
Нервная регуляция обмена веществ и энергии
В организме человека обменные процессы регулируются эндокринной системой. Гормоны влияют на:
- активность ферментов (приводят ферменты в активную форму, тормозят их деятельность);
- синтез ферментов (воздействуют на генетический аппарат клетки);
- проницаемость мембран (инсулин усиливает проницаемость мембран к глюкозе);
Влияние нервной системы на обменные процессы составляет ее трофическую функцию. Основную роль играет симпатическая нервная система, медиатором которой является норадреналин. Медиаторы вмешиваются в метаболизм клетки, влияя на их активность.
Центральная нервная система оказывает влияние на обмен веществ, воздействуя на эндокринные железы. Особая роль при этом принадлежит гипоталамусу. Гипоталамус через симпатическую нервную систему осуществляет регуляцию некоторых желез внутренней секреции, кроме того вырабатывает нейрогормоны, стимулирующие деятельность передней доли гипофиза, а через нее – ряд периферических эндокринных желез.
Тепловой гомеостаз является основным условием жизнедеятельности организма. Фактором, обеспечивающим непрерывное течение метаболизма в разных органах и тканях, является определенная температура крови.
Обмен веществ, заключающийся в процессах ассимиляции и диссимиляции, состоит из трех этапов:
1. Поступление различных органических и неорганических веществ в организм;
2. Их изменение в организме;
3. Выведение продуктов распада.
В процессе обмена веществ непрерывно идет превращение энергии, переход потенциальной энергии химических соединений, освобождаемой при их расщеплении, в другие виды энергии, в основном тепловую и механическую.
На всех этапах обмена веществ и энергии оба эти процесса неотделимы друг от друга. Это два проявления единого процесса.
Химические превращения различных веществ, протекающие в клетках, тканях и органах, составляют межуточный обмен.
Последний (также как и обмен веществ в целом) разделяют на обмен белков, жиров, обмен углеводов, минеральных солей и воды. Они являются неразрывными частями единого биологического процесса. Однако при некоторых физиологических и патологических условиях наблюдаются изменения в отдельных видах обмена веществ: белкового или жирового, или минерального, или углеводного.
39. Энергетический баланс организма. Основной обмен условия определения значение для клиники. Особенности основного обмена у детей
Энергетический баланс – соотношение между расходом энергии организмом человека и поступлением ее за счет пищи.
Различают 3 вида энергетического баланса:
- энергетическое равновесие - расход энергии соответствует ее поступлению, такой вид баланса является физиологичным для здорового взрослого человека;
- отрицательный энергетический баланс - расход энергии превышает энергопоступление. Наблюдается при различных видах голодания и характеризуется мобилизацией всех ресурсов организма на продукцию энергии для ликвидации энергетического дефицита.
- Положительный энергетический баланс характеризуется превышением энергетической ценности пищевого рациона над расходом энергии. Этот вид баланса является физиологичным для детей, беременных, кормящих женщин и т.д.
Для определения энергозатрат организма пользуются различными лабораторными и расчетными(табличными) методами.
К лабораторным методам относят:
- Метод прямой калориметрии основан на измерении тепла, которое выделяет организм при различных видах деятельности. Для этой цели используют калориметрическую камеру, в которой определяют количество тепла, выделенного человеком при выполнении определенного вида работы.
- Метод непрямой калориметрии заключается в том, что окислительные процессы, происходящие в организме, связаны с потреблением кислорода и выделением углекислоты. С этой целью вычисляют дыхательный коэффициент - отношение между количеством выделяемого углекислого газа и количеством поглощенного кислорода в 1 мин. По величине дыхательного коэффициента, пользуясь специальной таблицей, находят величину энергетического эквивалента кислорода, а затем вычисляют количество израсходованнной энергии в единицу времени. Определяя расход энергии в состоянии покоя и при выполнении той или иной работы, по разности полученных величин находят затраты энергии на выполнение работы.
Основной обмен - это минимальное количество энергии, необходимое для поддержания нормальной жизнедеятельности организма в состоянии полного покоя при исключении всех внутренних и внешних влиянии, которые могли бы повысить уровень обменных процессов. Основной обмен веществ определяют утром натощак (через 12-14 ч после последнего приема пищи) в положении лежа на спине, при полном расслаблении мышц, в комфортных температурных условиях окружающей среды (18-20°С). Выражают основной обмен количеством энергии, выделенной организмом (кДж или ккал).
В состоянии полного физического и психического покоя организм расходует энергию на: 1) постоянно совершающиеся химические процессы; 2) механическую работу, выполняемую отдельными органами (сердце, дыхательные мышцы, кровеносные сосуды, кишечник и др.); 3) не прекращающуюся деятельность железисто-секреторного аппарата.
Интенсивность основного обмена веществ в жировой ткани в 3 раза ниже, чем в остальной клеточной массе организма. Худые люди производят больше тепла на 1 кг массы тела, чем полные. Если рассчитать энерговыделения на 1 м 2 поверхности тела, то эта разница почти исчезает, так как, согласно правилу Рубнера, основной обмен веществ приблизительно пропорционален поверхности тела для разных видов животных и человека.
У женщин основной обмен веществ ниже, чем у мужчин. Это связано с тем, что у женщин меньше масса и поверхность тела. Отмечены сезонные колебания величины основного обмена веществ - повышение его весной и снижение зимой. На величину основного обмена веществ влияет предшествующая мышечная работа. Мышечная деятельность вызывает повышение обмена веществ пропорционально тяжести выполняемой работы.
К значительным изменениям основного обмена приводят нарушения функций органов и систем организма. При повышенной функции щитовидной железы, малярии, брюшном тифе, туберкулезе, сопровождающихся лихорадкой, основной обмен веществ усиливается.

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).
Регуляция обмена веществ и энергии. Центр регуляции обмена веществ. Модуляторы.
В данной главе представлены общие вопросы нейрогуморальной регуляции обмена веществ и энергии в организме и, главным образом, регуляция метаболизма. Конечной целью регуляции обмена веществ и энергии является обеспечение потребностей организма, его органов, тканей и отдельных клеток в энергии и в разнообразных веществах в соответствии с уровнем функциональной активности. В целостном организме постоянно существует необходимость согласования общих метаболических потребностей с потребностями клетки органа, ткани. Такое согласование достигается посредством распределения между органами и тканями веществ, поступающих из окружающей среды и синтезированных внутри организма.
Обмен веществ, протекающий внутри организма, не связан непосредственно с окружающей средой. Питательные вещества, прежде чем они смогут вступить в обменные процессы, должны быть получены из пищи в желудочно-кишечном тракте в молекулярной форме. Кислород, необходимый для биологического окисления, должен быть получен из воздуха в легких, доставлен в кровь, связан с гемоглобином и перенесен кровью к тканям. Скелетные мышцы, являясь в организме одним из мощных потребителей энергии, также обслуживают обмен веществ и энергии, обеспечивая поиск, прием и обработку пищи. Непосредственное отношение к обмену веществ и энергии имеет выделительная система. Таким образом, регуляция обмена веществ и энергии является мультипараметрической, включающей в себя регулирующие системы множества функций организма (например, дыхания, кровообращения, выделения, теплообмена и др.).
Роль центра в регуляции обмена веществ и энергии играют ядра гипоталамуса. Они имеют непосредственное отношение к генерации чувства голода и насыщения, теплообмену, осморегуляции. В гипоталамусе имеются полисенсорные нейроны, реагирующие на изменения концентрации глюкозы, водородных ионов, температуры тела, осмотического давления, т. е. важнейших гомеостатических констант внутренней среды организма. В ядрах гипоталамуса осуществляется анализ состояния внутренней среды и формируются управляющие сигналы, которые посредством эфферентных систем приспосабливают ход метаболизма к потребностям организма.
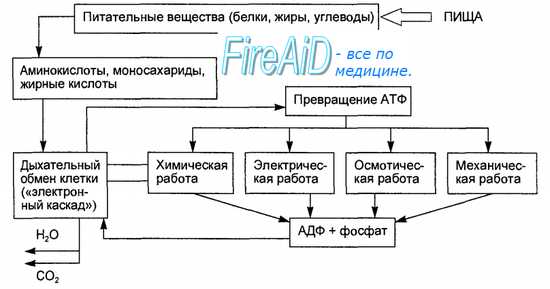
В качестве звеньев эфферентной системы регуляции обмена используются симпатический и парасимпатический отделы вегетативной нервной системы. Вьщеляющиеся их нервными окончаниями медиаторы оказывают прямое или опосредованное вторичными посредниками влияние на функцию и метаболизм тканей. Под управляющим влиянием гипоталамуса находится и используется в качестве эфферентной системы регуляции обмена веществ и энергии — эндокринная система. Гормоны гипоталамуса, гипофиза и других эндокринных желез оказывают прямое влияние на рост, размножение, дифференцировку, развитие и другие функции клеток. Гормоны принимают участие в поддержании в крови необходимого уровня таких веществ, как глюкоза, свободные жирные кислоты, минеральные вещества.
Химическая энергия питательных веществ используется для ресинтеза АТФ, выполнения всех видов работы и процессы, протекающие внутри клетки. Поэтому важнейшим эффектором, через который оказывается регулирующее воздействие на обмен веществ и энергии, являются клетки органов и тканей. Регуляция обмена веществ заключается в воздействии на скорость биохимических реакций, протекающих в клетках.
Метаболизм клетки в целом невозможен без интеграции многих биохимических превращений. Эта интеграция обеспечивается, главным образом, с помощью аденилатов, участвующих в регуляции любых метаболических превращений клетки.
Интеграция обмена белков, жиров и углеводов клетки осуществляется посредством общих для них источников энергии. При биосинтезе любых простых и сложных органических соединений, макромолекул и надмолекулярных структур в качестве общих источников энергии используется АТФ, которая поставляет энергию для процессов фосфорилирования, или НАД • Н, НАДФ • Н, поставляющих энергию для восстановления окисленных соединений других веществ. За общий энергетический запас клетки, полученный в ходе катаболизма, конкурируют все анаболические процессы, протекающие с затратой энергии. Так, например, при осуществлении печенью синтеза глюкозы из лактата и аминокислот (глюконеогенез) она не может одновременно синтезировать жиры и белки. Глюконеогенез сопровождается расщеплением в печени белков и жиров и окислением образующихся при этом жирных кислот, что ведет к освобождению энергии, необходимой для синтеза АТФ и НАД- Н, необходимых для глюконеоге-неза.
Еще одним проявлением интеграции метаболических превращений белков, жиров и углеводов в клетке является существование общих предшественников и общих промежуточных продуктов обмена веществ. Общим промежуточным продуктом обмена является ацетил-КоА. Важнейшими конечными путями превращений веществ в клетке являются цикл лимонной кислоты и реакции дыхательной цепи, протекающие в митохондриях. Цикл лимонной кислоты — главный источник С02 для последующих реакций глюконеогенеза, синтеза жирных кислот и мочевины.
Одним из механизмов согласования общих метаболических потребностей организма с потребностями клетки являются нервные и гормональные влияния на ключевые ферменты. Характерными особенностями этих ферментов являются: положение в начале того метаболического пути, к которому принадлежит фермент; приближенность расположения или ассоциированность со своим субстратом; реагирование не только на действие внутриклеточных регуляторов метаболизма, но и на внеклеточные нервные и гормональные воздействия.
Процесс гликогенолиза в мышцах может одновременно активироваться нервной системой и катехоламинами. Этот эффект достигается с участием ионов Са2+, который связывается с кальмодулином, являющимся субъединицей фосфорилазы. Она при этом активируется и приводит к мобилизации гликогена. Нервный механизм мобилизации гликогена осуществляется через меньшее число промежуточных этапов, чем гормональный. Этим достигается его быстродействие.
Удовлетворение энергетических потребностей организма посредством ускорения внутриклеточных процессов расщепления триглицеридов в жировой клетчатке достигается активацией гормончувствительной липазы. Повышение активности этого фермента (адреналином, норадреналином, глюкагоном) приводит к мобилизации свободных жирных кислот, являющихся основным энергетическим субстратом окисления в мышцах при выполнении ими интенсивной и длительной работы.
Переход органов и тканей с одного уровня функциональной активности на другой всегда сопровождается соответствующими изменениями их трофики (питания). Например, при рефлекторном сокращении скелетных мышц нервная система осуществляет не только пусковое действие, но и трофическое влияние путем усиления в них местного кровотока и интенсивности обмена веществ. Увеличение силы сокращений миокарда под влиянием симпатической нервной системы обеспечивается одновременным усилением коронарного кровотока и метаболизма в мышце сердца. О влиянии нервной системы на трофику скелетных мышц свидетельствует тот факт, что денервация мышцы приводит к постепенной атрофии мышечных волокон. Важнейшее значение в осуществлении трофической функции нервной системы играет ее симпатический отдел. Через симпато-адреналовую систему достигается не только активация обмена веществ и энергии в клетке.
Норадреналин и адреналин, выброс которых в кровоток возрастает при возбуждении симпатической нервной системы, вызывают увеличение глубины дыхания, расширяют мускулатуру бронхов, что способствует доставке кислорода в кровь. Адреналин, оказывая положительное инотропное и хронотропное действие на сердце, увеличивает минутный объем крови, повышает систолическое артериальное давление. В результате активации дыхания и кровообращения возрастает доставка кислорода к тканям.
В регуляции обмена веществ и энергии выделяют регуляцию обмена организма веществами и энергией с окружающей средой и регуляцию метаболизма в самом организме.
Регуляция обмена организма с окружающей средой питательными веществами рассматривается в главе 9.
Вопросы регуляции водно-солевого обмена описаны в главе 12. Регуляция обмена организма с окружающей средой теплом, как конечной формой превращения всех видов энергии, обсуждается в главе 11.
Поэтому здесь представлены общие вопросы нейрогуморальной регуляции обмена веществ и энергии в организме и, главным образом, регуляция метаболизма целостного организма.
Конечной целью регуляции обмена веществ и энергии является удовлетворение в соответствии с уровнем функциональной активности потребностей целостного организма, его органов, тканей и отдельных клеток в энергии и разнообразных пластических веществах. В целостном организме постоянно существует необходимость согласования общих метаболических потребностей организма с потребностями клетки органа, ткани. Такое согласование достигается посредством распределения между органами и тканями веществ, поступающих из окружающей среды, и перераспределения между ними веществ, синтезирующихся внутри организма.
Обмен веществ, протекающий внутри организма, не связан прямыми способами с окружающей средой. Питательные вещества,
Роль центра в регуляции обмена веществ и энергии играет гипоталамус. Это обусловлено тем, что в гипоталамусе локализованы нервные ядра и центры, имеющие непосредственное отношение к регуляции голода и насыщения, теплообмена, осморегуляции. В гипоталамусе идентифицированы полисенсорные нейроны, реагирующие сдвигами функциональной активности на изменения концентрации глюкозы, водородных ионов, температуры тела, осмотического давления, т.е. важнейших гомеостатических констант внутренней среды организма. В ядрах гипоталамуса осуществляется анализ состояния внутренней среды организма и формируются управляющие сигналы, которые посредством эфферентных систем приспосабливают ход метаболизма к потребностям организма.
В качестве звеньев эфферентной системы регуляции обмена используется симпатический и парасимпатический отделы вегетативной нервной системы. Выделяющиеся их нервными окончаниями медиаторы оказывают прямое или опосредованное вторичными посредниками влияние на функцию и метаболизм тканей. Под управляющим влиянием гипоталамуса находится и используется в качестве эфферентной системы регуляции обмена веществ и энергии — эндокринная система. Гормоны гипоталамуса, гипофиза и других эндокринных желез оказывают прямое влияние на рост, размножение, дифференцировку, развитие и другие функции клеток. Гормоны принимают участие в поддержании в крови необходимого уровня таких веществ, как глюкоза, свободные жировые кислоты, минеральные ионы (см. главу 5).
Обмен веществ (анаболизм и катаболизм), получение запасаемой в макроэргических связях АТФ энергии, выполнение различных видов работ с использованием метаболической энергии — это, как правило, процессы, протекающие внутри клетки. Поэтому важнейшим эффектором, через который можно оказать регулирующее воздействие на обмен веществ и энергии, является клетка органов и тканей. Регуляция обмена веществ заключается в воздействии на скорость биохимических реакций, протекающих в клетках.
Наиболее частыми эффектами регуляторных воздействий на клетку являются изменения: каталитической активности ферментов и их концентрации, сродства фермента и субстрата, свойств микросреды,
Метаболизм клетки в целом невозможен без интеграции многих биохимических превращений и сама возможность его осуществления определяется энергетическим и окислительно-восстановительным потенциалом клетки. Эта общая интеграция метаболизма обеспечивается, главным образом, с помощью аденилатов, участвующих в регуляции любых метаболических превращений клетки.
Интеграция обмена белков, жиров и углеводов клетки осуществляется посредством общих для них источников энергии. Действительно, при биосинтезе любых простых и сложных органических слоединений, марокмолекул и надмолекулярных структур в качестве общих источников энергии используется АТФ, которая поставляет энергию для процессов фосфорилирования, или НАД • Н, НАДФ • Н, поставляющие энергию для восстановления окислительных соединений. Таким образом, если в клетке осуществлять синтез (анаболизм) определенных веществ, то он может происходить за счет затраты химической энергии одного из общих подвижных источников (АТФ, НАД • Н, НАДФ-Н), которые образуются при катаболизме других веществ (см.рис.10.1).
За общий энергетический запас клетки, полученный в ходе катаболизма и являющийся движущей силой разнообразных превращений, конкурируют все анаболические и другие процессы, протекающие с затратой энергии. Так, например, осуществление глюкоста-тической функции печени, основанной на способности печени синтезировать глюкозу из лактата и аминокислот <глюконеогенез), несовместимо с одновременным синтезом жиров и белков. Глюконеогенез сопровождается расщеплением в печени белков и жиров и окислением образующихся при этом жирных кислот, что ведет к освобождению энергии, необходимой для синтеза АТФ и НАД • Н, в свою очередь требующихся для глюконеогенеза.
Еще одним проявлением интеграции метаболических превращений белков, жиров и углеводов является существование общих предшественников и общих промежуточных продуктов обмена веществ. Это — общий фонд углерода, общий промежуточный продукт обмена- ацетил- КоА и другие вещества. Важнейшими конечными путями превращений, связующими метаболические процессы на различных этапах, являются цикл лимонной кислоты и реакции дыхательной цепи, протекающие в митохондриях. Так, цикл лимонной кислоты — главный источник СО2 для последующих реакций глюконеогенеза, синтеза жирных кислот и мочевины.
Одним из механизмов согласования общих метаболических потребностей организма с потребностями клетки являются нервные и
Примерами ключевых ферментов являются гликогенфосфорилаза, фосфофруктокиназа, липаза. Их роль в процессах регуляции метаболизма видна, в частности, при подготовке организма к "борьбе или бегству". При повышении в этих условиях в крови уровня адреналина до 10 -9 М он связывается с адренорецепторами плазматической мембраны, активирует аденилатциклазу, которая катализирует превращение АТФ в циклический АМФ. Последний активирует гликогенфосфорилазу, многократно усиливающую расщепление гликогена в печени.
Процесс гликогенолиза в мышцах может одновременно активироваться нервной системой и катехоламинами. Этот эффект достигается посредством выделения ионов Са ++ , его связывания с кальмо-дулином, являющимся субъединицей фосфорилазы, которая при этом активируется и приводит к мобилизации гликогена. Нервный механизм мобилизации гликогена осуществляется через меньшее число промежуточных этапов, чем гормональный. Этим достигается его быстродействие.
Удовлетворение энергетических потребностей организма посредством ускорения внутриклеточных процессов расщепления триглице-ридов в жировой клетчатке достигается активацией гормончувстви-тельной липазы. Повышение активности этого фермента (адреналином, норадреналином, глюкагоном) приводит к мобилизации свободных жирных кислот, являющихся основным энергетическим субстратом окисления в мышцах при выполнении ими интенсивной и длительной работы.
Переход органов и тканей с одного уровня функциональной активности на другой всегда сопровождается соответствующими изменениями их трофики. Например, при рефлекторном сокращении скелетных мышц нервная система осуществляет не только пусковое действие, но и трофическое путем усиления в них местного кровотока и интенсивности обмена веществ. Увеличение силы сокращений миокарда под влиянием симпатической нервной системы обеспечивается одновременным усилением коронарного кровотока и метаболизма в мышце сердца. О влиянии нервной системы на трофику скелетных мышц свидетельствует тот факт, что денервация мышцы приводит к постепенной атрофии мышечных волокон. Важнейшее значение в осуществлении трофической функции нервной системы играет ее симпатический отдел. Через симпато-адреналовую систему достигается не только активация обмена веществ и энергии в клетке, но и создаются дополнительные условия для ускорения метаболизма. Норадреналин и адреналин, выброс которых в кровоток возрастает при возбуждении симпатической нервной системы,
Одним из интегральных показателей внутренней среды, отражающим обмен в организме углеводов, белков и жиров, является концентрация в крови глюкозы. Глюкоза является не только энергетическим субстратом, необходимым для синтеза жиров и белков, но и источником их синтеза. В печени происходит новообразование углеводов из жирных кислот и аминокислот.
Нормальное функционирование клеток нервной системы, мышц, для которых глюкоза является важнейшим энергосубстратом, возможно при условии, что приток к ним глюкозы обеспечит их энергетические потребности. Это достигается при содержании в литре крови у человека в среднем 1 г (0,8-1,2 г) глюкозы (рис. 10.3.).
При снижении содержания глюкозы в литре крови до уровня менее 0,5 г, вызванном голоданием, передозировкой инсулина, имеет место недостаточность снабжения энергией клеток мозга. Нарушение их функций проявляется учащением сердцебиения, слабостью и тремором мышц, головокружением, усилением потоотделения, ощущением голода. При дальнейшем снижении концентрации глюкозы в крови указанное состояние, именуемое гипогликемией, может перейти в гипогликемическую кому, характеризующуюся угнетением функций мозга вплоть до потери сознания. Введение в кровь глюкозы, прием сахарозы, инъекция глюкагона предупреждают или ослабляют эти проявления гипогликемии.
В нормальных условиях во всей крови организма содержится около 5 г глюкозы. При среднесуточном потреблении с пищей взрослым человеком, занимающимся физическим трудом, 430 г углеводов в условиях относительного покоя, тканями ежеминутно потребляется около 0,3 г глюкозы. При этом запасов глюкозы в циркулирующей крови достаточно для питания тканей на 3-5 минут и без ее восполнения неминуема гипогликемия. Потребление глюкозы возрастает при физической и психоэмоциональной нагрузках. Так как периодический (несколько раз в день) прием углеводов с пищей не обеспечивает постоянного и равномерного притока глюкозы из кишечника в кровь, в организме существуют механизмы, восполняющие убыль глюкозы из крови в количествах, эквивалентных ее потреблению тканями. Механизмы с другой направленностью действия обеспечивают в нормальных условиях превращение глюкозы в запасаемую форму — гликоген. При уровне более 1,8 г в литре крови происходит выведение ее из организма с мочой.
Избыток глюкозы, всосавшейся из кишечника в кровь воротной вены, поглощается гепатоцитами. При повышении в них концент-
Рис. 10.3 Система регуляции уровня глюкозы в крови (Пояснения в тексте)
Содержание глюкозы в крови оценивается глюкорецепторами переднего гипоталамуса, а также его полисенсорными нейронами. В ответ на повышении уровня глюкозы в крови выше "заданного значения" (>1,2 г/л) повышается активность нейронов гипоталамуса, которые посредством влияния парасимпатической нервной системы на поджелудочную железу усиливают секрецию инсулина.
При понижении уровня глюкозы в крови уменьшается ее поглощение гепатоцитами. В поджелудочной железе снижается секреторная активность В-клеток, уменьшается секреция инсулина. Тормозятся процессы превращения глюкозы в гликоген в печени и мышцах, уменьшается поглощение и усвоение глюкозы скелетными и гладкими мышцами, жировыми клетками. При участии этих механизмов замедляется или предотвращается дальнейшее понижение уровня глюкозы в крови, которое могло бы привести к развитию гипогликемии.
При уменьшении концентрации глюкозы в крови имеет место повышении тонуса симпатической нервной системы. Под ее влиянием усиливается секреция в мозговом веществе надпочечников адреналина и норадреналина. Адреналин, стимулируя распад гликогена в печени и мышцах вызывает повышение концентрации сахара в крови. По этому свойству адреналин является наиболее важным антагонистом инсулина среди других гормонов системы регуляции уровня сахара в крови. Например, норадреналин обладает слабовы-раженной способностью повышать уровень глюкозы в крови.
Под влиянием симпатической нервной системы стимулируется выработка а-клетками поджелудочной железы глюкагона, который активирует распад гликогена печени, стимулирует глюконеогенез и приводит к повышению уровня глюкозы в крови.
Понижение в крови концентрации глюкозы, являющейся для организма одним из наиболее важных энергетических субстратов, вызывает развитие стресса. В ответ на снижение уровня сахара крови глюкорецепторные нейроны гипоталамуса через рилизинг-гормоны стимулируют секрецию гипофизом в кровь гормона роста и адрено-кортикотропного гормона. Под влиянием гормона роста уменьшается проницаемость клеточных мембран для глюкозы, усиливается глю-
Секретируемые под действием адренокортикотропного гормона в коре надпочечников глкжокортикоиды активируют ферменты глюко-неогенеза в печени и этим способствуют увеличению содержания сахара в крови. Одновременно под действием глкжокортикоидов уменьшается включение аминокислот в белки и увеличивается скорость выведения из организма азота. Глкжокортикоиды повышают эффективность липолиза в жировой ткани и мобилизации в кровь свободных жирных кислот.
Регуляция обмена веществ и энергии в целостном организме находится под контролем нервной системы и ее высших отделов. Об этом свидетельствуют факты условнорефлекторного изменения интенсивности метаболизма у спортсменов в предстартовом состоянии, у рабочих перед началом выполнения тяжелой физической работы, у водолазов перед их погружением в воду. В этих случаях увеличивается скорость потребления организмом кислорода, возрастает минутный объем дыхания, минутный объем кровотока, усиливается энергообмен.
Развивающееся при снижении в крови содержания глюкозы, свободных жирных кислот, аминокислот чувство голода обусловливает поведенческую реакцию, направленную на поиск и прием пищи и восполнение в организме питательных веществ.
Условнорефлекторные изменения обмена веществ и энергии наблюдаются у человека в предстартовых и предрабочих состояниях. У спортсменов до начала соревнования, а у рабочего перед работой отмечается повышение обмена веществ, температуры тела, увеличивается потребление кислорода и выделение углекислого газа. Можно вызвать условнорефлекторные изменения обмена веществ, энергетических и тепловых процессов улюдей на словесный раздражитель.
Влияние нервной системы на обменные и энергетические процессы в организме осуществляется несколькими путями:
- Непосредственное влияние нервной системы (через гипоталамус, эфферентные нервы) на ткани и органы;
- опосредованное влияние нервной системы через гипофиз (соматотропин);
- опосредованное влияние нервной системы через тропные гормоны гипофиза и периферические железы внутренней секреции;
-прямоевлияниенервной системы (гипоталамус) на активность желез внутренней секреции и через них на обменные процессы в тканях и органах.
Основным отделом центральной нервной системы, который регулирует все виды обменных и энергетических процессов, является гипоталамус. Выраженное влияние на обменные процессы и теплообразование оказываютжелезы внутренней секреции. Гормоны коры надпочечников и щитовидной железы в больших количествах усиливают катаболизм, т. е. распад белков.
В организме ярко проявляется тесное взаимосвязанное влияние нервной и эндокринной систем на обменные и энергетические процессы. Так, возбуждение симпатической нервной системы не только оказывает прямое стимулирующее влияние на обменные процессы, но при этом увеличивается секреция гормонов щитовидной железы и надпочечников (тироксин и адреналин). За счет этого дополнительно усиливается обмен веществ и энергии. Кроме того, эти гормоны сами повышают тонус симпатического отдела нервной системы. Значительные изменения в метаболизме и теплообмене происходят при дефиците в организме гормонов желез внутренней секреции. Например, недостаток тироксина приводит к снижению основного обмена. Это связано с уменьшением потребления кислорода тканями и ослаблением теплообразования. В результате снижается температура тела.
Гормоны желез внутренней секреции участвуют в регуляции обмена веществ и энергии, изменяя проницаемость клеточных мембран (инсулин), активируя ферментные системы организма (адреналин, глюкагон и др.) и влияя на их биосинтез (глюкокортикоиды).
Таким образом, регуляция обмена веществ и энергии осуществляется нервной и эндокринной системами, которые обеспечивают приспособление организма к меняющимся условиям его обитания.
Биологическая роль белков
Белок, являясь важнейшим компонентом питания, обеспечивающим пластические и энергетические нужды организма, справедливо назван протеином, показывающим первую его роль в питании. Роль белков в питании человека трудно переоценить. Сама жизнь является одним из способов существования белковых тел. Биологическая роль белков
Белок можно отнести к жизненно важным пищевым веществам, без которых невозможны жизнь, рост и развитие организма. Достаточность белка в питании и высокое его качество позволяют создать оптимальные условия внутренней среды для нормальной жизнедеятельности организма, его развития и высокой работоспособности. Белок является главной составной частью пищевого рациона, определяющей характер питания. На фоне высокого уровня белка отмечается наиболее полное проявление в организме биологических свойств других компонентов питания. Белки обеспечивают структуру и каталитические функции ферментов и гормонов, выполняют защитные функции, участвуют в образовании многих важных структур белковой природы: иммунных тел, специфических ?-глобулинов, белка крови пропердина, играющего известную роль в создании естественного иммунитета, участвуют в образовании тканевых белков, таких как миозин и актин, обеспечивающих мышечные сокращения, глобина, входящего в состав гемоглобина эритроцитов крови и выполняющего важнейшую функцию дыхания. Белок, образующий зрительный пурпур (родопсин) сетчатки глаза, обеспечивает нормальное восприятие света, и др.
Следует отметить, что белки определяют активность многих биологически активных веществ: витаминов, а также фосфолипидов, отвечающих за холестериновый обмен. Белки определяют активность тех витаминов, эндогенный синтез которых осуществляется из аминокислот. Например, из триптофана – витамина PР (никотиновая кислота), обмен метионина – связан с синтезом витамина U (метилметионин-сульфоний). Установлено, что белковая недостаточность может привести к недостаточности витамина С и биофлаваноидов (витамина Р). Нарушение в печени синтеза холина (группы витаминоподобных веществ) приводит к жировой инфильтрации печени.
При больших физических нагрузках, а также при недостаточном поступлении жиров и углеводов белки участвуют в энергетическом обмене организма.
Наиболее тяжелые последствия в состоянии здоровья, нередко на всю жизнь, оставляет такой вид недостаточности питания, как алиментарная дистрофия, чаще всего возникающая при отрицательном энергетическом балансе, когда в энергетические процессы включаются не только пищевые химические вещества, поступающие с пищей, но и собственные, структурные белки организма. В алиментарной дистрофии выделяют отечную и безотечную формы с явлениями или без явлений витаминной недостаточности.
Может сложиться впечатление, что заболевания алиментарного характера возникают только при недостаточном поступлении белка в организм. Это не совсем так! При избыточном поступлении белка у детей первых трех месяцев жизни появляются симптомы дегидратации, гипертермии и явления обменного ацидоза, что резко увеличивает нагрузку на почки. Обычно это возникает, когда при искусственном вскармливании используют неадаптированные молочные смеси, негуманизированные типы молока.
Обменные нарушения в организме могут появиться и при несбалансированности аминокислотного состава поступающих белков.
Заменимые и незаменимые аминокислоты, значение и потребность в них
В настоящее время известно 80 аминокислот, наибольшее значение в питании имеют 30, которые наиболее часто встречаются в продуктах и чаще всего потребляются человеком. К ним относятся следующие.
1. Алифатические аминокислоты:
а) моноаминомонокарбоновые – глицин, аланин, изолейцин, лейцин, валин;
б) оксимоноаминокарбоновые – серин, треонин;
в) моноаминодикарбоновые – аспаргиновая, глютаминовая;
г) амиды моноаминодикарбоновых кислот – аспарагин, глутамин;
д) диаминомонокарбоновые – аргинин, лизин;
е) серосодержащие – гистин, цистеин, метионин.
2. Ароматические аминокислоты: фенилаланин, тирозин.
3. Гетероциклические аминокислоты: триптофан, гистидин, пролин, оксипролин.
Наибольшее значение в питании представляют незаменимые аминокислоты, которые не могут синтезироваться в организме и поступают только извне – с продуктами питания. К их числу относят 8 аминокислот: метионин, лизин, триптофан, треонин, фенилаланин, валин, лейцин, изолейцин. В эту группу входят и аминокислоты, которые в детском организме не синтезируются или синтезируются в недостаточном количестве. Прежде всего это гистидин. Предметом дискуссий является также вопрос о незаменимости в детском возрасте глицина, цистина, а у недоношенных детей также глицина и тирозина. Биологическая активность гормонов АКТГ, инсулина, а также коэнзима А и глютатиона определена наличием в их составе SH-групп цистина. У новорожденных детей из-за недостатка цистеназы лимитирован переход метионина в цистин. В организме взрослого человека тирозин легко образуется из фенилаланина, а цистин – из метионина, однако обратной заменяемости нет. Таким образом, можно считать, что число незаменимых аминокислот составляет 11—12.
Поступающий белок считается полноценным, если в нем присутствуют все незаменимые аминокислоты в сбалансированном состоянии. К таким белкам по своему химическому составу приближаются белки молока, мяса, рыбы, яиц, усвояемость которых около 90 %. Белки растительного происхождения (мука, крупа, бобовые) не содержат полного набора незаменимых аминокислот и поэтому относятся к разряду неполноценных. В частности, в них содержится недостаточное количество лизина. Усвоение таких белков составляет, по некоторым данным, 60 %.
Для изучения биологической ценности белков используют две группы методов: биологические и химические. В основе биологических лежит оценка скорости роста и степени утилизации пищевых белков организмом. Данные методы являются трудоемкими и дорогостоящими.
Химический метод колоночной хроматографии позволяет быстро и объективно определить содержание аминокислот в пищевых белках. На основании этих данных биологическую ценность белков определяют путем сравнения аминокислотного состава изучаемого белка со справочной шкалой аминокислот гипотетического идеального белка или аминограмм высококачественных стандартных белков. Этот методический прием получил название аминокислотного СКОРА = отношению количества АК в мг в 1 г исследуемого белка к количеству АК в мг в 1 г идеального белка, умноженного на 100 %.
Важным показателем качества пищевого белка служит также степень его усвояемости. По степени переваривания протеолитическими ферментами пищевые белки располагаются следующим образом:
1) белки рыбы и молока;
3) белки хлеба и круп.
Белки рыбы лучше усваиваются из-за отсутствия в их составе белка соединительной ткани. Белковая полноценность мяса оценивается по соотношению между триптофаном и оксипролином. Для мяса высокого качества это соотношение составляет 5,8.
Каждая аминокислота из группы эссенциальных играет определенную роль. Их недостаток или избыток ведет к каким-либо изменениям в организме.
Биологическая роль незаменимых аминокислот
Гистидин играет важную роль в образовании гемоглобина крови. Недостаток гистидина приводит к снижению уровня гемоглобина в крови. При декарбоксилировании гистидин превращается в гистамин – вещество, имеющее большое значение в расширении сосудистой стенки и ее проницаемости, влияет на выделение желудочного пищеварительного сока. Недостаток гистидина, так же как и избыток, ухудшает условно-рефлекторную деятельность.
Валин – физиологическая роль данной НАК недостаточно ясна. При недостаточном поступлении у лабораторных животных отмечаются расстройства координации движений, гиперестезия.
Изолейцин наряду с лейцином входит в состав всех белков организма (за исключением гемоглобина). В плазме крови содержится 0,89 мг% изолейцина. Отсутствие изолейцина в пище приводит к отрицательному азотистому балансу, к замедлению процессов роста и развития.
Лизин относится к одной из наиболее важных незаменимых аминокислот. Он входит в триаду аминокислот, особенно учитываемых при определении общей полноценности питания: триптофан, лизин, метионин. Оптимальное соотношение этих аминокислот составляет: 1 : 3 : 2 или 1 : 3 : 3, если взять метионин + цистин (серосодержащие аминокислоты). Недостаток в пище лизина приводит к нарушению кровообращения, снижению количества эритроцитов и уменьшению в них гемоглобина. Также отмечаются нарушение азотистого баланса, истощение мышц, нарушение кальцификации костей. Происходит также ряд изменений в печени и легких. Потребность в лизине составляет 3—5 г в сутки. В значительных количествах лизин содержится в твороге, мясе, рыбе.
Метионин играет важную роль в процессах метилирования и трансметилирования. Это основной донатор метильных групп, которые используются организмом для синтеза холина (витамина группы В). Метионин относится к липотропным веществам. Он оказывает влияние на обмен жиров и фосфолипидов в печени и таким образом играет важную роль в профилактике и лечении атеросклероза. Установлена связь метионина с обменом витамина В12 и фолиевой кислотой, которые стимулируют отделение метильных групп метионина, обеспечивая таким образом синтез холина в организме. Метионин имеет большое значение для функции надпочечников и необходим для синтеза адреналина. Суточная потребность в метионине составляет около 3 г. Основным источником метионина следует считать молоко и молочные продукты: в 100 г казеина содержится 3 г метионина.
Фенилаланин связан с функцией щитовидной железы и надпочечников. Он дает ядро для синтеза тироксина – основной аминокислоты, образующей белок щитовидной железы. Из фенилаланина может синтезироваться тирозин и далее адреналин. Однако обратного синтеза из тирозина-фенилаланин не происходит.
Существуют стандарты сбалансированности НАК, разработанные с учетом возрастных данных. Для взрослого человека (г/сутки): триптофана – 1, лейцина 4—6, изолейцина 3—4, валина 3—4, треонина 2—3, лизина 3—5, метионина 2—4, фенилаланина 2—4, гистидина 1,5—2.
Заменимые аминокислоты
Потребность организма в заменимых аминокислотах удовлетворяется в основном за счет эндогенного синтеза, или реутилизации. За счет реутилизации образуется 2/3 собственных белков организма. Ориентировочная суточная потребность взрослого человека в основных заменимых аминокислотах следующая (г/сутки): аргинин – 6, цистин – 2—3, тирозин – 3—4, аланин – 3, серин – 3, глутаминовая кислота – 16, аспирагиновая кислота – 6, пролин – 5, глюкокол (глицин) – 3.
Заменимые аминокислоты выполняют в организме весьма важные функции, причем некоторые из них (аргинин, цистин, тирозин, глутаминовая кислота) играют физиологическую роль не меньшую, чем незаменимые (эссенциальные) аминокислоты.
Потребность в белках зависит от возраста, пола, характера трудовой деятельности, климатических и национальных особенностей и т. д. Исследованиями установлено, что азотистое равновесие в организме взрослого человека поддерживается при поступлении не менее 55—60 г белка, однако эта величина не учитывает стрессовые ситуации, болезни, интенсивные физические нагрузки. В связи с этим в нашей стране установлена оптимальная потребность взрослого человека в белке 90—100 г/сутки. При этом в пищевом рационе за счет белка должно обеспечиваться в среднем 11—13 % общей его энергетической ценности, а в процентном отношении белок животного происхождения должен составлять не менее 55 %.
Американскими и шведскими учеными установлены ультраминимальные нормы потребления белков на основании эндогенного распада тканевых белков при безбелковых диетах: 20—25 г/сутки. Однако такие нормы при постоянном использовании не удовлетворяют потребности организма человека и не обеспечивают нормальной работоспособности, так как при распаде тканевых белков образующиеся аминокислоты, используемые в дальнейшем для ресинтеза белка, не могут обеспечить должную замену животного белка, поступающего с пищей, и это приводит к отрицательному азотистому балансу.
Энергетическая потребность людей первой группы интенсивности труда (группа умственного труда) составляет 2500 ккал. 13 % от этой величины составляет 325 ккал. Таким образом, потребность в белке у студентов составляет приблизительно 80 г (325 ккал: 4 ккал = 81,25 г) белка.
У детей потребность в белках определяется возрастными нормами. Количество белка из-за преобладания в организме пластических процессов на 1 кг массы тела увеличено. В среднем эта величина составляет 4 г/кг у детей от 1 до 3 лет жизни, 3,5 —4 г/кг для детей 3—7 лет, 3 г/кг – для детей 8—10 лет и детей старше 11 лет – 2,5—2 г/кг, в то время как в среднем у взрослых 1,2—1,5 г/кг в сутки.
Читайте также:
