
Нервная регуляция поведения животных
В этой главе мы рассмотрим нервную регуляцию поведения, начиная с изложения ее общих принципов; затем дадим обзор типов нервной системы, встречающихся у животных, и проследим, как общая организация нервной системы соотносится с поведением.
Нервные клетки
Нервная система состоит из нервных клеток, называемых нейронами, которые специализированы для передачи информации от одной клетки другой. У каждого нейрона имеется тело с ядром и множество ветвящихся отростков (рис. 11.1). Обычно у клетки много коротких отростков, называемых дендритами, и один длинный - аксон. Дендриты образуют связи с близлежащими нейронами, а аксоны проводят сигналы на сравнительно большие расстояния.
Если потенциал покоя падает ниже определенного порогового значения, вдоль мембраны распространяется потенциал действия. Он длится недолго и вызывается закономерными изменениями относительных концентраций ионов Na + и К + по обе стороны мембраны (рис. 11.3). Потенциал действия проходит к концу аксона в виде электрической волны. Он всегда имеет одну и ту же амплитуду (высоту), обычно зависящую от диаметра аксона. По более крупным аксонам распространяются более высокие потенциалы действия (и с большей скоростью), чем по более тонким аксонам.
После прохождения каждого потенциала возникает рефрактерный период, в течение которого мембрана восстанавливает свое нормальное ионное равновесие и нормальный потенциал покоя. Поскольку во время рефрактерного периода новый потенциал действия возникнуть не может, рефрактерные свойства аксона определяют максимальную частоту потенциалов действия.
 | Рис. 1 1.1. Одна нервная клеткa образует синаптический контакт с другой. |
и зависит от силы его стимуляции. Таким образом, сообщение, которое посылает нейрон, кодируется частотой (рис. 11.4).
Мембраны аксонов и дендритов не образуют физических связей с другими нейронами, а очень близко подходят к ним в соединениях. называемых синапсами. Обычно в синапсе выделяются очень малые количества химических нейромедиаторов, которые влияют на потенциал покоя воспринимающей мембраны и, следовательно, на готовность воспринимающего нейрона генерировать потенциалы действия.
Нейроны могут быть стимулированы другими нейронами, повреждением или сенсорными рецепторами. Во всех случаях принцип один и тот же. Стимуляция вызывает изменение мембранного потенциала, и когда он достигает порогового уровня, генерируется потенциал действия. Теперь рассмотрим, как этот процесс происходит в сенсорных рецепторах.
Сенсорные рецепторы
Сенсорные рецепторы - это специализированные (часто нервные) клетки, ответственные за преобразование и передачу информации. Как и обычные нервные клетки, они имеют дендриты и один или более аксонов. Рецепторы специализированы в соответствии с той энергией среды.
на которую они реагируют. Например, фоторецепторы содержат пигменты, которые химически изменяются под действием света, и при такой стимуляции возникает электрический потенциал. В механорецепторах происходят электрохимические изменения вследствие деформации мембраны клетки. Преобразование энергии обычно совершается в теле клетки, и для всех рецепторов характерно, что энергия окружающей среды превращается в градуальный электрический потенциал, называемый генераторным потенциалом, который обычно пропорционален интенсивности стимуляции рецептора. Когда генераторный потенциал достигает определенного порогового уровня, он запускает потенциал действия, который бежит по
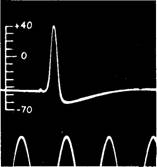 |
| Рис. 11.2. Потенциал действия гигантского аксона кальмара. (Hodgkin, Huxley, 1945.) |
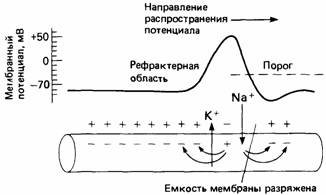 | Рис. 11.3. Распространение потенциала действия благодаря прохождению ионов К + и Na + через мембрану аксона. |
аксону рецепторной клетки. В этом заключается передаточная часть сенсорного процесса, причем информация обычно кодируется так, что, чем сильнее стимул, тем выше частота потенциалов действия. В отсутствие стимуляции генераторный потенциал постепенно снижается до уровня покоя. Когда он падает ниже порогового значения, потенциалы действия перестают генерироваться. При возобновлении стимуляции может возникнуть короткая задержка (латентный период), пока генераторный потенциал возрастает от уровня покоя до порогового. При прерывистой стимуляции он ритмически повышается и понижается, генерируя залпы потенциалов действия. Однако, если частота прерывистой стимуляции достаточно высока, генераторный потенциал может не успевать снизиться в перерывах между стимулами, и тогда генерация потенциалов действия станет непрерывной. Этим объясняется то, что при очень высокой частоте прерывистой стимуляции мы не способны отличать ее от непрерывной. Этот феномен слияния мельканий присущ всем органам чувств, что наиболее очевидно в случае зрения. Тот факт, что быстро мелькающий свет вызывает такое же зрительное ощущение, что и постоянный, делает возможным телевидение и кино.
Потенциалы действия, передающие сенсорную информацию, ничем не отличаются от любых других нервных импульсов. Их величина определяется размерами аксона, а частота - силой стимуляции. Каждый тип рецепторов посылает импульсы прямо или опосредованно в определенный отдел мозга. Испытываемые ощущения зависят не от типа рецептора или сообщений, которые он посылает, а от той части
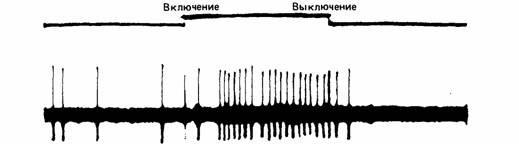 |
| Рис. 11.4. Внеклеточное отведение нервных импульсов от нейрона в зрительной системе кошки. Обратите внимание на повышение частоты импульсации при включении стимула. (Guthrie, 1980.) |
Мышцы и железы
Нервная система управляет поведением и до некоторой степени внутренней средой животного (см. гл. 15). Это управление производится приказами, отдаваемыми мышцам и железам.
В мышечных клетках имеются сложные белковые молекулы, способные к сокращению и расслаблению. Нервные окончания связаны с мышцами через синапсы, сходные с теми, какими соединены друг с другом нейроны. Придя в нервно-мышечное соединение, нервные импульсы вызывают электрические потенциалы, заставляющие мышцу сокращаться. Ее расслабление возникает при отсутствии стимуляции. Сокращаясь, мышца укорачивается, если этому не препятствует удерживание обоих ее концов. При расслаблении мышца может удлиниться, но только если ее растягивают другие мышцы или какая-нибудь внешняя сила. Мышцы обычно расположены антагонистическими, противодействующими друг другу группами. У некоторых беспозвоночных, например у кольчатых червей, мышечному сокращению может препятствовать гидростати-
 | 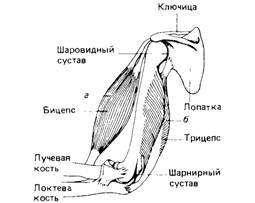 |
| Рис. 11.5. Механическое сопряжение мышц и скелета в ноге насекомого. Мышцы заключены в скелет. А. Мышца а - сгибатель (сгибает конечность), мышца б - разгибатель (выпрямляет конечность). Б. Мышцы проходят через сустав; отношения обратные: мышца а - разгибатель, мышца б - сгибатель. | Рис. 11.6. Механическое сопряжение мышц и скелета в руке человека. Мышцы лежат снаружи скелета. Мышца а - сгибатель, мышца б - разгибатель. |
ческое давление, повышающееся при сжатии мускулатурой части полости тела. Это давление заставляет мышцы удлиняться при расслаблении. У других беспозвоночных, например членистоногих, мышцы находятся внутри жесткого наружного скелета, который образует не-
обходимую систему рычагов для антагонистических групп мышц (рис. 11.5). У позвоночных животных такой системой служит внутренний скелет, а мышцы расположены так, что тянут его части в противоположные стороны (рис. 11.6). Одна группа мышц расслабляется, когда другая сокращается.
Некоторые железы находятся под нервным контролем. У позвоночных к ним относятся, например, слюнные железы, мозговая часть надпочечников, вырабатывающая адреналин, и задняя доля гипофиза, в которой образуется несколько важных гормонов. Секреты этих желез могут влиять на поведение косвенно, воздействуя на внутреннее состояние животного, как будет показано в конце этой главы.
Соместетическая система
В коже, скелетных мышцах 1 и внутренних органах позвоночных находится множество типов рецепторов. Некоторые из них показаны на рис. 11.7. Беспозвоночные также имеют широкий диапазон рецепторов. Человек обладает пятью типами кожных рецепторов, вызывающих ощущения прикосновения, давления, тепла, холода и боли. Болевых рецепторов много, в 27 раз больше, чем Холодовых, и в 270 раз больше, чем тепловых. Некоторые кожные рецепторы отличаются быстрой сенсорной адаптацией. В ответ на ступенчатое изменение стимуляции частота нервных импульсов быстро повышается, а затем снижается до уровня покоя. Это значит, что рецептор служит хорошим индикатором изменений в силе стимуляции, но плохим индикатором ее абсолютного уровня. Это дает преимущества в тех случаях, когда от кожных рецепторов требуется быстрая информация об изменениях среды, которые могут подействовать на организм, например об изменениях температуры.
Рецепторы, расположенные глубоко в теле, выполняют множество разнообразных функций, в том числе отмечают изменение кровяного давления, напряжение мышц, количество соли в крови и т. д. Мы не осознаем непосредственно информацию, передаваемую большинством интероцепторов. Они не вызывают ощущений. Иногда их действия комбинируются, давая ощущения голода, жажды или тошноты, но это происходит вследствие сложных процессов в мозгу, которые не всегда связывают ощущение с определенными частями тела. Вероятно, это объясняется тем, что действие, которое должно быть произведено в ответ на голод и жажду, является гораздо менее непосредственным, чем ответ на прикосновение или поверхностное изменение температуры.
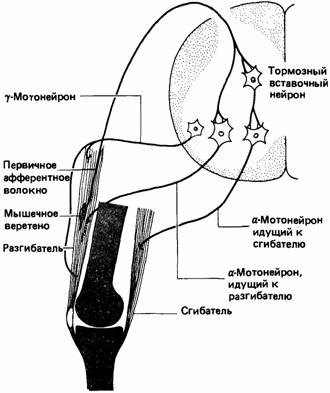
Ориентация животных по отношению к силе тяжести или внешним стимулам, подобным свету, отчасти зависит от информации о пространственном соотношении разных частей тела. У млекопитающих такая информация поступает от вестибулярной системы и рецепторов в суставах, мышцах и сухожилиях. Суставные рецепторы дают информацию об угловом положении каждого сустава (Howard, Templeton, 1966). В сухожилиях млекопитающих заложены сухожильные рецепторные органы Гольджи, чувствительные к напряжению. Они посылают сигналы в спинной мозг и участвуют в простом рефлексе, который противодействует повышению мышечного напряжения.
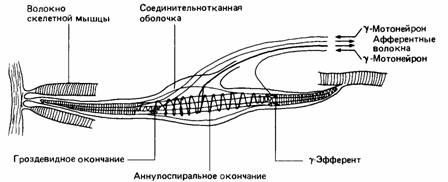
В мышцах находятся мышечные веретена, чувствительные к изменениям длины мышцы (рис. 11.8). Они состоят из видоизмененных мышечных волокон, вокруг средней части которых закручено спиральное нервное окончание, называемое
1 Рецепторы в скелетных мышцах обычно объединяют в кинестетическую систему. - Прим. перев.
 |
| Рис. 11. 7. Некоторые рецепторы, лежащие в коже, и виды чувствительности, с которой они связаны. (По Keeton, 1972.) |
первичным (или аннулоспиралъным) окончанием. При удлинении мышцы веретено растягивается и посылает быстрые сигналы в спинной мозг. Встречаются также вторичные гроздевидные окончания, посылающие более медленные сигналы. Во многих веретенах млекопитающих имеются и первичные, и вторичные окончания, а в других - только первичные (Prosser, 1973). Эти веретена участвуют в простом рефлексе, который противодействует удлинению мышцы.
Мышечные веретена заключены в фузиформную соединительную ткань, и их мышечные волокна называются интрафузальными в противоположность обычным экстрафузальным. Последние иннервируются альфа-мотонейронами, тела которых находятся в спинном мозгу. У млекопитающих интрафузальные волокна интернируются более мелкими гамма-мотонейронами, поддерживающими веретено в тоническом состоянии активности, благодаря чему для его активации требуется меньшее мышечное растяжение. Поскольку мышечные веретена расположены параллельно экстрафузальным волокнам, при сокращении мышцы они склонны к расслаблению. Гамма-нейроны могут приказать интрафузальным волокнам напрячься, в результате чего веретено сохранит состояние готовности (рис. 11.9).
У птиц мышечные веретена напоминают веретена млекопитающих и экстра- и интрафузальные волокна расположены параллельно. Но у ящерицы Tiliqua мышечные веретена, по-видимому, лежат последовательно с экстрафузальными волокнами ( Prosser , 1973). Рептилии и амфибии лишены гамма-мотонейронной системы; как интра-, так и экстрафузальные волокна связаны у них с альфа-мотонейронами. У рыб нет мышечных веретен, но в их волокнах находятся рецепторы, чувствительные к их угловой скорости.
У членистоногих имеется множество видов рецепторов растяжения; они делятся на два главных типа: 1) лежащие между элементами наружного скелета и реагирующие на вибрации в кутикуле; 2) прикрепленные к сухожилиям и сигнализирующие об изменениях растяжения и давления (Prosser, 1973). Так, крабы обладают рецепторами, которые сигнализируют о положении и движении сустава, а у падальных мух сигналы рецепторов растяжения
 |
| Рис. 11.8. Мышечное веретено млекопитающего. |
в кишке тормозят поедание пищи, когда кишка заполнена.
Большинство биологов согласится с тем, что одним из главных эволюционных направлений в животном мире является совершенствование нервной системы: Таким образом, чтобы проследить эволюцию сенсорных процессов, по-видимому, разумно ориентироваться на сложность ее устройства. Однако мы располагаем лишь
 |
| Рис. 11.9. Иннервация мышечного веретена. |
очень немногими прямыми данными о нервной системе в прошлом, так как мягкая нервная ткань редко сохраняется в окаменелом состоянии. Косвенные сведения иногда можно получить по остаткам скелета, в особенности по хорошо сохранившемуся черепу позвоночных. Большая часть наших выводов об эволюции нервной системы получена при изучении современных представителей типов животных, о которых известно, что они мало изменились за миллионы лет.
Регулирующая роль эндокринной и нервной систем в живом организме. В процессе эволюции в наиболее выгодном положении оказывались существа, имевшие совершенную систему команд, управляющих организмом. Любые частично дополняющие друг друга системы давали их носителям преимущества в жестоких доисторических условиях. В настоящее время все высшие организмы имеют дополняющие друг друга системы регуляции функций. Примером могут служить эндокринная и нервная системы, осуществляющие регуляцию основных жизненно важных функций организма.
Принцип работы нервной системы основан на преобразовании внешних раздражений в электрохимические импульсы и, далее, в ответную реакцию организма. Деятельность всех желез внутренней секреции, с самого их возникновения, не была автономной, а регулировалась центральной нервной системой по нервным проводникам, продуктами нейросекреции или посредством гормонов других эндокринных желез, выведение которых в кровь происходило в результате нервных импульсов. Поэтому говорить о самостоятельной гормональной, независимой от нервной регуляции совершенно некорректно. У всех многоклеточных животных, начиная с низших червей, регуляцию и интеграцию всех функций организма осуществляет центральная нервная система. Нервная система обеспечивает ответные реакции целостного организма на все воздействия внешней или внутренней среды, вызывающие раздражения рецепторов. Однако воздействия центральной нервной системы на эффекторы могут осуществляться двумя путями: путем передачи импульсов возбуждения по эфферентным нервам ( нервно-проводниковый путь) и путем введения в кровь или лимфу гормонов и других физиологически активных веществ ( гуморальный путь).
В настоящее время, рассматривая особенности жизнедеятельности организма животного, в том числе нервной регуляции функций, мы видим, что некоторые части нервной системы – головного мозга, обладают дополнительными функциями, выделяя гормоны – вещества, обладающие физиологической активностью и регулирующие целый ряд функций организма, например деятельность щитовидной железы, половых желез и т.д. Это наиболее эволюционно древние участки мозга, к которым относятся гипофиз и гипоталамус, одновременно входящие в состав эндокринной системы. Нервные клетки, обладающие секреторной активностью, в настоящее время обнаруживаются у всех беспозвоночных и позвоночных животных. Некоторые продукты обмена веществ нервной клетки, так называемые нейросекреты, приобрели сигнальный характер и взяли на себя регуляторные функции. При этом отдельные группы нервных клеток специализировались на выработке медиаторов нервного возбуждения.
Взаимодействие с окружающей средой и обитающими в ней организмами часто осуществляется через систему гормональной регуляции. Так, например, особые вещества, называемые феромонами, интенсивно выделяемые самками в окружающую среду в период эструса, оказывают влияние на особей своего вида, привлекая самца к самке, готовой к размножению. Самцам некоторых видов, для того чтобы почувствовать запах самки, бывает достаточно нескольких молекул феромона на один кубический метр воздуха.
Регуляторная деятельность желез внутренней секреции выражается в их взаимовлиянии, воздействии на органы-мишени, а также в антагонистическом действии некоторых гормонов на функции органов мишеней. В настоящий момент известно свыше пятидесяти подобных регуляторов, обеспечивающих нормальное функционирование организма.
Активность эндокринной системы меняется в течение жизни от этапа становления секреторной функции железы, который формируется в зависимости от железы, или в эмбриональный период (гипофиз), или позже (половые железы), через этап полноценного функционирования к постепенному угасанию.
Влияние нервной системы на функционирование желез внутренней секреции. Влияние нервной системы на функционирование желез внутренней секреции может осуществляться как опосредованно, через изменение концентрации определенных веществ, влияющих на эти железы, так и непосредственно, путем нервной регуляции. Некоторые гормоны секретируются в кровь только в результате возбуждения гормонообразующих клеток, наступающего рефлекторно в ответ на раздражение определенных рецепторов. Секреция других гормонов, напротив, систематически тормозится нервными импульсами и происходит только после прекращения их поступления из ЦНС, вызванного воздействием определенного комплекса факторов окружающей среды.
В качестве примера стимулирующего действия нервных импульсов можно привести тот факт, что механическое раздражение сосков сосущим детенышем рефлекторно вызывает поступление в кровь гормона окситоцина, выделяемого задней долей гипофиза. Окситоцин, в свою очередь, стимулирует сокращение миоэпителиальных клеток молочной железы и гладких мышечных клеток в стенках молочных протоков, что способствует молокоотдаче.
Тормозящее воздействие нервных импульсов на секрецию гормона хорошо иллюстрирует следующий пример. У самок тараканов особые эндокринные железы, так называемые прилежащие тела, вырабатывают гонадотропный гормон, стимулирующий рост ооцитов в яичниках. Однако нервные импульсы, идущие из мозга, тормозят функцию прилежащих тел и секрецию этого гормона. Погружение сперматофора в совокупительную сумку самки во время спаривания рефлекторно прекращает тормозящее действие нервных импульсов на прилежащие тела, что ведет к выведению в гемолимфу их гормона, вызывающего рост ооцитов и поступление в них желтка. Это способствует росту и развитию оплодотворенных яйцеклеток.
Влияние гормонов на высшую нервную деятельность собак изучалось сотрудниками школы И.П. Павлова.
Так, например, было установлено, что удаление щитовидной железы у щенков вызывает общую задержку развития и роста. У них практически не выражен половой тип, половой инстинкт отсутствует. Выработка условных рефлексов у таких животных затруднена и требует многократных повторений для закрепления. Очень трудно бывает выработать и дифференцировочное торможение. Серьезные нарушения поведения после тиреоидэктомии наступают и у взрослых собак. Введение гормона щитовидной железы, напротив, значительно повышает возбудимость нервных клеток коры больших полушарий.
Серьезные изменения в организме животных наступают после удаления половых желез. Как показали опыты И.П. Павлова, после кастрации самцов собак, у них наблюдается некоторое нарушение условно-рефлекторной деятельности, причем особенно сильно нарушается процесс торможения. У собак сильных типов ВНД нормальная работы коры головного мозга спустя некоторое время восстанавливается. Введение мужского полового гормону повышало величину условных рефлексов как у кастрированных, так и у интактных животных.
Глобальные повреждения высшей нервной деятельности с нарушением процессов возбуждения и торможения, а также со снижением устойчивости к сильным раздражителям вызывает у животных удаление надпочечников. Введение малых доз кортизона или дезоксикортикостерона ведет к усилению процессов возбуждения и внутреннего торможения в коре больших полушарий. Однако введение больших доз этих гормонов нарушает как положительные, так и отрицательные условные рефлексы, и развивается запредельное торможение.
Адреналин сужает кровеносные сосуды кожи и внутренних органов, за исключением сосудов головного мозга и сердца, учащает сердечные сокращения. Адреналин действует возбуждающим образом на симпатическую нервную систему и восходящую и нисходящую части ретикулярной формации. Это приводит к повышению возбудимости нервной системы; продуцируются специфические возбуждающие вещества – симпатины. Животное проявляет усиленную двигательную активность, увеличивается его агрессивность и т.д.
Роль нейрогуморальной регуляции в процессе адаптации организма к условиям окружающей среды. Параллельное развитие систем регуляции привело к образованию двух самостоятельных систем, дополняющих друг друга и способных как к экстренной, так и тонкой длительной регуляции. Обе эти системы – нервная и гуморальная, или, иначе, эндокринная, – осуществляя нейрогуморальную регуляцию, играют важную роль в процессах адаптации организма к условиям окружающей среды.
При воздействии различных экстремальных факторов, как физических (жара, холод, травма), так и психических (опасность, конфликт, радость), в организме возникает общая неспецифическая нейрогормональная реакция организма, т.е. так называемый стресс.
Факторы, вызывающие состояние стресса, Г. Селье (1974) назвал стрессорами, а совокупность изменений, происходящих в организме под воздействием стрессоров, – адаптационным синдромом. Ученые выделяют свыше двадцати видов стресса, например: эмоциональный, социальный, гипокинетический, репродуктивный, вакцинальный, лекарственный, инфекционный, пищевой, транспортировочный, гипоксический, болевой, температурный, световой, шумовой и т.д.
В развитии адаптационного синдрома выделяют четыре фазы:
Выраженность адаптационного синдрома зависит от силы воздействующих факторов и от функционального состояния многих физиологических систем, а также от характера поведения животного. Так, например, стрессором для собаки может быть испуг, перенапряжение при дрессировке, смена хозяина, часто появление новой собаки или нового члена семьи, перемена места жительства и т.д. Причем один и тот же фактор для одного животного не играет никакой роли, а для другого может оказаться сильнейшим стрессором.
В качестве стрессоров могут выступать как отрицательные, так и положительные воздействия. Без некоторого уровня стресса невозможна никакая активная деятельность. Стресс может быть не только вреден, но и полезен для организма, он мобилизует его возможности, повышает устойчивость к отрицательным воздействиям (инфекциям, кровопотере и др.), может приводить к облегчению и даже полному исчезновению многих соматических заболеваний. Вредный стресс, возникающий вследствие чрезмерной интенсивности стрессорного фактора или вследствие неадекватной реакции гормональной системы при любом, в том числе слабом, воздействии факторов, снижает сопротивляемость организма, вызывая возникновение и ухудшение течения многих заболеваний.
Важное значение для характера последствий действия стресса имеют поведенческие реакции на стрессовую ситуацию. Активный поиск способов изменения стрессовой ситуации способствует устойчивости организма и не ведет к развитию заболеваний. Отказ от активного поиска приводит к развитию фазы истощения и, в тяжелых случаях, может привести организм к гибели. Индикатором этих типов поведения и важным механизмом их регуляции является уровень катехоламинов в мозге. Таким образом, нервно-гуморальная система определяет характер реагирования организма на стресс.
При повышении плотности популяции возрастает степень конкуренции между ее членами за территорию, пищу и т.д. Увеличивается число социальных контактов, в том числе отрицательных, между животными. В результате у них происходит нарастание признаков адаптационного синдрома, увеличивается уровень кортикостероидов в крови, гипертрофируются надпочечники, разрушается иммунная система, что, в первую очередь, проявляется в процессах размножения. Увеличивается эмбриональная смертность, снижающая численность пометов, нарушается процесс лактации, что ведет к гибели подсосного молодняка. У наиболее предрасположенных к стрессу особей затормаживаются процессы сперматогенеза и онтогенеза, и они выпадают из размножения. Кроме того, в популяции увеличивается число заболеваний, являющихся прямым следствием стресса, в том числе сердечно-сосудистой и пищеварительной систем. Снижение иммунного статуса делает животных менее защищенными от инфекционных заболеваний. Если в популяции не оказывается особей, способных противостоять стрессу, то она может полностью погибнуть. За счет отбора, способствующему выживанию особей, более устойчивых к его воздействию, стресс может приводить к изменению генетической структуры популяции. Таким образом, стресс играет как отрицательную – деструктивную, так и положительную – конструктивную роль и его можно рассматривать как один из мощных механизмов естественного отбора.
Регулирующая роль эндокринной и нервной систем в живом организме
В процессе эволюции в наиболее выгодном положении оказывались существа, имевшие совершенную систему команд, управляющих организмом. Любые частично дополняющие друг друга системы давали их носителям преимущества в жестоких доисторических условиях. В настоящее время все высшие организмы имеют дополняющие друг друга системы регуляции функций. Примером могут служить эндокринная и нервная системы, осуществляющие регуляцию основных жизненно важных функций организма.
Принцип работы нервной системы основан на преобразовании внешних раздражений в электрохимические импульсы и далее в ответную реакцию организма. Деятельность всех желез внутренней секреции с самого их возникновения не была автономной, а регулировалась центральной нервной системой по нервным проводникам продуктами нейросекреции или посредством гормонов других эндокринных желез, выведение которых в кровь происходило в результате нервных импульсов. Поэтому говорить о самостоятельной гормональной регуляции, независимой от нервной, совершенно некорректно.
У всех многоклеточных животных, начиная с низших червей, регуляцию и интеграцию всех функций организма осуществляет центральная нервная система. Нервная система обеспечивает ответные реакции целостного организма на все воздействия внешней или внутренней среды, вызывающие раздражения рецепторов. Однако воздействия центральной нервной системы на эффекторы могут осуществляться двумя путями: путем передачи импульсов возбуждения по эфферентным нервам (нервно-проводниковый путь) и путем введения в кровь или лимфу гормонов и других физиологически активных веществ (гуморальный путь).
Рассматривая особенности жизнедеятельности организма животного, в том числе нервной регуляции функций, мы видим, что некоторые части нервной системы (головного мозга) обладают дополнительными функциями, выделяя гормоны – вещества, обладающие физиологической активностью и регулирующие целый ряд функций организма, например деятельность щитовидной железы, половых желез и т.д. Это наиболее эволюционно древние участки мозга, к которым относятся гипофиз и гипоталамус, одновременно входящие в состав эндокринной системы. Первные клетки, обладающие секреторной активностью, в настоящее время обнаруживаются у всех беспозвоночных и позвоночных животных. Некоторые продукты обмена веществ нервной клетки, так называемые нейросекреты, приобрели сигнальный характер и взяли на себя регуляторные функции. При этом отдельные группы нервных клеток специализировались на выработке медиаторов нервного возбуждения.
Взаимодействие с окружающей средой и обитающими в ней организмами часто осуществляется именно через систему гормональной регуляции. Так, например, особые вещества, называемые феромонами, интенсивно выделяемые самками в окружающую среду в период эструса, оказывают влияние на особей своего вида, привлекая самца к самке, готовой к размножению. Самцам некоторых видов, для того чтобы почувствовать запах самки, бывает достаточно нескольких молекул феромона на один кубический метр воздуха.
Железы внутренней секреции образованы скоплениями железистого эпителия, пронизанного большим количеством кровеносных и лимфатических сосудов, а также нервных окончаний. Выделяемые ими гормоны оказывают регулирующее влияние на определенные ткани или органы. Для осуществления действия гормона необходимо выполнение следующих условий: синтез гормона, его активация (созревание), доставка к месту "работы" и наличие в органах или тканях, на которые оказывается влияние, клеток- мишеней со специфическими рецепторами данного гормона.
Регуляторная деятельность желез внутренней секреции выражается в их взаимовлиянии, воздействии на органы-мишени, а также в антагонистическом действии некоторых гормонов на функции органов-мишеней. В настоящий момент известно свыше пятидесяти подобных регуляторов, обеспечивающих нормальное функционирование организма.
Деятельность желез внутренней секреции подвержена ритмическим колебаниям, как суточным, так и сезонным, что отражает приспособленность к существованию данного вида в определенной экологической нише. Широко известны колебания активности половых желез в связи с циклами размножения или щитовидной железы в связи с адаптацией к зимним условиям. Экспериментально показано и существование ритмов, связанных с фазами Луны. Цикличность биологических процессов обеспечивает максимальную эффективность работы организма и обеспечивается также циклическими изменениями деятельности эндокринной системы. Подобная периодичность в нарастании уровня тех или иных гормонов в крови способствует формированию в ЦНС очагов возбуждения – доминант, "запускающих" инстинктивное поведение.
Активность эндокринной системы меняется в течение жизни от этапа становления секреторной функции железы, который формируется в зависимости от железы или в эмбриональный период (гипофиз), или позже (половые железы), через этап полноценного функционирования к постепенному угасанию.
Читайте также:
