
Нервная регуляция дыхания у животных
Под регуляцией дыхания понимают поддержание оптимального содержания кислорода и диоксида углерода в альвеолярном воздухе и в крови за счет изменения частоты и глубины дыхательных движений. Частота и глубина дыхательных движений обусловлены ритмом и силой генерации импульсов в дыхательном центре, расположенном в продолговатом мозге, в зависимости от его возбудимости. Возбудимость определяется напряжением диоксида углерода в крови и потоком импульсов с рецепторных зон сосудов, дыхательных путей, мышц.
Регуляция частоты дыхательных движений. Регуляция частоты дыхательных движений осуществляется центром дыхания, который включает в себя центры вдоха, выдоха и пневмотаксиса; центру вдоха принадлежит главная роль. В центре вдоха ритмически залпами рождаются импульсы в единицу времени, определяя частоту дыхания. Импульсы из центра вдоха поступают к вдыхательным мышцам и диафрагме, вызывая вдох такой продолжительности и глубины, который соответствует сложившимся условиям и характеризуется определенным объемом поступившего в легкие воздуха, силой сокращения вдыхательных мышц. Количество импульсов, рожденных в центре вдоха в единицу времени, зависит от его возбудимости: чем выше возбудимость, тем чаще рождаются импульсы, а значит, и чаще дыхательные движения.
Регуляция смены вдоха выдохом, выдоха вдохом. Регуляция смены вдоха выдохом, выдоха вдохом осуществляется рефлекторно. Возбуждение, возникающее в центре вдоха, обеспечивает акт вдоха, который сопровождается растяжением легких и возбуждением механорецепторов легочных альвеол. Импульсы с рецепторов по афферентным волокнам блуждающих нервов поступают уже в центр выдоха и возбуждают его нейроны. Одновременно непосредственно через центр пневмотаксиса центр вдоха также возбуждает центр выдоха. Нейроны центра выдоха, возбуждаясь, по законам реципрокных отношений тормозят активность нейронов центра вдоха, и вдох прекращается. Центр выдоха посылает информацию к мышцам экспираторам, вызывает их сокращение, и осуществляется акт выдоха. Так происходит чередование вдоха и выдоха. Количество залпов импульсов, поступающих из центра вдоха в единицу времени, и сила этих залпов зависят от возбудимости нейронов центра дыхания, специфики обмена веществ, особой чувствительности нейронов к окружающей их гуморальной среде, к поступающей информации с хеморецепторов сосудов, дыхательных путей и легких, мышц и пищеварительного аппарата. Избыток в крови и альвеолярном воздухе диоксида углерода и недостаток кислорода, усиление потребления кислорода и образования диоксида углерода в мышцах и других органах при усилении их деятельности вызывают следующие реакции: повышение возбудимости дыхательного центра, увеличение частоты рождения импульсов в центре вдоха, учащение дыхания и, как следствие, восстановление оптимального содержания кислорода и диоксида углерода в альвеолярном воздухе и крови. И наоборот, избыток в крови и альвеолярном воздухе кислорода ведет к урежению дыхательных движений и уменьшению вентиляции легких. В связи с приспособлением к изменившимся условиям число дыхательных движений у животных может увеличиться в 4 .5 раз, дыхательный объем воздуха в 4 .8 раз, минутный объем дыхания в 10 .25 раз.
Прочие статьи:
Яйцо и его особенности у птиц
Отложенное яйцо имеет плотную наружную оболочку — известковую скорлупу, прикрытую тонкой кутикулярной надскорлуповой оболочкой, предохраняющей яйцо от проникновения в него через поры в скорлупе различных микроорганизмов. Под скорлупой нах .
Самоорганизация в открытых неравновесных системах
Проблема самоорганизации материальных систем в XX веке становится одной из центральных проблем науки. Существенный вклад в решение этой проблемы вносит системный и информационный подходы. Терминология, выработанная в этих областях исследо .
В чём заключается явление катализа?
Катализ (от греч. κατάλυσις, восходит к καταλύειν — разрушение) — явление изменения скорости химической или биохимической реакции в присутствии веще .
Ритмические сокращения дыхательной мускулатуры обусловлено тем, что к ним из центральной нервной системы беспрерывно и ритмично поступают импульсы.
Исследованиями Н.А.Миславского (1885) установлено, что дыхательные движения непосредственно связаны с продолговатым мозгом. Отделение продолговатого мозга от спинного полностью прекращают дыхательные движения. Совокупность нейронов продолговатого мозга, деятельность которого может обеспечить ритмические движения называются дыхательным центром. Дыхательный центр расположен на дне четвертого желудочка в ретикулярной формации и состоит из правой и левой симметрически расположенных половин. Каждая его половина связана со своей стороной спинного мозга и состоит их центра вдоха и центра выдоха - это основной или “рабочий” дыхательный центр.
На изменения дыхательных движений оказывает влияние нейтроны спинного, продолговатого, среднего, промежуточного мозга и коры больших полушарий. В верхней части воролива моста находится так называемый пневмотоксический центр, который контролирует центр продолговатого мозга.
В регуляции дыхания участвует кора головного мозга, что доказывается выработкой условного рефлекса.
В организме происходит саморегуляция дыхания.
В легких, плевре, грудных и брюшных мышцах имеются окончания афферентных нервных волокон, которые возбуждаются во время вдоха и выдоха. Возбуждение передается в дыхательный центр продолговатого мозга и вызывает возбуждение центра вдоха или выдоха.
Во время вдоха, когда легкие в достаточной степени растягиваются, возникает механическое раздражение нервных окончаний в легких и плевре, возбуждение передается в продолговатый мозг, где центр вдоха тормозится, а центр выдоха возбуждается - вдох прекращается и начинается выдох.
При выдохе, когда легкие спадаются, раздражаются нервные окончания в легких и плевре, возбуждение передается в продолговатый мозг, где центр выдоха тормозит, а центр вдоха возбуждается, в результате чего выдох прекращается, наступает вдох. Следовательно, вдох рефлекторно регулирует выдох, а выдох регулирует вдох (теория Геренга-Брейера). При перерезке обоих блуждающих нервов дыхание сохраняется, но оно становится более глубоким и редким.
Дыхательный аппарат у птиц резко отличается от дыхательного аппарата млекопитающих животных.
Аппарат дыхания у птиц делится на воздухоносные пути, легкие и воздухоносные мешки. Грудная кость относительно большого размера и далеко заходит назад и вниз, прикрывая часть брюшных органов. Диафрагмы у птиц нет, легкие невелики и расположены под позвоночником в углублениях между ребрами. Легкие связаны с воздухоносными мешками и прочно прикреплены к ребрам и позвоночнику.
Всего у птиц четыре парные и один непарный воздухоносные мешки. Грудные и брюшные воздухоносные мешки располагаются между петлями кишечника, вокруг сердца и заходят отростками в трубчатые кости. Они создают резерв воздуха и облегчают вес птицы, но в газообмене не участвуют. У птиц газообмен происходит в легких.
При вдохе воздух поступает в трахею, бронхи, бронхиолы, воздухоносные капилляры, где происходит газообмен, и заполняет воздухоносные мешки. При выдохе воздух из воздухоносных мешков движется в обратном направлении, в легких снова происходит газообмен. Такая двукратная циркуляция воздуха через легкие обеспечивает интенсивный газообмен.
Контрольные вопросы: 1.Дыхание и его значение для организма. 2. Акт вдоха и выдоха. 3. Типы и частота дыхания у животных. 4. Жизненная емкость легких. 5. Значение верхних дыхательных путей. 6. Роль отрицательного давления в грудной полости в процессе дыхания. 7. Обмен газов в легких. 8. Состав вдыхаемого и выдыхаемого воздуха. 10. Перенос газов кровью. 11. Обмен газов в тканях. 12. Нервная регуляция дыхания. 13. Гуморальная регуляция дыхания. 14. Механизм первого вдоха. 15. Особенности дыхания у птиц.
Глава 5. Пищеварение
Пищеварение – это ряд сложных физиологических процессов, обеспечивающих размельчение и химическое расщепление питательных веществ корма в желудочно-кишечном тракте до низкомолекулярных соединений лишенных видовой специфичности и их всасывание в кровь и лимфу.
В пищеварительном тракте корм подвергается:
1) Физической обработке, благодаря чему он измельчается, увеличивается его поверхность, и он становится более доступным для его дальнейшего переваривания;
2) Биологической обработке, которая осуществляется микроорганизмами, населяющими в больших количествах пищеварительный тракт;
3) Химической обработке, которая происходит под действием ферментов, выполняющих роль биологических катализаторов химических процессов, обеспечивающих гидролиз питательных веществ, благодаря чему они превращаются в компоненты, лишенные видовой специфичности и доступные для их всасывания.
Органы пищеварения выполняют следующие функции:
1) Секреторную, заключающуюся в выработке пищеварительных соков содержащих ферменты;
2) Моторно-эвакуаторную или двигательную функцию, которая обеспечивает прием корма, его пережевывание, заглатывание, перемешивание, продвижение содержимого по длине пищеварительного тракта и выведение из организма непереваренных остатков корма.
3) Всасывательную - обеспечивающую поступление питательных веществ после их соответствующей переработки, в кровь и лимфу.
4) Экскреторную (выделительную) функцию, которая обеспечивает выведение из организма продуктов различных видов обмена веществ и непереваренных остатков корма.
5) Инкреторную - связанную с выработкой пищеварительными железами энтериновых гормонов и гормоноподобных веществ, влияющих не только на функции пищеварительного тракта, но и на другие системы организма.
6) Защитную, которая заключается в выполнении роли барьера, защищающего организм от проникновения вредных агентов.
7) Рецепторную (анализаторную) функцию, которая проявляется в оценке качества поступающего в организм корма.
В различных организмах в процессе их эволюционного развития образовалось несколько типов пищеварения. В зависимости от источника образования ферментов различают:
1. Собственное пищеварение — когда сам организм вырабатывает для себя пищеварительные ферменты.
2. Симбионтное пищеварение, осуществляемое ферментами микроорганизмов, что особенно большое значение имеет в пищеварении жвачных животных;
3. Аутолитическое — наиболее древний тип пищеварения происходит под влиянием ферментов, заключенных в самом корме.
В зависимости от места, где осуществляются пищеварительные процессы, выделяют внутриклеточный тип пищеварения, происходящий в пищеварительных вакуолях одноклеточных организмов.
Внеклеточное пищеварение можно разделить на полостное, осуществляемое в полости желудка, кишечника, и пристеночное или мембранное, протекающее на мембранах клеток слизистой оболочки пищеварительного тракта.
Кроме того, выделяют внешнее пищеварение, происходящее за пределами организма (например у пауков); коллективное, когда в этом процессе участвуют несколько особей многочисленного сожительства организмов (у пчел, муравьев).
Пищеварение в ротовой полости
Прием корма. Животные отыскивают корм и определяют его пищевую пригодность с участием органов зрения, обоняния, осязания, вкуса. Лошади, мелкий рогатый скот принимают корм главным образом хорошо подвижными губами и отрывают его резцами; крупный рогатый скот, свиньи — языком, губами; плотоядные — резцами и клыками; птицы — клювом. Питье воды происходит путем погружения в нее губной щели с последующим насасыванием движениями щек и языка. Плотоядные воду и жидкий корм лакают, птицы захватывают воду клювом, запрокидывают голову, чем облегчают ее заглатывание. Прием корма — акт произвольный и осуществляется по принципу цепных рефлексов, когда конец одного рефлекса является началом другого.
Жевание осуществляется разнообразными движениями нижней челюсти, благодаря чему корм измельчается, дробится, перетирается. В результате этого увеличивается его поверхность, он хорошо увлажняется слюной и становится доступным для проглатывания.
Жевание — акт рефлекторный, но произвольный. Возникшее от раздражения кормом рецепторов ротовой полости возбуждение по афферентным нервам (язычная ветвь тройничного нерва, языкоглоточный нерв, верхнегортанная веточка блуждающего нерва) передается в центр жевания продолговатого мозга. От него возбуждение по эфферентным волокнам тройничного, лицевого и подъязычного нервов поступает к жевательным мышцам и за счет их сокращения происходит акт жевания. С измельчением грубых частиц корма раздражение рецепторов ротовой полости уменьшается, в результате чего частота жевательных движений и их сила становятся более слабыми и направлены они теперь, главным образом, на формирование пищевого кома и подготовку его к глотанию. Высшие центры жевания располагаются в гипоталамусе и в моторной зоне коры головного мозга.
Слюнные железы. В ротовую полость впадают протоки трех пар слюнных желез – околоушных, подъязычных и нижнечелюстных.
1. Смачивает корм и облегчает его пережевывание.
2. Растворяя частицы корма, слюна участвует в определении его вкусовых качеств.
3. Слизистая часть слюны (муцин) склеивает мелкие частицы корма, формирует пищевой корм, ослизняет его и таким образом облегчает проглатывание.
4. За счет своей щелочности нейтрализует избыток кислот, образующихся в желудке.
5. Участвует в теплорегуляции у животных, не имеющих потовых желез. Например, у собак при высокой температуре с падающей изо рта слюной удаляется часть тепловой энергии.
6. Защитная роль слюны осуществляется за счет наличия в ней лизоцима, обладающего бактерицидными свойствами.
7. Слюна обладает низким поверхностным натяжением, содержит аскорбиновую кислоту и этим участвует в регуляции видового состава микрофлоры в преджелудках жвачных животных.
8. За счет присутствия в слюне мочевины и других азотсодержащих веществ она участвует в азотистом обмене.
9. Имея в своем составе пищеварительные ферменты, слюна способствует гидролизу углеводов в желудке.
10. Слюна содержит тромбопластические вещества, поэтому в какой то мере обладает кровеостанавливающим действием.
Слюноотделение — это сложнорефлекторный акт, осуществляемый вследствие раздражения механо-, хемо- и терморецепторов ротовой полости кормовыми или другими раздражающими веществами. Возбуждение от рецепторов по волокнам афферентных нервов передается в продолговатый мозг в центр слюновыделения и далее таламус, гипоталамус и кору головного мозга. От центра слюновыделения возбуждение по волокнам эфферентных симпатических и парасимпатических нервов переходит к слюнным железам и они начинают выделять слюну. Эфферентные парасимпатические волокна идут в составе лицевого и языкоглоточного нервов. Этот механизм выделения слюны определяется как безусловнорефлекторный. Парасимпатические влияния вызывают обильное выделение жидкой, водянистой слюны с небольшим содержанием в ней органических веществ. Симпатические нервы, напротив, уменьшают количество выделяемой слюны, но в ней содержится больше органических веществ. Регуляция количества выделения воды и органических веществ осуществляется нервным центром за счет различной информации, поступающей к нему по афферентным нервам. Слюна выделяется также при виде, запаха корма, в определенное время кормления животных и при других манипуляциях, связанных с предстоящим приемом корма. Это условнорефлекторный механизм выделения слюны.
Глотание — сложнорефлекторный акт, обеспечивающий эвакуацию корма из ротовой полости в пищевод.
Передвижение пищи по пищеводу осуществляется рефлекторно за счет перистальтических сокращений мышц пищевода. Началом этого рефлекса является акт глотания.
Пищеварение в желудке
Поступивший в желудок корм подвергается дальнейшей физической, химической и биологической обработке. Строение желудка, как и всего пищеварительного тракта, определяется особенностями питания животных. По своим анатомическим и функциональным особенностям различают простые однокамерные железистого типа желудки — у человека, собак, кошек; однокамерные усложненные желудки переходного желудочно-кишечного типа — у свиней, лошадей, кроликов; сложные многокамерные желудки — у жвачных животных и двухкамерные — у птиц. По особенностям строения слизистой оболочки и расположению в ней железистых клеток в простом однокамерном желудке выделяют три функционально различные зоны:
1. Кардиальная зона, примыкающая к пищеводу с расположенными в ней добавочными клетками, вырабатывающими слизистый секрет (слизь);
2. Донная (фундальная зона), включающая дно, тело и часть малой кривизны желудка с наличием в ней главных клеток, вырабатывающих ферменты, обкладочных клеток вырабатывающих соляную кислоту и небольшого количества добавочных клеток;
3. Пилорическая зона, прилегающая к пилорусу, содержит главные и добавочные клетки, вырабатывающие ферменты и слизь. Соляная кислота в пилорической части желудка не вырабатывается.
Состав желудочного сока.
Чистый желудочный сок представляет собой бесцветную, прозрачную жидкость кислой реакции (рН 0,8–1,2) с наличием небольшого количества слизи и клеток отторгнутого эпителия. Кислая реакция сока обусловлена наличием в нем соляной кислоты и других кислореагирующих соединений. В состав неорганической части сока входят минеральные вещества, имеющиеся в слюне. Органическая часть сока представлена белками, аминокислотами, ферментами, мочевиной, мочевой кислотой.
Ферменты желудочного сока
В желудочном соке выделено семь видов неактивных предшественников (проферментов), находящихся в клетках желудочных желез в виде гранул пепсиногенов, объединенных под общим названием пепсины. В полости желудка пепсиноген активируется HCI путем отщепления от него ингибирующего белкового комплекса.
Различают следующие основные пепсины:
1. Пепсин А-группа ферментов, гидролизирующих белки при рН - 1,5-2,0;
2. Пепсин С (желудочный катепсин) реализует свое действие при рН 3,2 -3,5;
3. Пепсин В (парапепсин, желатиназа) разжижает желатин, действует на белки соединительной ткани при рН менее 5,6;
4. Пепсин Д (реннин, химозин) действует в присутствии ионов кальция на казеиноген молока и переводит его в казеин, створаживая молоко.
К другим ферментам желудочного сока относятся желудочная липаза, расщепляющая эмульгированные жиры (жир молока) на глицерин и жирные кислоты.
Уреаза расщепляет мочевину при рН=8,0 до аммиака, который нейтрализует HCI.
Лизоцим (мурамидаза) обладает антибактериальным свойством.
Желудочная слизь и ее значение
Слизь вырабатывается всеми, но больше добавочными клетками слизистой оболочки желудка (мукоциты). Слой слизи предохраняет оболочку желудка от механических, химических, температурных повреждений, от ее самопереваривания (аутолиза) под действием HCI и пепсинов.
Значение соляной кислоты в пищеварении. Находясь в свободном и связанном состоянии, она выполняет большую роль в пищеварении:
1. Активирует превращение пепсиногена в пепсин и создает для его действия кислую среду;
2. Гормон просекретин переводит в активную форму секретин, влияющий на секрецию панкреатического сока;
3. Декальцинирует и этим разрыхляет кости;
4. Денатурирует белки, в результате чего они набухают, что облегчает их гидролиз;
5. Действует бактерицидно на гнилостную микрофлору;
6. Участвует в механизме перехода содержимого из желудка в кишечник;
7. Способствует створаживанию молока в желудке;
8. Активирует моторику желудка;
9. Активирует гормон прогастрин в гастрин, участвующий в регуляции желудочного соковыделения.
Механизм секреции желудочного сока
Секреция сока происходит под влиянием разнообразных внешних и внутренних стимуляторов. Условно различают три наслаивающихся друг на друга фазы выделения сока.
Сложнорефлекторная фаза первоначально связана с условнорефлекторными реакциями на раздражение зрительных, слуховых, обонятельных рецепторов, с которыми впоследствии присоединяются безусловнорефлекторные раздражения рецепторов ротовой полости, связанные с приемом корма и жеванием. Рефлекторная фаза продолжается 1–2 ч, доказывается опытом мнимого кормления (рис. 4), выделяющийся в это время желудочный сок, обладает высокой кислотностью и переваривающей силой.
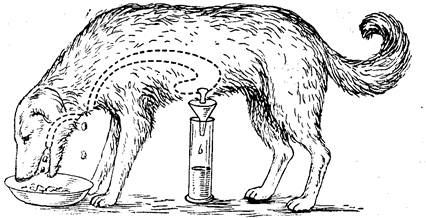
Рис.4. Мнимое кормление эзофаготомированной собаки с фистулой
На сложнорефлекторную фазу постепенно наслаивается желудочная (нервно-гуморальная) фаза. К продолжающемуся еще выделению сока от первой фазы на секрецию уже начинают влиять механические и химические факторы корма, а также гормоны гастрин, энтерогастрин, гистамин. Роль продуктов переваривания корма и других химических экстрактивных веществ в секреции сока доказывается опытом с незаметным для животного вкладыванием корма через фистулу непосредственно в желудок — в обход сложнорефлекторной фазы. В этих случаях соковыделение начинается только через 20–30 и более минут, когда появятся первые продукты гидролиза корма.
Кишечная фаза происходит при переходе содержимого из желудка в кишечник. Желудочная секреция в начале этой фазы еще увеличивается за счет химических веществ, всасывающихся в кишечнике, а затем она постепенно затухает вследствие образования в кишечнике секретина, который является антагонистом гастрина.
Регуляция дыхания - это согласованное нервное управление дыхательными мышцами, последовательно осуществляющими дыхательные циклы, состоящие из вдоха и выдоха.
Дыхательный центр - это сложное многоуровневое структурно-функциональное образование мозга, осуществляющее автоматическую и произвольную регуляцию дыхания.
Дыхание - процесс автоматический, но он поддается произвольной регуляции. Без такой регуляции невозможна была бы речь. Вместе с тем, управление дыханием построено на рефлекторных принципах: как безусловно-рефлекторных, так и условно-рефлекторных.
Регуляция дыхания построена на общих принципах автоматической регуляции, которые используются в организме.
Пейсмейкерные нейроны (нейроны - "создатели ритма") обеспечивают автоматическое возникновение возбуждения в дыхательном центре даже в том случае, если не будут раздражаться дыхательные рецепторы.
Тормозные нейроны обеспечивают автоматическое подавление этого возбуждения через определённое время.
В дыхательном центре используется принцип реципрокного (т.е. взаимоисключающего) взаимодействия двух центров: вдоха и выдоха. Их возбуждение находится в обратно пропорциональной зависимости. Это означает, что возбуждение одного центра (например, центра вдоха) тормозит связанный с ним второй центр (центр выдоха).
Функции дыхательного центра
- Обеспечение вдоха.
- Обеспечение выдоха.
- Обеспечение автоматии дыхания.
- Обеспечение приспособления параметров дыхания к условиям внешней среды и деятельности организма.
Например, при повышении температуры (как в окружающей среде, так и в организме) дыхание учащается.
Уровни дыхательного центра
1. Спинальный (в спинном мозге). В спинном мозге расположены центры, координирующие деятельность диафрагмы и дыхательных мышц - L-мотонейроны в передних рогах спинного мозга. Диафрагмальные нейроны - в шейных сегментах, межреберные - в грудных. При перерезке проводящих путей между спинным и головным мозгом дыхание нарушается, т.к. спинальные центры не обладают автономностью (т.е. самостоятельностью) и не поддерживают автоматию дыхания.
2. Бульбарный (в продолговатом мозге) - основной отдел дыхательного центра. В продолговатом мозге и варолиевом мосту располагаются 2 основных вида нейронов дыхательного центра - инспираторные (вдыхательные) и экспираторные (выдыхательные).
Инспираторные (вдыхательные) - возбуждаются за 0,01-0,02 с до начала активного вдоха. Во время вдоха у них увеличивается частота импульсов, а затем мгновенно прекращается. Подразделяются на несколько видов.
Виды инспираторных нейронов
По влиянию на другие нейроны:
- тормозные (прекращают вдох)
- облегчающие (стимулируют вдох).
По времени возбуждения:
- ранние (за несколько сотых долей секунды до вдоха)
- поздние (активны в процессе всего вдоха).
По связям с экспираторными нейронами:
- в бульбарном дыхательном центре
- в ретикулярной формации продолговатого мозга.
В дорсальном ядре 95% - инспираторные нейроны, в вентральном - 50%. Нейроны дорсального ядра связаны с диафрагмой, а вентрального - с межрёберными мышцами.
Экспираторные (выдыхательные) - возбуждение возникает за несколько сотых долей секунды до начала выдоха.
Различают:
- ранние,
- поздние,
- экспираторно-инспираторные.
В дорсальном ядре 5% нейронов являются экспираторными, а в вентральном - 50%. В целом экспираторных нейронов значительно меньше, чем инспираторных. Получается, что вдох важнее выдоха.
Автоматию дыхания обеспечивают комплексы из 4-х нейронов с обязательным присутствием тормозных.
Взаимодействие с другими центрами мозга
Дыхательные инспираторные и экспираторные нейроны имеют выход не только на дыхательные мышцы, но и на другие ядра продолговатого мозга. Например, при возбуждении дыхательного центра реципрокно тормозится центр глотания и в то же время, наоборот, возбуждается сосудо-двигательный центр регуляции сердечной деятельности.
На бульбарном уровне (т.е. в продолговатом мозге) можно выделить пневмотаксический центр, расположенный на уровне варолиева моста, выше инспираторных и экспираторных нейронов. Этот центр регулирует их активность и обеспечивает смену вдоха и выдоха. Инспираторные нейроны обеспечивают вдох и одновременно от них возбуждение поступает в пневмотаксический центр. Оттуда возбуждение бежит к экспираторным нейронам, которые возбуждаются и обеспечивают выдох. Если перерезать пути между продолговатым мозгом и варолиевым мостом, то уменьшится частота дыхательных движений, засчёт того, что уменьшается активирующее действие ПТДЦ (пневмотаксического дыхательного центра) на инспираторные и экспираторные нейроны. Это также приводит к удлинению вдоха засчёт длительного сохранения тормозного влияния экспираторных нейронов на инспираторные.
3. Супрапонтиальный (т.е. "надмостовый") - включает в себя несколько областей промежуточного мозга:
Гипоталамическая область - при раздражении вызывает гиперпноэ - увеличение частоты дыхательных движений и глубины дыхания. Задняя группа ядер гипоталамуса вызывает гиперпноэ, передняя группа действует противоположным образом. Именно засчёт дыхательного центра гипоталамуса дыхание реагирует на температуру окружающей среды.
Гипоталамус совместно с таламусом обеспечивает изменение дыхания при эмоциональных реакциях.
Таламус - обеспечивает изменение дыхания при болевых ощущениях.
Мозжечок - приспосабливает дыхание к мышечной активности.
4. Моторная и премоторная зона коры больших полушарий головного мозга. Обеспечивает условно-рефлекторную регуляцию дыхания. Всего за 10-15 сочетаний можно выработать дыхательный условный рефлекс. Засчёт этого механизма, например, у спортсменов перед стартом возникает гиперпноэ.
Асратян Э.А. в своих опытах удалял у животных эти области коры. При физической нагрузке у них быстро возникала одышка - диспноэ, т.к. им не хватало этого уровня регуляции дыхания.
Дыхательные центры коры дают возможность произвольного изменения дыхания.
Регуляция деятельности дыхательного центра
Бульбарный отдел дыхательного центра является главным, он обеспечивает автоматию дыхания, но его деятельность может изменяться под действием гуморальных и рефлекторных влияний.
Гуморальные влияния на дыхательный центр
Опыт Фредерика (1890). Он сделал перекрестное кровообращение у двух собак - голова каждой собаки получила кровь от туловища другой собаки. У одной собаки зажимали трахею, следовательно, возрастал уровень углекислого газа и понижался уровень кислорода в крови. После этого другая собака начинала часто дышать. Возникало гиперпноэ. В следствие этого в крови уменьшался уровень СО2 и возрастал уровень О2. Эта кровь поступала к голове первой собаки и тормозила ее дыхательный центр. Гуморальное торможение дыхательного центра могло довести эту первую собаку до апноэ, т.е. остановки дыхания.
Факторы, гуморально влияющие на дыхательный центр:
Избыток СО2 - гиперкарбия, вызывает активацию дыхательного центра.
Недостаток О2 - гипоксилия, вызывает активацию дыхательного центра.
Ацидоз - накопление ионов водорода (закисление), активирует дыхательный центр.
Недостаток СО2 - торможение дыхательного центра.
Избыток О2 - торможение дыхательного центра.
Алколоз - +++торможение дыхательного центра
Сами нейроны продолговатого мозга засчет высокой активности вырабатывают много СО2 и локально воздействуют на самих себя. Положительная обратная связь (сами себя усиливают).
Кроме прямого действия СО2 на нейроны продолговатого мозга существует рефлекторное действие через рефлексогенные зоны сердечно-сосудистой системы (рефлексы Рейманса). При гиперкарбии возбуждаются хеморецепторы и от них возбуждение поступает к хемочувствительным нейронам ретикулярной формации и к хемочувствительным нейронам коры головного мозга.
Рефлекторное влияние на дыхательный центр.
1. Постоянное влияние.
Рефлекс Гелинга-Брейера. Механорецепторы в тканях легких и дыхательных путей возбуждаются при растяжении и спадении легких. Они чувствительны к растяжению. От них импульсы по вакусу (блуждающий нерв) идет в продолговатый мозг к инспираторным L-мотонейронам. Вдох прекращается и начинается пассивный выдох. Этот рефлекс обеспечивает смену вдоха и выдоха и поддерживает активность нейронов дыхательного центра.
При перегрузке вакуса и перерезке рефлекс отменяется: снижается частота дыхательных движений, смена вдоха и выдоха осуществляется резко.
Другие рефлексы:
растяжение легочной ткани тормозит последующий вдох (экспираторно-облегчающий рефлекс).
Растяжение легочной ткани при вдохе сверх нормального уровня вызывает дополнительный вздох (парадоксальный рефлекс Хеда).
Рефлекс Гейманса - возникает от хеморецепторов сердечно-сосудистой системы на концентрацию СО2 и О2.
Рефлекторное влияние с пропреорецепторов дыхательных мышц - при сокращении дыхательных мышц возникает поток импульсов от пропреорецепторов к ЦНС. По принципу обратной связи изменяется активность инспираторных и экспираторных нейронов. При недостаточном сокращении инспираторных мышц возникает респираторно-облегчающий эффект и вдох усиливается.
2. Непостоянные
Ирритантные - расположены в дыхательных путях под эпителием. Являются одновременно механо- и хеморецепторами. Имеют очень высокий порог раздражения, поэтому работают в экстраординарных случаях. Например, при понижении легочной вентиляции объем легких уменьшается, возбуждаются ирритантные рецепторы и вызывают рефлекс форсированного вдоха. В качестве хеморецепторов эти же рецепторы возбуждаются биологически активными веществами - никотин, гистамин, простогландин. Возникает чувство жжения, першения и в ответ - защитный кашлевой рефлекс. В случае патологии ирритантные рецепторы могут вызвать спазм дыхательных путей.
в альвеолах рецепторы юкста-альвеолярные и юкста-капиллярные реагируют на объем легких и биологически активные вещества в капиллярах. Повышают частоту дыхания и сокращают бронхи.
На слизистых оболочках дыхательных путей - экстерорецепторы. Кашель, чихание, задержка дыхания.
На коже - тепловые и холодовые рецепторы. Задержка дыхания и активация дыхания.
Болевые рецепторы - кратковременная задержка дыхания, затем усиление.
Энтерорецепторы - с желудка.
Пропреорецепторы - со скелетных мышц.
Механорецепторы - с сердечно-сосудистой системы.
Читайте также:
