
Нервная цепочка на брюшной стороне тела
Хордовые — это тип животных, отличающийся разнообразием представителей, достигших эволюционного прогресса. В наше время хордовых насчитывают около 60 тысяч видов. К ним относятся рыбы, лягушки, ящерицы, птицы, звери и др. Представители хордовых обитают в воде, на суше, в воздухе и почве. В ходе эволюции они приспособились к самым разнообразным условиям окружающей среды.
Несмотря на свое разнообразие все хордовые обладают общим планом строения, который напоминает перевернутый в спино-брюшном направлении общий план строения большинства беспозвоночных. У хордовых нервная трубка находится над кишечником (и хордой или позвоночником), кровь течет на брюшной стороне тела от хвоста к голове, на спинной — от головы к хвосту. А у большинства беспозвоночных имеется брюшной нервная цепочка, кровь течет в обратном по-сравнению с хордовыми направлении (на спине — от хвоста к голове, на брюхе — от головы к хвосту).
Главной отличительной особенностью хордовых является наличие внутреннего осевого скелета. У наиболее примитивных представителей (ланцетник, некоторые группы рыб) роль осевого скелета выполняет хорда, которая выглядит как плотный упругий, но достаточно эластичный продольный тяж (стержень). Он состоит из хрящеподобной ткани. У большинства более высоко организованных представителей хордовых в процессе эмбрионального развития на месте хорды развивается позвоночный столб (позвоночник). Он может быть хрящевым или костным. Хорда образуется из продольного выроста кишечной трубки со спинной стороны, т. е. имеет энтодермальное происхождение.
Именно у типа Хордовые нервная система достигла наивысшего развития. Характерна трубчатая нервная система. Нервная трубка закладывается над хордой и имеет эктодермальное происхождение. У большинства передний отдел нервной трубки расширяется и образует головной мозг. При этом невроцель (полость нервной трубки) преобразуется в желудочки мозга.
Пищеварительная трубка находится под хордой, а под пищеварительной трубкой находится сердце (или аналогичный ему сосуд).
Жабры образуются не на поверхности тела, а внутри него — в глотке. Таким образом глотка оказывается пронизанной жаберными отверстиями. Они сохраняются у всех первичноводных хордовых, у остальных — жабры есть только у эмбрионов в начальных стадиях их развития.
Кровеносная система хордовых замкнутая.
Хордовых относят к группе вторичноротых животных, так как в процессе эмбрионального развития рот у них образуется не со стороны впячивания бластулы, как у большинства беспозвоночных, которых в связи с этим называют первичноротыми, а с противоположной стороны. На месте впячивания бластулы у хордовых развивается анальное отверстие.
Все хордовые относятся к вторичнополостным животным.
Тип Хордовые включает три подтипа. Это головохордовые, или бесчерепные (ланцетники), личиночнохордовые (оболочники) и позвоночные, или черепные (все остальные). У оболочников хорда имеется только на стадии личинки. Наиболее многочисленный по видовому составу и распространенности подтип Позвоночные.
26. Происхождение нервной системы хордовых
Наиболее часто обсуждаемые гипотезы происхождения не могут объяснить появление одного из основных признаков хордовых — трубчатой нервной системы, которая располагается на спинной стороне тела. Мне хотелось бы использовать другой подход. На основании анализа морфологической организации нервной системы можно попытаться воссоздать последовательность этапов её филогенетического становления.
Стоит обратить внимание на строение современных личиночно-хордовых (Olson, Holmberg, Lilliemarc, 1990; Meglitsch, Schram, 1991).
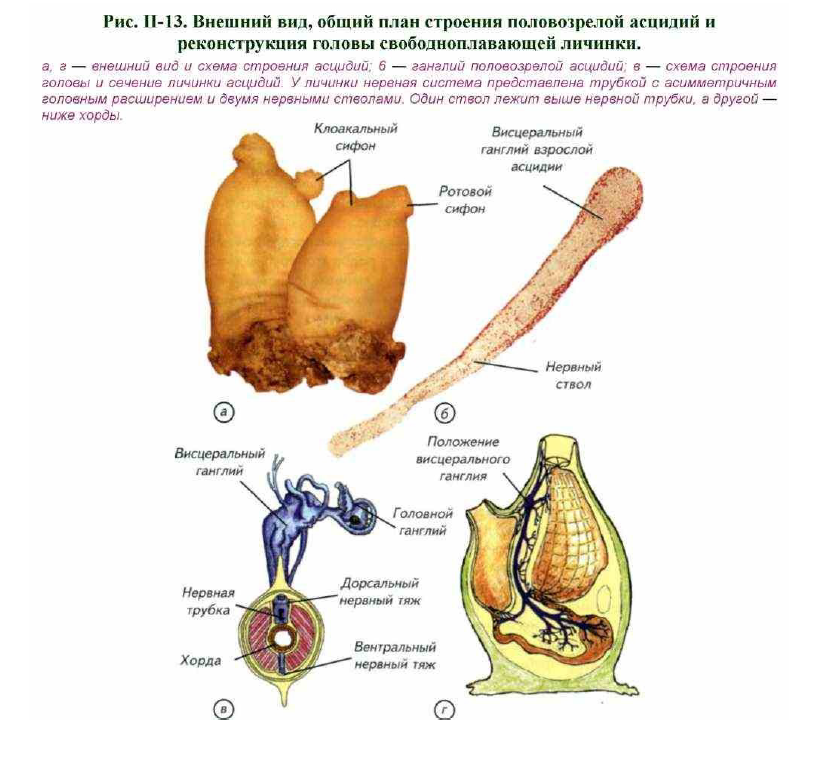
Это большая группа морских животных, имеющих на стадии личинки выраженную спинную струну — хорду, над которой расположена нервная трубка (рис. II-13). Их личинки, как правило, свободноплавающие и всегда
снабжены хвостом. Наиболее характерным представителем этой группы является асцидия. Свободноплавающие личинки асцидий по общему виду немного напоминают головастиков лягушек с овальным телом и длинным, сжатым с боков хвостом. По оси хвоста идёт хорда, представляющая собой
столбик студенистого вещества, одетый одним слоем клеток. Над хордой расположена нервная трубка, а по бокам — мышцы.
В теле личинки нервная трубка утолщается, а в головной зоне приобретает вид пузырька (см. рис. II-13). Это чувствительное образование является предшественником мозга и содержит рецепторы. Личинка
асцидий может воспринимать свет с помощью светочувствительного глазка и узнавать о направлении гравитационного поля благодаря статоцисту. Активно перемещающееся животное имеет глотку, сердце,
околосердечную полость и выделительную систему, открывающуюся наружу одиночным отверстием. В таком состоянии большинство личинок находится только несколько часов. Затем они прикрепляются к какому-
либо предмету при помощи специальных сосочков и превращаются во взрослое оседлое животное.
Такое превращение носит название регрессивного метаморфоза. Он заключается в увеличении числа глоточных дыхательных отверстий, полном исчезновении хвоста и хорды. Наибольшим изменениям
подвергается нервная система. Исчезают хвостовой отдел нервной трубки, глазок и статоцист (Torrence, Cloney, 1983). Вместо головного пузырька и туловищной нервной трубки формируется одиночный ганглий,
свойственный нервной системе беспозвоночных (см. рис. II-13). Он лежит между ротовым и анальным отверстиями, вытянут в спинно-брюшном направлении и формирует несколько нервных стволов. На брюшной
стороне нервного узла расположена субнейральная железа, которую гомологизируют с гипофизом позвоночных. Таким образом, из активной свободноплавающей личинки с хорошо развитыми органами чувств,
хордой и сложноорганизованной нервной системой возникает сидячее взрослое животное. У прикреплённой асцидии органы, указывающие на родство с позвоночными, изменяются до неузнаваемости. Большую часть
тела оседлой формы занимает пищеварительный тракт U-образной формы с обширной глоточной полостью.
Описанный жизненный цикл не является универсальным для всех оболочников. У некоторых примитивных родов этой группы хвостатой стадии развития нет, а у других развитие личинки происходит в
родительском организме. Этому способствует гермафродитизм (двуполость) взрослых оболочников.
Следующими по уровню организации являются бесчерепные, или головохордовые, которые обладают всеми основными признаками хордовых (Hirakow, Kajita, 1990). Они имеют во взрослом состоянии
хорошо развитые хорду, спинной мозг и жаберные щели. Единственным представителем группы является ланцетник (Amphioxus lanceolatus) — небольшое животное с прозрачным телом, которое заострено с обоих
концов и сжато с боков. Нервная система ланцетника расположена над хордой, между мышцами правой и левой стороны тела (см. рис. I-12, б-г).
При этом мышечные волокна не иннервируются отростками нервных клеток ланцетника. Сами мышечные клетки образуют отростки, которые входят в нервную трубку. По ним поступает управляющий сигнал в
мышечные волокна. Аналогичное строение имеют нервно-мышечные связи у круглых червей. Их мышечные клетки также образуют отростки, соединяющие мышцы с нервными стволами, проходящими вдоль тела (см.
рис. I-12, д).
Центральная нервная система имеет вид трубки, которая в поперечном разрезе представляет собой треугольник с полым центральным каналом.
Сама трубка состоит преимущественно из продольных нервных волокон и тел клеток, сгруппированных вокруг центрального канала. Периферическая нервная система состоит из нервов, отходящих от центральной нервной
системы. Они делятся на две группы: первая образована несимметричнымипарами нервов, отходящих от передней части нервной трубки, называемой мозгом; вторая группа состоит из большого числа спинных нервов, отходящих от спинного мозга (рис. II-14).
Первая пара головных нервов отходит от переднего края нервной трубки, а вторая — от его спинной стороны. Обе пары нервов имеют нервные узлы, которые размещены в головной части ланцетника. Спинные
нервы распределены посегментно и соответственно расположению миомеров нервы правой и левой стороны отходят поочередно, а не друг против друга. В каждом сегменте с каждой стороны тела есть два нерва:
спинной и брюшной. Спинной нерв отходит одним корешком от верхней стороны спинного мозга. Нервы идут к коже и к мышцам, являясь как чувствительными, так и двигательными. Брюшной нерв состоит из
многочисленных отдельных волокон, которые входят в нервную трубку от миомеров (см. рис. II-12, г; II-14).
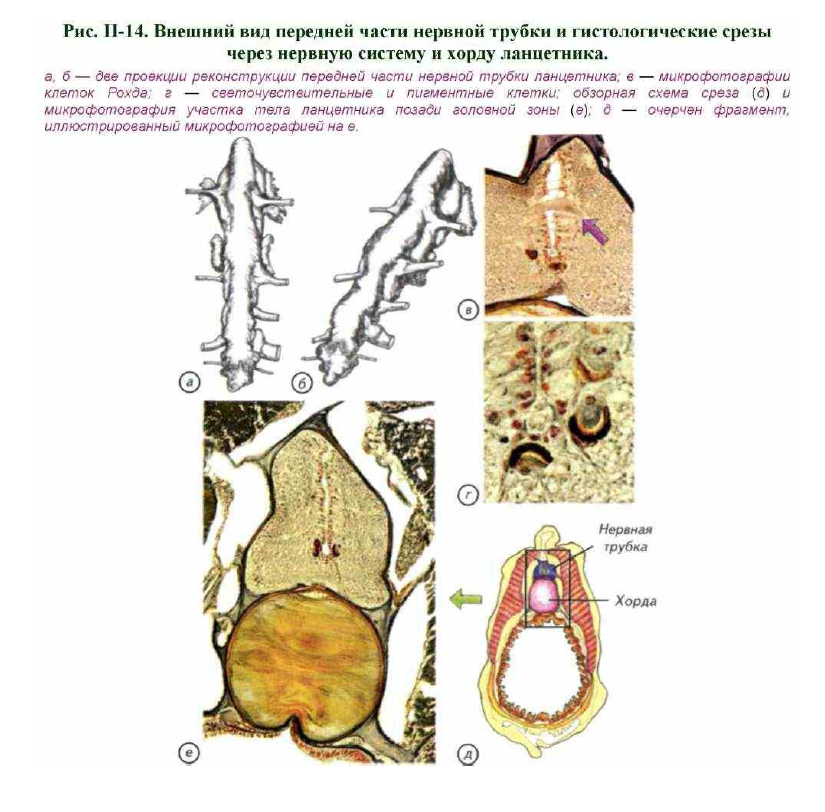
В передней части нервной трубки центральный канал незначительно расширяется, образуя большую полость желудочек передней части нервной трубки, а несколько позади него формируется спинное расширение. Это
образование, как и вся нервная трубка, покрыто тонкой соединительнотканной оболочкой. Передний верхний отдел нервной трубки выдаётся небольшим полым мешком (асимметричная ямка Кёлликера), который соединяется с органом обоняния и носит название средней обонятельной доли. На его заднем конце с брюшной стороны есть небольшое углубление, называемое воронкой. Органы чувств ланцетника представлены асимметричной обонятельной ямкой, открывающейся наружу на левой стороне головы. Этот хеморецепторный орган имеет полость, которая открывается через нейропор в центральный канал
нервной трубки. Органы зрения ланцетника весьма своеобразны. В головной части они представлены непарным глазом, который выглядит как и пигментное пятно. Небольшие светочувствительные клетки расположены в толще нервной трубки на всём её протяжении. Каждая из них частично окружена пигментной клеткой. Такая пара клеток представляет собой отдельный простой глазок, способный улавливать различия освещённости.
Органы осязания представлены ротовыми щупальцами, а орган вкуса — небольшой ямкой Гатчека, которая расположена на верхней стороне предротовой воронки (Jefferies, 1986).
Строение нервной системы ланцетника имеет ряд особенностей, которые проливают свет на происхождение трубчатой нервной системы хордовых. Она является нервным стволом с узкой центральной щелью
внутри. Эта щель часто пересекается аксонами и дендритами нейронов, которые расположены вокруг спинномозгового канала. В головной части мозга ланцетника спинномозговой канал расширяется и становится
мозговым желудочком. В самой ростральной части нервной трубки находится непарное пигментное пятно. Каудальнее этого пятна расположена асимметричная обонятельная долька. Долька обнаружена
только слева и соединена нервными волокнами с одиночной обонятельной ямкой. Строение нервов ланцетника асимметрично, что нехарактерно для хордовых. Известны две пары симметричных нервов, отходящих от
головной части нервной трубки. Они не имеют двигательных ветвей, что позволяет считать их настоящими головными нервами. Эти нервы парные, но размеры левых нервов более чем в 2 раза превышают правые. Непарный обонятельный нерв состоит из нескольких разрозненных волокон. Он начинается от обонятельной дольки и оканчивается в одиночном обонятельном мешочке.
Кроме упомянутых нервов, выявлены ещё два нерва. Один нерв является парным и расположен на уровне обонятельной дольки (см. рис. II 14, а, б). Второй нерв непарный и выходит из левой половины мозга между
первой и второй парами симметричных нервов. Таким образом, у ланцетника имеется 5 головных нервов, которые нуждаются в простейшей классификации. Обонятельный асимметричный нерв логично называть I
головным нервом, первую симметричную пару — II головным нервом, симметричную пару нервов, лежащую каудальнее, — III головным нервом, асимметричный левосторонний нерв — IV головным нервом, а последнюю
головную пару нервов — V головным нервом. Вентрально расположенные симметричные двигательные корешки нервов, лежащие каудальнее V пары головных нервов, уже следует относить к дериватам спинного мозга. Надо отметить, что эта классификация не имеет отношения к гомологизации
нервов ланцетника с позвоночными.
Внутреннее строение нервной системы также даёт достаточно фактов, свидетельствующих о сближении морфологических особенностей ланцетника с возможными предками хордовых. Это асимметричный
глазок, который лежит в переднем крае головного желудочка, асимметричная обонятельная долька и гигантские нейроны, которые находятся в центре желудочка (см. рис. II-14, в). Гигантские нейроны
имеют уникальное для хордовых морфологическое строение. Их отростки распространяются вдоль нервной трубки и объединяют нервные центры управления сегментированной мускулатурой. Благодаря интегративным
функциям гигантских нейронов ланцетник может совершать волнообразные движения. Нейроны с аналогичным расположением и морфологией отростков нехарактерны для позвоночных. Они больше
напоминают организацию нервной системы беспозвоночных.
В процессе развития ланцетника закладка органов асимметрична. Рот ланцетника закладывается на левой стороне тела, что совпадает с одним из вариантов предлагаемой гипотезы происхождения хордовых. Надо
подчеркнуть, что эмбриональная асимметрия ланцетника затронула и жаберный аппарат, который закладывается на правой стороне тела. На этой стороне тела формируются как правые, так и левые жаберные щели.
Ротовое отверстие закладывается на брюшной стороне. Только при формировании взрослого ланцетника жаберные щели перемещаются через брюшную поверхность и становятся парными. Ротовое отверстие в свою
очередь передвигается слева в центр, и симметрия восстанавливается.
Перечисленные особенности морфологической организации личиночно- и головохордовых позволяют предложить двухвариантную гипотезу возникновения нервной системы хордовых. Ключевым препятствием
любой гипотезы является вопрос перемещения нервной системы беспозвоночных из вентрального положения в дорсальное. На этот вопрос можно предложить два формально непротиворечивых ответа. Первым
вариантом можно считать предположение о перемещении центральной нервной системы на спинную сторону в результате поворота или переворота всего животного. Второй вариант предполагает, что нервная
система никогда не была на вентральной стороне тела и сформировалась совершенно иным образом.
Для обоих вариантов гипотез существуют общие проблемы. Является ли трубчатая нервная система гомологом брюшной нервной цепочки беспозвоночных? Если да, то как возникла полость внутри нервной трубки
и откуда произошли вегетативные нервные ганглии, расположенные около внутренних органов? Нужно ответить на вопрос о происхождении черепно- мозговых и спинномозговых нервов, а также и о причинах асимметрии нервной системы низших хордовых. Если предположить, что гомологии между нервной системой беспозвоночных и дорсальной нервной трубкой хордовых нет, то правомерен вопрос об источнике происхождения трубчатой нервной системы. Предположение о новообразовании трубчатой
нервной системы хордовых крайне маловероятно. По-видимому, преемственность между позвоночными и беспозвоночными следует искать в строении червеобразных и в морфологических особенностях
современных хордовых и позвоночных животных (Савельев, 2001).
Допуская, что кишечнополостные являются безусловными предками основных групп животных, мы получим гипотетическое существо, изображённое на рис. II-15, а. Внутри животного проходит пищеварительный канал, который представляет собой гастроваскулярную полость кишечнополостных, открытую спереди ротовым отверстием. У этого организма ещё нет билатеральной симметрии, и оно могло представлять собой какой- либо гидроид, перешедший к свободно-подвижному образу жизни. Примерное представление об этом организме и
его нервной системе может дать рис. II-4. Такое существо могло свободно перемещаться в трёхмерной водной среде в результате сокращений щупалец, перистальтически или пассивно с током воды. Чтобы
поддерживать своё тело во взвешенном состоянии, необходимо обеспечить его плавучесть.
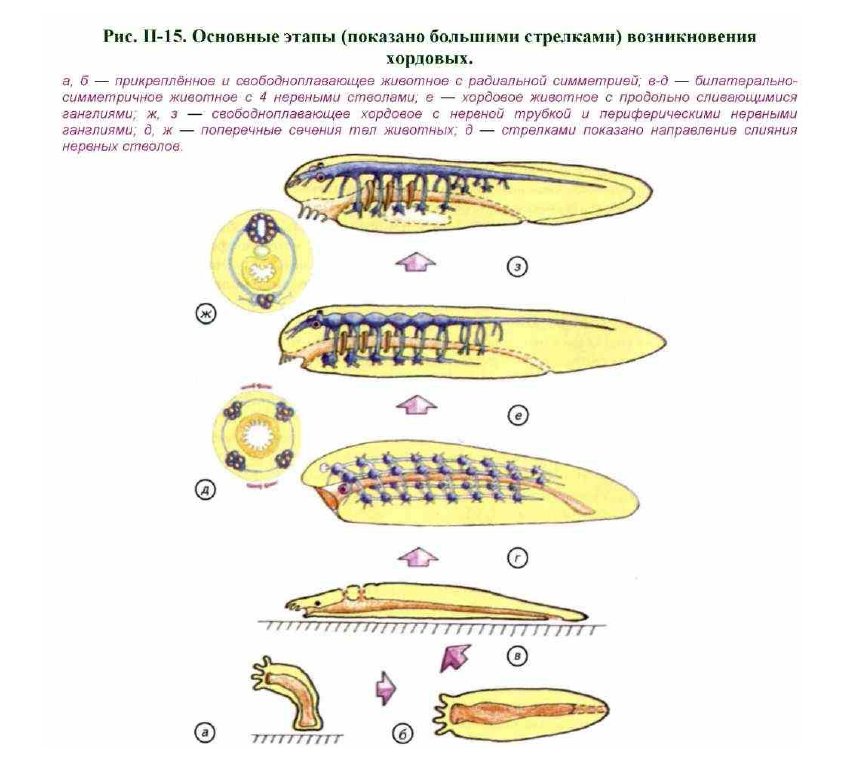
Увеличение объёма мезоглеи, в которой находятся нервные клетки, стало наиболее простым способом повышения плавучести кишечнополостных, поэтому расстояние между гастродермисом и
эпидермисом значительно увеличилось. По-видимому, спустя некоторое время основным достижением перехода к такому образу жизни явилось появление скоплений нервных клеток, которые сформировали
окологлоточное нервное кольцо и аборальный орган. Наиболее вероятной группой животных, похожих на описываемую модель, являются гребневики.
Животное с радиальной симметрией не нуждалось в усложнении организации, пока не перешло к придонному образу жизни. На первых этапах оно представляло собой кожно-мышечный мешок без отчётливой
оси симметрии. При передвижении по дну неизбежно должны были возникнуть спинная и брюшная поверхность эпидермиса, ротовое и анальное отверстия и билатеральная симметрия. Надо отметить, что
механизмы появления вторичноротости не имеет смысла рассматривать одновременно с эволюцией нервной системы. Это самостоятельная проблема, которая не имеет однозначных решений.
Можно предположить, что с переходом к придонному образу жизни с активным движением по субстрату у подобного предка хордовых могли появиться затруднения с дыханием. В воде придонной части океанов и
морей растворённого кислорода намного меньше, чем под поверхностью или в толще воды. Возникла потребность в усилении метаболизма. Если это произошло, то на спинной стороне кишечной трубки могли
сформироваться щели, которые играют роль жаберного аппарата (см. рис. II-15, в). Появление гомологов жабр у таких животных маловероятно, но полностью отрицать такую возможность нельзя, поскольку, не зная
размеров тела этого животного, мы не можем оценить его метаболизм.
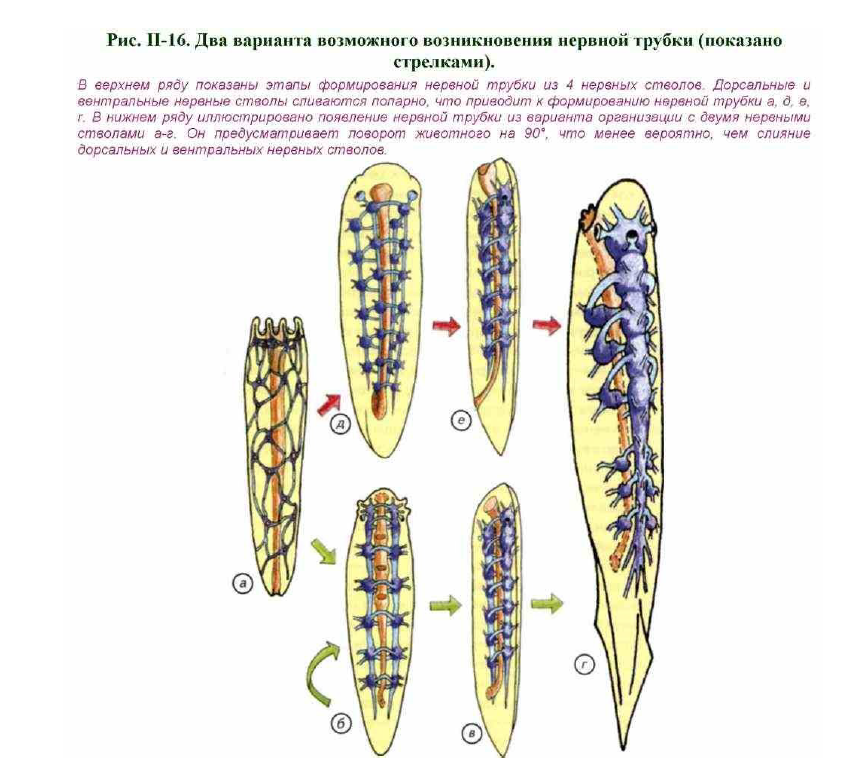
Замена радиальной симметрии на билатеральную с неизбежностью приводит к возникновению одной или нескольких симметричных пар нервных тяжей, идущих вдоль тела.
Исходя из элементарных принципов эффективной работы нервной системы, мы можем остановиться на двух вариантах организации: 2 или 4 нервных тяжа (рис. II-16, б, д). Два нервных ствола могут располагаться на
боковых краях уплощённого и вытянутого тела животного (см. рис. II-16, б). Краевое положение нервных тяжей обусловлено тем, что каждый тяж имеет рецепторы, расположенные на поверхности тела, а боковые
поверхности являются основными контактными зонами. Чем ближе к ним будет расположен нервный тяж, тем быстрее будет реакция организма на оказываемое воздействие. Такое строение нервной системы характерно для многих плоских червей, хотя описываемое животное является исключительно гипотетической моделью и не имеет отношения к рецентным формам. Два нервных головных ганглия переходят в широко
расставленные нервные тяжи, расположенные по бокам тела животного. В центре расположен пищеварительный канал, дорсальнее и вентральнее которого проходят поперечные комиссуры, соединяющие левый и правый нервные тяжи (см. рис. II-16, б).
Первичноводные позвоночные
Тип позвоночных (Vertebrata) включает в себя подтип бесчерепных (Acrania) и подтип черепных (Craniata). Единственными представителями бесчерепных являются многочисленные ланцетники (Amphioxi), которые
рассматривались в предыдущем разделе, посвящённом происхождению позвоночных. Настоящий раздел охватывает обширную и разнообразную группу первичноводных черепных позвоночных. Они сформировались и долгое время морфологически изменялись в водной среде. В процессе эволюции именно этой группы сложилась нервная система, характерная для всех остальных позвоночных. Впервые возникло трубчатое строение нервной системы. Полости желудочков головного и спинного мозга стали непрерывными, появилась циркуляция внутрижелудочковой мозговой жидкости. В сочетании с эффективной кровеносной системой и
внутренним скелетом это создало потенциальную возможность увеличивать размеры нервной системы первых водных позвоночных практически без ограничений. Эти эволюционные перспективы наращивания размеров мозга и параллельного увеличения сложности поведения не были реализованы в водной среде. Потребовались особые условия выхода на сушу, которые стали стимулом для развития новых качеств мозга и его ассоциативных функций (Northcutt, Davis, 1983).
Однако благодаря первичноводным позвоночным нервная система приобрела те универсальные черты, которые послужили морфологической основой для дальнейшей эволюции. Его конструкция оказалась настолько эффективной, что по существу не подверглась принципиальным изменениям за всю историю эволюции позвоночных (Nieuwenhuys, 1982).
Конечно, мозг адаптивно перестраивался во всех систематических группах, но принципы структурной организации центральной и периферической нервной системы сохранились без изменений. На конструкцию нервной системы радикально не повлияли появление ни челюстного аппарата, ни плавников, ни конечностей, ни выход на сушу, ни адаптация к полёту, нии теплокровность.
Нервная система, как и позвоночник, оказалась тем органом, который обеспечивал выживание и прогресс всей группы позвоночных.
Морфофункциональные отделы головного мозга, как и их основные функции, остаются неизменными на всём протяжении эволюции от миноги до человека. Эта универсальность строения нервной системы говорит об
общности происхождения всех позвоночных. Делать предположения о полифилитичности происхождения позвоночных довольно трудно.
Допущение параллельного возникновения и эволюции сотен связанных между собой структур мозга весьма сомнительно.
1. Найдите ошибки в приведенном тексте. Укажите номера предложений, в которых сделаны ошибки, исправьте их. (1) Одноклеточные животные обладают всем комплексом свойств целостного организма: дыханием, питанием, выделением, наследственностью, размножением, раздражимостью, подвижностью. (2) Многоклеточные животные, начиная с типа Плоские черви, относятся к группе трехслойных организмов. (3) Впервые в эволюции многоклеточных животных клеточная дифференциация появилась у плоских червей. (4) У плоских червей полость тела заполнена жидкостью. (5) У кольчатых червей кровеносная система замкнутого типа. (6) Аскариды – представители типа Круглые черви – не выводятся из кишечника человека с экскрементами, так как у них имеются присоски.
1) 3 - дифференцировка клеток впервые в эволюции появилась у представителей типа Кишечнополостные;
2) 4 – у плоских червей полость тела отсутствует, промежутки между органами и стенкой тела заполнены клетками паренхимой;
3) 6 – аскариды не имеют присосок, они постоянно движутся навстречу каловым массам.
2. Укажите не менее трех отличий в строении животных типа Круглые черви от свободноживущих животных типа Плоские черви.
1) У круглых червей первичная полость тела, заполненная жидкостью, у плоских червей полость тела отсутствует, промежутки между органами заполнены паренхимой.
2) У круглых червей только продольные мышцы (черви могут только изгибаться), у плоских червей – продольные, кольцевые, косые мышцы.
3) У круглых червей сквозная пищеварительная система, у плоских червей – слепозамкнутая пищеварительная система.
4) Круглые черви имеют спинной и брюшной нервные стволы, плоские черви – боковые нервные стволы.
3. При пересадке дождевых червей на поля, где их до этого было мало, удается повысить урожайность некоторых культур. Объясните данный факт.
1) дождевые черви рыхлят почву, сохраняя в ней влагу и способствуя проникновению кислорода, что необходимо для роста и развития растения;
2) дождевые черви обогащают почву перегноем, что повышает урожайность
4. Найдите ошибки в приведенном тексте. Укажите номера предложений, в которых сделаны ошибки, исправьте их. (1) Кольчатые черви – это наиболее высокоорганизованные животные среди других типов червей. (2) У кольчатых червей кровеносная система незамкнутого типа. (3) Тело кольчатого червя состоит из одинаковых члеников. (4) Полость тела у кольчатых червей отсутствует. (5) Нервная система кольчатых червей представлена окологлоточным кольцом и спинной нервной цепочкой. (6) Кольчатые черви обладают регенерацией. (7) Среди них различают многощетинковых, малощетинковых и пиявок.
2 – Кровеносная система замкнутого типа.
4 – У них впервые появилась вторичная полость тела – целом.
5 – Нервная цепочка расположена на брюшной стороне тела.

5. Рассмотрите предложенную схему. Запишите в ответе пропущенный термин, обозначенный на схеме знаком вопроса.
26. Происхождение нервной системы хордовых
Наиболее часто обсуждаемые гипотезы происхождения не могут объяснить появление одного из основных признаков хордовых — трубчатой нервной системы, которая располагается на спинной стороне тела. Мне хотелось бы использовать другой подход. На основании анализа морфологической организации нервной системы можно попытаться воссоздать последовательность этапов её филогенетического становления.
Стоит обратить внимание на строение современных личиночно-хордовых (Olson, Holmberg, Lilliemarc, 1990; Meglitsch, Schram, 1991).
Это большая группа морских животных, имеющих на стадии личинки выраженную спинную струну — хорду, над которой расположена нервная трубка (рис. II-13). Их личинки, как правило, свободноплавающие и всегда
снабжены хвостом. Наиболее характерным представителем этой группы является асцидия. Свободноплавающие личинки асцидий по общему виду немного напоминают головастиков лягушек с овальным телом и длинным, сжатым с боков хвостом. По оси хвоста идёт хорда, представляющая собой
столбик студенистого вещества, одетый одним слоем клеток. Над хордой расположена нервная трубка, а по бокам — мышцы.
В теле личинки нервная трубка утолщается, а в головной зоне приобретает вид пузырька (см. рис. II-13). Это чувствительное образование является предшественником мозга и содержит рецепторы. Личинка
асцидий может воспринимать свет с помощью светочувствительного глазка и узнавать о направлении гравитационного поля благодаря статоцисту. Активно перемещающееся животное имеет глотку, сердце,
околосердечную полость и выделительную систему, открывающуюся наружу одиночным отверстием. В таком состоянии большинство личинок находится только несколько часов. Затем они прикрепляются к какому-
либо предмету при помощи специальных сосочков и превращаются во взрослое оседлое животное.
Такое превращение носит название регрессивного метаморфоза. Он заключается в увеличении числа глоточных дыхательных отверстий, полном исчезновении хвоста и хорды. Наибольшим изменениям
подвергается нервная система. Исчезают хвостовой отдел нервной трубки, глазок и статоцист (Torrence, Cloney, 1983). Вместо головного пузырька и туловищной нервной трубки формируется одиночный ганглий,
свойственный нервной системе беспозвоночных (см. рис. II-13). Он лежит между ротовым и анальным отверстиями, вытянут в спинно-брюшном направлении и формирует несколько нервных стволов. На брюшной
стороне нервного узла расположена субнейральная железа, которую гомологизируют с гипофизом позвоночных. Таким образом, из активной свободноплавающей личинки с хорошо развитыми органами чувств,
хордой и сложноорганизованной нервной системой возникает сидячее взрослое животное. У прикреплённой асцидии органы, указывающие на родство с позвоночными, изменяются до неузнаваемости. Большую часть
тела оседлой формы занимает пищеварительный тракт U-образной формы с обширной глоточной полостью.
Описанный жизненный цикл не является универсальным для всех оболочников. У некоторых примитивных родов этой группы хвостатой стадии развития нет, а у других развитие личинки происходит в
родительском организме. Этому способствует гермафродитизм (двуполость) взрослых оболочников.
Следующими по уровню организации являются бесчерепные, или головохордовые, которые обладают всеми основными признаками хордовых (Hirakow, Kajita, 1990). Они имеют во взрослом состоянии
хорошо развитые хорду, спинной мозг и жаберные щели. Единственным представителем группы является ланцетник (Amphioxus lanceolatus) — небольшое животное с прозрачным телом, которое заострено с обоих
концов и сжато с боков. Нервная система ланцетника расположена над хордой, между мышцами правой и левой стороны тела (см. рис. I-12, б-г).
При этом мышечные волокна не иннервируются отростками нервных клеток ланцетника. Сами мышечные клетки образуют отростки, которые входят в нервную трубку. По ним поступает управляющий сигнал в
мышечные волокна. Аналогичное строение имеют нервно-мышечные связи у круглых червей. Их мышечные клетки также образуют отростки, соединяющие мышцы с нервными стволами, проходящими вдоль тела (см.
рис. I-12, д).
Центральная нервная система имеет вид трубки, которая в поперечном разрезе представляет собой треугольник с полым центральным каналом.
Сама трубка состоит преимущественно из продольных нервных волокон и тел клеток, сгруппированных вокруг центрального канала. Периферическая нервная система состоит из нервов, отходящих от центральной нервной
системы. Они делятся на две группы: первая образована несимметричнымипарами нервов, отходящих от передней части нервной трубки, называемой мозгом; вторая группа состоит из большого числа спинных нервов, отходящих от спинного мозга (рис. II-14).
Первая пара головных нервов отходит от переднего края нервной трубки, а вторая — от его спинной стороны. Обе пары нервов имеют нервные узлы, которые размещены в головной части ланцетника. Спинные
нервы распределены посегментно и соответственно расположению миомеров нервы правой и левой стороны отходят поочередно, а не друг против друга. В каждом сегменте с каждой стороны тела есть два нерва:
спинной и брюшной. Спинной нерв отходит одним корешком от верхней стороны спинного мозга. Нервы идут к коже и к мышцам, являясь как чувствительными, так и двигательными. Брюшной нерв состоит из
многочисленных отдельных волокон, которые входят в нервную трубку от миомеров (см. рис. II-12, г; II-14).
В передней части нервной трубки центральный канал незначительно расширяется, образуя большую полость желудочек передней части нервной трубки, а несколько позади него формируется спинное расширение. Это
образование, как и вся нервная трубка, покрыто тонкой соединительнотканной оболочкой. Передний верхний отдел нервной трубки выдаётся небольшим полым мешком (асимметричная ямка Кёлликера), который соединяется с органом обоняния и носит название средней обонятельной доли. На его заднем конце с брюшной стороны есть небольшое углубление, называемое воронкой. Органы чувств ланцетника представлены асимметричной обонятельной ямкой, открывающейся наружу на левой стороне головы. Этот хеморецепторный орган имеет полость, которая открывается через нейропор в центральный канал
нервной трубки. Органы зрения ланцетника весьма своеобразны. В головной части они представлены непарным глазом, который выглядит как и пигментное пятно. Небольшие светочувствительные клетки расположены в толще нервной трубки на всём её протяжении. Каждая из них частично окружена пигментной клеткой. Такая пара клеток представляет собой отдельный простой глазок, способный улавливать различия освещённости.
Органы осязания представлены ротовыми щупальцами, а орган вкуса — небольшой ямкой Гатчека, которая расположена на верхней стороне предротовой воронки (Jefferies, 1986).
Строение нервной системы ланцетника имеет ряд особенностей, которые проливают свет на происхождение трубчатой нервной системы хордовых. Она является нервным стволом с узкой центральной щелью
внутри. Эта щель часто пересекается аксонами и дендритами нейронов, которые расположены вокруг спинномозгового канала. В головной части мозга ланцетника спинномозговой канал расширяется и становится
мозговым желудочком. В самой ростральной части нервной трубки находится непарное пигментное пятно. Каудальнее этого пятна расположена асимметричная обонятельная долька. Долька обнаружена
только слева и соединена нервными волокнами с одиночной обонятельной ямкой. Строение нервов ланцетника асимметрично, что нехарактерно для хордовых. Известны две пары симметричных нервов, отходящих от
головной части нервной трубки. Они не имеют двигательных ветвей, что позволяет считать их настоящими головными нервами. Эти нервы парные, но размеры левых нервов более чем в 2 раза превышают правые. Непарный обонятельный нерв состоит из нескольких разрозненных волокон. Он начинается от обонятельной дольки и оканчивается в одиночном обонятельном мешочке.
Кроме упомянутых нервов, выявлены ещё два нерва. Один нерв является парным и расположен на уровне обонятельной дольки (см. рис. II 14, а, б). Второй нерв непарный и выходит из левой половины мозга между
первой и второй парами симметричных нервов. Таким образом, у ланцетника имеется 5 головных нервов, которые нуждаются в простейшей классификации. Обонятельный асимметричный нерв логично называть I
головным нервом, первую симметричную пару — II головным нервом, симметричную пару нервов, лежащую каудальнее, — III головным нервом, асимметричный левосторонний нерв — IV головным нервом, а последнюю
головную пару нервов — V головным нервом. Вентрально расположенные симметричные двигательные корешки нервов, лежащие каудальнее V пары головных нервов, уже следует относить к дериватам спинного мозга. Надо отметить, что эта классификация не имеет отношения к гомологизации
нервов ланцетника с позвоночными.
Внутреннее строение нервной системы также даёт достаточно фактов, свидетельствующих о сближении морфологических особенностей ланцетника с возможными предками хордовых. Это асимметричный
глазок, который лежит в переднем крае головного желудочка, асимметричная обонятельная долька и гигантские нейроны, которые находятся в центре желудочка (см. рис. II-14, в). Гигантские нейроны
имеют уникальное для хордовых морфологическое строение. Их отростки распространяются вдоль нервной трубки и объединяют нервные центры управления сегментированной мускулатурой. Благодаря интегративным
функциям гигантских нейронов ланцетник может совершать волнообразные движения. Нейроны с аналогичным расположением и морфологией отростков нехарактерны для позвоночных. Они больше
напоминают организацию нервной системы беспозвоночных.
В процессе развития ланцетника закладка органов асимметрична. Рот ланцетника закладывается на левой стороне тела, что совпадает с одним из вариантов предлагаемой гипотезы происхождения хордовых. Надо
подчеркнуть, что эмбриональная асимметрия ланцетника затронула и жаберный аппарат, который закладывается на правой стороне тела. На этой стороне тела формируются как правые, так и левые жаберные щели.
Ротовое отверстие закладывается на брюшной стороне. Только при формировании взрослого ланцетника жаберные щели перемещаются через брюшную поверхность и становятся парными. Ротовое отверстие в свою
очередь передвигается слева в центр, и симметрия восстанавливается.
Перечисленные особенности морфологической организации личиночно- и головохордовых позволяют предложить двухвариантную гипотезу возникновения нервной системы хордовых. Ключевым препятствием
любой гипотезы является вопрос перемещения нервной системы беспозвоночных из вентрального положения в дорсальное. На этот вопрос можно предложить два формально непротиворечивых ответа. Первым
вариантом можно считать предположение о перемещении центральной нервной системы на спинную сторону в результате поворота или переворота всего животного. Второй вариант предполагает, что нервная
система никогда не была на вентральной стороне тела и сформировалась совершенно иным образом.
Для обоих вариантов гипотез существуют общие проблемы. Является ли трубчатая нервная система гомологом брюшной нервной цепочки беспозвоночных? Если да, то как возникла полость внутри нервной трубки
и откуда произошли вегетативные нервные ганглии, расположенные около внутренних органов? Нужно ответить на вопрос о происхождении черепно- мозговых и спинномозговых нервов, а также и о причинах асимметрии нервной системы низших хордовых. Если предположить, что гомологии между нервной системой беспозвоночных и дорсальной нервной трубкой хордовых нет, то правомерен вопрос об источнике происхождения трубчатой нервной системы. Предположение о новообразовании трубчатой
нервной системы хордовых крайне маловероятно. По-видимому, преемственность между позвоночными и беспозвоночными следует искать в строении червеобразных и в морфологических особенностях
современных хордовых и позвоночных животных (Савельев, 2001).
Допуская, что кишечнополостные являются безусловными предками основных групп животных, мы получим гипотетическое существо, изображённое на рис. II-15, а. Внутри животного проходит пищеварительный канал, который представляет собой гастроваскулярную полость кишечнополостных, открытую спереди ротовым отверстием. У этого организма ещё нет билатеральной симметрии, и оно могло представлять собой какой- либо гидроид, перешедший к свободно-подвижному образу жизни. Примерное представление об этом организме и
его нервной системе может дать рис. II-4. Такое существо могло свободно перемещаться в трёхмерной водной среде в результате сокращений щупалец, перистальтически или пассивно с током воды. Чтобы
поддерживать своё тело во взвешенном состоянии, необходимо обеспечить его плавучесть.
Увеличение объёма мезоглеи, в которой находятся нервные клетки, стало наиболее простым способом повышения плавучести кишечнополостных, поэтому расстояние между гастродермисом и
эпидермисом значительно увеличилось. По-видимому, спустя некоторое время основным достижением перехода к такому образу жизни явилось появление скоплений нервных клеток, которые сформировали
окологлоточное нервное кольцо и аборальный орган. Наиболее вероятной группой животных, похожих на описываемую модель, являются гребневики.
Животное с радиальной симметрией не нуждалось в усложнении организации, пока не перешло к придонному образу жизни. На первых этапах оно представляло собой кожно-мышечный мешок без отчётливой
оси симметрии. При передвижении по дну неизбежно должны были возникнуть спинная и брюшная поверхность эпидермиса, ротовое и анальное отверстия и билатеральная симметрия. Надо отметить, что
механизмы появления вторичноротости не имеет смысла рассматривать одновременно с эволюцией нервной системы. Это самостоятельная проблема, которая не имеет однозначных решений.
Можно предположить, что с переходом к придонному образу жизни с активным движением по субстрату у подобного предка хордовых могли появиться затруднения с дыханием. В воде придонной части океанов и
морей растворённого кислорода намного меньше, чем под поверхностью или в толще воды. Возникла потребность в усилении метаболизма. Если это произошло, то на спинной стороне кишечной трубки могли
сформироваться щели, которые играют роль жаберного аппарата (см. рис. II-15, в). Появление гомологов жабр у таких животных маловероятно, но полностью отрицать такую возможность нельзя, поскольку, не зная
размеров тела этого животного, мы не можем оценить его метаболизм.
Замена радиальной симметрии на билатеральную с неизбежностью приводит к возникновению одной или нескольких симметричных пар нервных тяжей, идущих вдоль тела.
Исходя из элементарных принципов эффективной работы нервной системы, мы можем остановиться на двух вариантах организации: 2 или 4 нервных тяжа (рис. II-16, б, д). Два нервных ствола могут располагаться на
боковых краях уплощённого и вытянутого тела животного (см. рис. II-16, б). Краевое положение нервных тяжей обусловлено тем, что каждый тяж имеет рецепторы, расположенные на поверхности тела, а боковые
поверхности являются основными контактными зонами. Чем ближе к ним будет расположен нервный тяж, тем быстрее будет реакция организма на оказываемое воздействие. Такое строение нервной системы характерно для многих плоских червей, хотя описываемое животное является исключительно гипотетической моделью и не имеет отношения к рецентным формам. Два нервных головных ганглия переходят в широко
расставленные нервные тяжи, расположенные по бокам тела животного. В центре расположен пищеварительный канал, дорсальнее и вентральнее которого проходят поперечные комиссуры, соединяющие левый и правый нервные тяжи (см. рис. II-16, б).
Первичноводные позвоночные
Тип позвоночных (Vertebrata) включает в себя подтип бесчерепных (Acrania) и подтип черепных (Craniata). Единственными представителями бесчерепных являются многочисленные ланцетники (Amphioxi), которые
рассматривались в предыдущем разделе, посвящённом происхождению позвоночных. Настоящий раздел охватывает обширную и разнообразную группу первичноводных черепных позвоночных. Они сформировались и долгое время морфологически изменялись в водной среде. В процессе эволюции именно этой группы сложилась нервная система, характерная для всех остальных позвоночных. Впервые возникло трубчатое строение нервной системы. Полости желудочков головного и спинного мозга стали непрерывными, появилась циркуляция внутрижелудочковой мозговой жидкости. В сочетании с эффективной кровеносной системой и
внутренним скелетом это создало потенциальную возможность увеличивать размеры нервной системы первых водных позвоночных практически без ограничений. Эти эволюционные перспективы наращивания размеров мозга и параллельного увеличения сложности поведения не были реализованы в водной среде. Потребовались особые условия выхода на сушу, которые стали стимулом для развития новых качеств мозга и его ассоциативных функций (Northcutt, Davis, 1983).
Однако благодаря первичноводным позвоночным нервная система приобрела те универсальные черты, которые послужили морфологической основой для дальнейшей эволюции. Его конструкция оказалась настолько эффективной, что по существу не подверглась принципиальным изменениям за всю историю эволюции позвоночных (Nieuwenhuys, 1982).
Конечно, мозг адаптивно перестраивался во всех систематических группах, но принципы структурной организации центральной и периферической нервной системы сохранились без изменений. На конструкцию нервной системы радикально не повлияли появление ни челюстного аппарата, ни плавников, ни конечностей, ни выход на сушу, ни адаптация к полёту, нии теплокровность.
Нервная система, как и позвоночник, оказалась тем органом, который обеспечивал выживание и прогресс всей группы позвоночных.
Морфофункциональные отделы головного мозга, как и их основные функции, остаются неизменными на всём протяжении эволюции от миноги до человека. Эта универсальность строения нервной системы говорит об
общности происхождения всех позвоночных. Делать предположения о полифилитичности происхождения позвоночных довольно трудно.
Допущение параллельного возникновения и эволюции сотен связанных между собой структур мозга весьма сомнительно.
Читайте также:
