, в). В некоторых случаях она достигает 150 см/с, что делает актиний изощрёнными охотниками за значительно более эволюционно продвинутыми позвоночными. Окологлоточное нервное кольцо было явным достижением, но оно не могло дифференциально управлять всем телом или обеспечить контроль за свободным плаванием.
У свободно живущих многоклеточных (кишечнополостные) существует сеть нервных клеток, сгущающаяся в тех местах организма, которые выполняют функцию перемещения тела или захвата и заглатывания пищи. Таким образом, у наиболее примитивного представителя кишечнополостных (пресноводной губки) сеть короткоотростчатых нервных клеток сгущается вблизи подошвы, вокруг щупалец и ротового отверстия. У более сложно организованного кишечнополостного (медузы) сеть уплотняется в тяжи, образующие два кольца вокруг колокола (зонтика). Согласованные сокращения мышц колокола медузы позволяют ей перемещаться в воде по принципу реактивного двигателя.
В нервной сети кишечнополостных нейроны соединяются между собой отростками как протоплазматическим (непрерывным), так и эфаптическим (непосредственный контакт между отростками) способами. Обнаружены и настоящие синаптические контакты. Такая организация нервной системы называется сетевидной, или диффузной.
Так, уиглокожих в центре вокруг пищевода нервные клетки, концентрируясь, образуют три нервных кольца, от которых в каждый луч (у морской звезды) отходят соответственно по три нервных ствола. В стволах имеются и тела нервных клеток, связанных с периферической нервной сетью. Справедливо было бы выделить диффузно-узловой тип организации нервной системы, промежуточный между примитивным диффузным и сложным централизованным. Его развитие связано прежде всего с совершенствованием анализаторной функции.
Уже умедуз можно обнаружить скопления нервных клеток в виде узлов (краевые тельца), выполняющих функцию анализа информации, получаемой от органов чувств (глазков, органа равновесия). В связи с концентрацией органов чувств на переднем конце тела у животных с билатеральным строением тела там же скапливаются нервные клетки в виде узла – "головного мозга". Например, уплоских червей от головного узла к переднему концу тела отходят головные нервы к органам чувств (глазам, органам осязания и т. д.), а к заднему – несколько продольных нервных тяжей ("стволов"), соединяющихся между собой поперечными перемычками. Тяжи состоят из длинных отростков нервных клеток, а также из самих нервных клеток с короткими отростками. Подобную конструкцию называют ортагоном. Такое же строение нервной системы и у круглых червей, но у них тяжи начинаются от окологлоточного кольца.
Централизованная узловая (ганглионарная) нервная система дождевого червя представлена окологлоточным нервным кольцом и брюшной нервной цепочкой. В окологлоточном кольце сильно развитые надглоточные ганглии соединены перемычками с менее развитыми подглоточными узлами; которые, в свою очередь, связаны с нервной цепочкой Цепочка состоит из парных ганглиев, располагающихся в каждом сегменте тела червя Головные надглоточные ганглии переплетаются, в основном, с дистантными рецепторами. Однако дальнейшее развитие церебральных (головных) надглоточных ганглиев зависит от развития интегративной (ассоциативной) функции.
У моллюсков имеется несколько пар ганглиев в "стратегически важных" частях тела:
ü церебральные ганглии, связанные преимущественно с рецепторами головы, расположены в головной части тела;
ü педальные (моторные) ганглии – у начала ноги;
ü висцеральные ганглии – вблизи внутренних органов;
ü плевральные ганглии – между головой и ногой. Ганглии соединены между собой нервными стволами, а с периферией – нервами. У двустворчатых моллюсков нервная система менее развита и состоит лишь из трех пар узлов.
Ø у головоногих моллюсков (кальмары, осьминоги, каракатицы) ЦНС является наиболее сложной:
ü головнойганглий у них стал единственным, а все остальные слились с ним, образовав мощную массу нервной ткани (головной мозг). Головной мозг защищен хрящевой капсулой, имеет множество (до 14) долей. Последние осуществляют связь с хорошо развитыми органами чувств и выполняют анализаторную (сенсорную) функцию:
· подглоточные доли, а также базальные доли надглоточного отдела замыкаются на различных мышцах и определяют моторную функцию,
· задним долям мозга свойственны интегративные (ассоциативные) функции.
Функции головного мозга головоногих моллюсков и позвоночных животных совпадают. Задние доли по своим функциям и свойствам сходны с ассоциативной корой млекопитающих.
ü нижнего (окологлоточного) – тритоцеребрума,
ü среднего – дейтероцеребрума;
У ракообразных ЦНС имеет головные (надглоточные и подглоточные) ганглии и цепочку их в головогруди. У некоторых раков цепочка ганглиев сливается в один крупный узел, у паукообразных вместе с надглоточным ганглием она образует в головогруди сложноорганизованный мозг с большим количеством нервных клеток.
У насекомых нервная система кроме головного мозга имеет цепочку парных ганглиев, которые объединяются в крупные узлы:
ü брюшные узлы выполняют преимущественно висцеральные функции,
ü грудные – моторные функции (организация движения ног и крыльев).
Сенсорные же (анализаторные) и интегративные (ассоциативные) функции контролируются у насекомых головным мозгом.
Читайте также:
