
Нервное волокно функциональное значение отдельных структурных элементов
Они обладают двумя воротами:
Будучи потенциалчувствительными, быстрые натриевые каналы обладают потенциалзависимостью и времязависимостью (рис. 1.8, А):
потенциалзависимость: активационные ворота при потенциале покоя закрыты, инактивационные — открыты; в ответ на деполяризацию активационные ворота открываются, инактивационные — закрываются;
времязависимость: активационные ворота в ответ на деполяризацию открываются чрезвычайно быстро (за десятые доли миллисекунды), инактивационные закрываются также достаточно быстро, но все же медленнее.
в состоянии покоя быстрые натриевые каналы закрыты (так как закрыты активационные ворота);
в ответ на деполяризацию быстрые натриевые каналы сначала быстро открываются (инактивационные ворота еще не закрылись, активационные уже открылись); это так называемая натриевая активация;
затем с некоторой задержкой эти каналы закрываются (так как в ответ на деполяризацию закрываются инактивационные ворота); это так называемая натриевая инактивация.
Потенциалчувствительные калиевые каналы
Они обладают только наружными (активационными) воротами.
Данные каналы также характеризуются потенциалзависимостью и времязависимостью (рис. 1.8, Б):
потенциалзависимость: ворота при потенциале покоя закрыты, а в ответ на деполяризацию открываются;
времязависимость: ворота в ответ на деполяризацию открываются сравнительно медленно (за миллисекунды).
в состоянии покоя потенциалчувствительные калиевые каналы закрыты;
при деполяризации потенциалчувствительные калиевые каналы сравнительно медленно открываются (калиевая активация);
при продолжающейся деполяризации эти каналы остаются открытыми и закрываются лишь тогда, когда мембранный потенциал возвращается к уровню покоя.
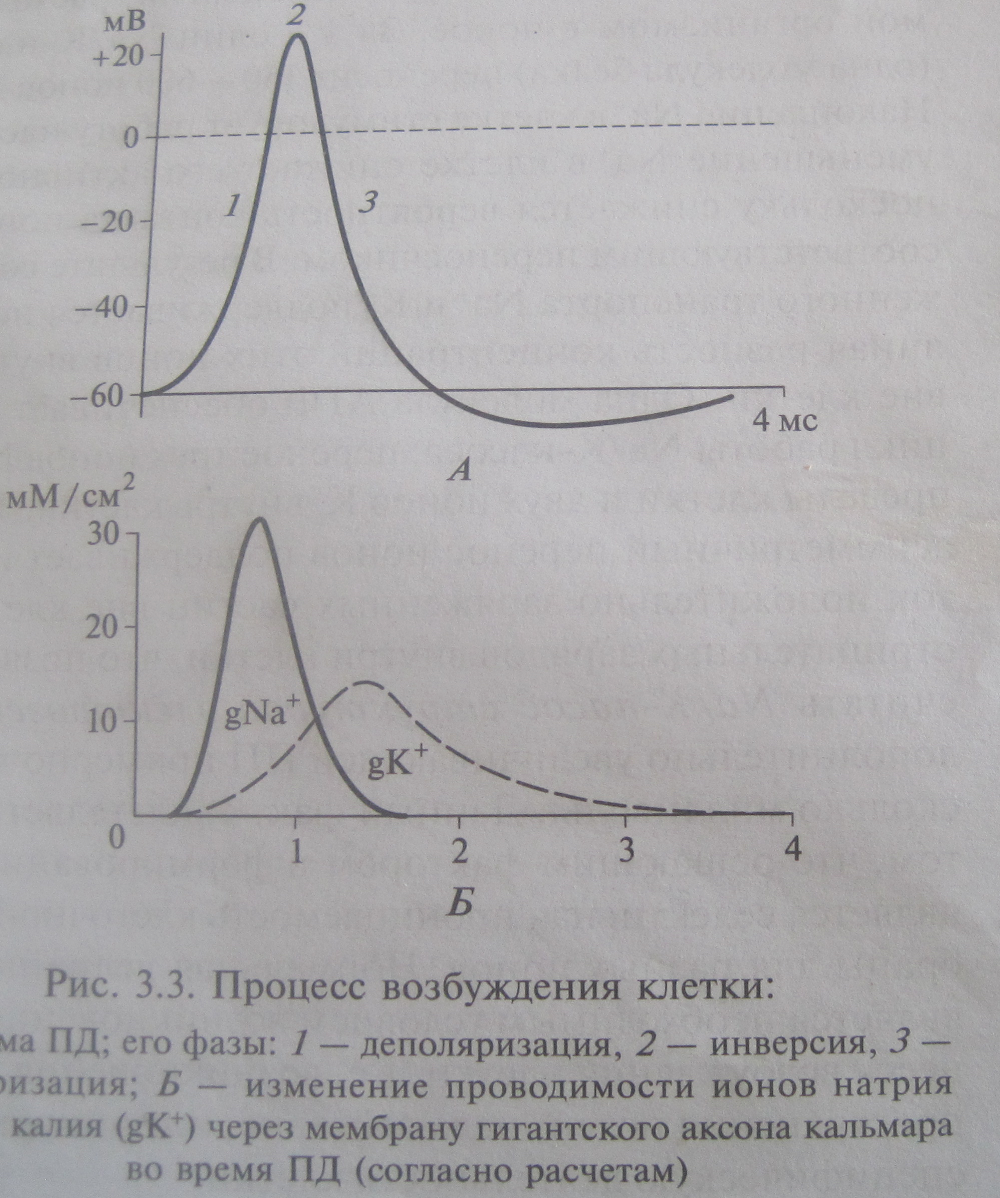
Фаза деполяризации ПД обусловлена входом в клетку Na + . Действительно, деполяризация может быть обусловлена только входом катиона, а из двух катионов, участвующих в создании биопотенциалов (Na + и K + ), только Na + стремится войти в клетку. Из этого следует, что в фазу деполяризации натриевая проницаемость должна резко повысится — то есть должны открыться быстрые натриевые каналы. Эти каналы, как уже говорилось, открываются в ответ на деполяризацию. Следовательно, деполяризация является единственным раздражителем, способным вызвать ПД.
Итак, с одной стороны, деполяризация вызывает открывание натриевых каналов, а с другой — вход Na + по этим каналам вызывает деполяризацию. Таким образом, вход Na + в фазу деполяризации ПД носит самоусиливающийся характер: деполяризация приводит к открыванию натриевых каналов; в результате входит Na + ; вход Na + приводит к еще большей деполяризации; нарастающая деполяризация вызывает открывание новых натриевых каналов и т.д.
На пике ПД движущие силы, проницаемости и потоки Na + и K + меняются следующим образом.
движущая сила (электрохимический градиент) резко падает, так как клетка становится заряженной положительно изнутри, и электрический градиент для Na + становится направленным наружу (противоположно концентрационному);
проницаемость также падает, так как закрываются инактивационные ворота быстрых натриевых каналов;
следовательно, входящий ток Na + резко снижается.
движущая сила (электрохимический градиент) резко возрастает, так как клетка становится заряженной положительно изнутри, и электрический градиент для K + становится направленным наружу (как и концентрационный);
проницаемость, будучи большой уже в покое, дополнительно возрастает, так как открываются потенциалчувствительные калиевые каналы;
следовательно, выходящий ток K + резко повышается.
Таким образом, фаза реполяризации ПД обусловлена выходом K + на фоне прекращающегося входа Na + .
По окончании ПД часто возникают так называемые следовые потенциалы, в большинстве случаев обусловленные тем, что ворота ионных каналов не успевают вернуться в состояние, характерное для потенциала покоя. Если следовой потенциал положительнее потенциала покоя, то он называется следовой деполяризацией, если отрицательнее — следовой гиперполяризацией. Эти потенциалы в разных клетках разные; в некоторых клетках их нет вовсе, в других может быть несколько сменяющих друг друга следовых потенциалов. В качестве примера на рис. 1.10 приведена следовая гиперполяризация. Видно, что она обусловлена временным повышением калиевой проницаемости (из-за медленного закрывания потенциалчувствительных калиевых каналов) и, как следствие, смещением мембранного потенциала в сторону EK.
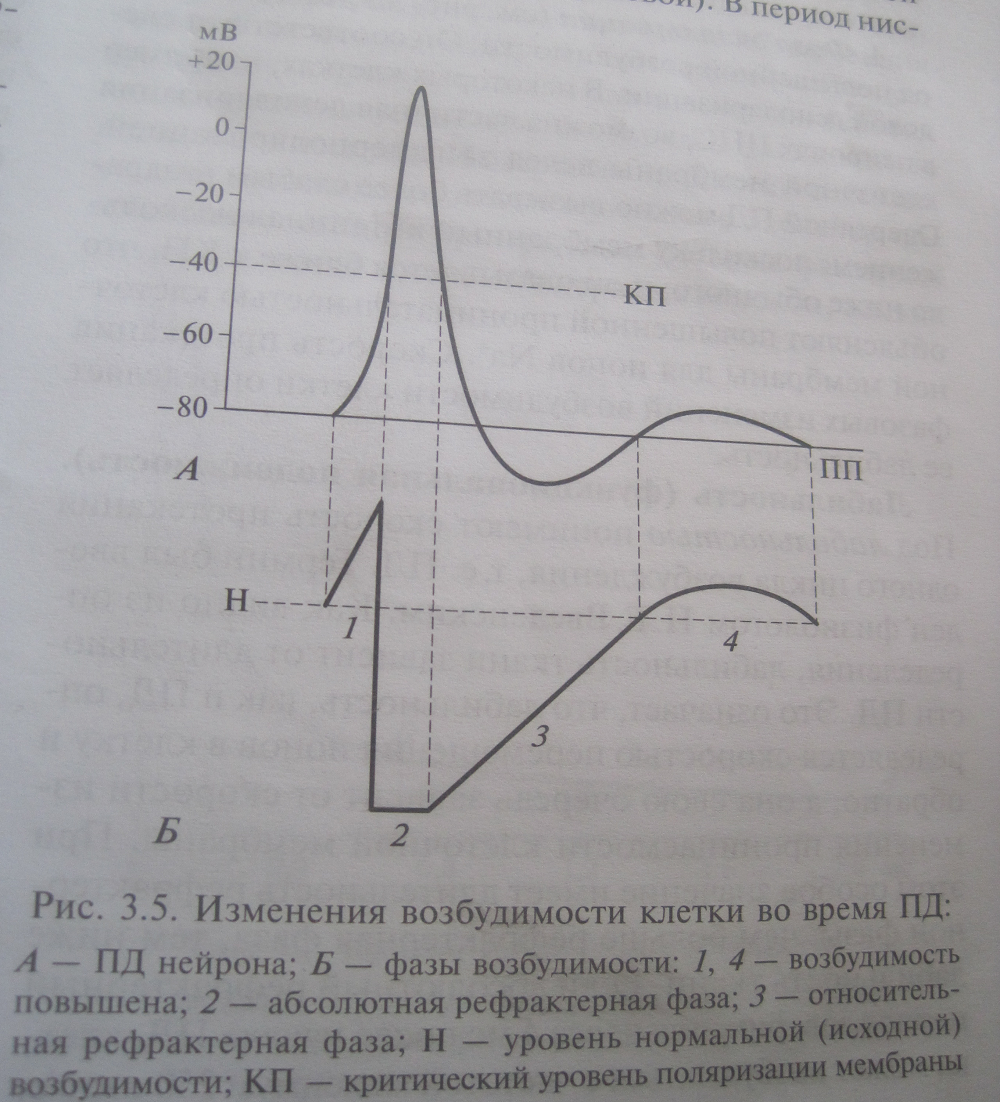
Фазовые изменения возбудимости ткани во время ее возбуждения - потенциала действия (график), их механизм. Сравнительная характеристика локального потенциала и потенциала действия. Закон “все или ничего“.
1. Во время локального ответа возбудимость повышена. Причина: мембранный потенциал приближается к Eкр.
3. Начиная от второй трети фазы реполяризации возбудимость постепенно восстанавливается, хотя и остается сниженной по сравнению с исходной (ПД может быть вызван, но только сильными раздражителями). Это период относительной невозбудимости, или относительной рефрактерности. Причина: постепенно вновь открываются инактивационные ворота быстрых натриевых каналов. Если следовых потенциалов нет, то возбудимость восстанавливается почти одновременно с окончанием фазы реполяризации (рис 1.14, А). Если же имеются следовые потенциалы, то изменения возбудимости более сложны. В частности, во время следовой гиперполяризации возбудимость понижена, поэтому период относительной рефрактерности длится дольше — до окончания следовой гиперполяризации (рис. 1.14, Б). Причины: 1) мембранный потенциал смещен в сторону более отрицательных значений; 2) калиевая проницаемость повышена, и поэтому Eкр смещен в сторону более положительных значений (рис. 1.13, В). Обе эти причины приводят к росту порога деполяризации, и, следовательно, снижению возбудимости.
Каковы бы ни были особенности изменений возбудимости в отдельных нейронах, главной и важнейшей особенностью этих изменений является наличие абсолютной рефрактерности: почти все время, пока длится ПД, повторный ПД вызвать невозможно.

Критерии для оценки возбудимости ткани. Явление аккомодации возбудимой ткани. Лабильность: понятие, мера лабильности, факторы ее определяющие, схема эксперимента для определения лабильности нерва, синапса и мышцы.
Возбудимость клетки оценивается несколькими параметрами.
Пороговый потенциал – минимальная величина, на которую надо уменьшить МПП, чтобы вызвать возбуждение в виде ПД. Небольшая величина порогового потенциала свидетельствует о высокой возбудимости клетки.
Пороговая сила – наименьшая сила раздражителя, способная вызвать возбуждение при неограничении времени ее действия. Реобаза – наименьшая сила электрического тока, способная вызвать импульсное возбуждение при неограничении времени раздражения. Большая пороговая сила свидетельствует о низкой возбудимости.
Пороговое время – минимальное время, в течение которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать ее возбуждение. Хронаксия – наименьшее время, в течение которого должен действовать электрический ток в две реобазы, чтобы вызвать возбуждение.
Аккомодация — это снижение возбудимости при уменьшении крутизны (скорости нарастания) раздражителя. Причина аккомодации — та же, что и длительной деполяризации: при медленно нарастающей деполяризации успевают развиться натриевая инактивация и калиевая активация.
Лабильность – функциональная подвижность, скорость протекания отдельных циклов возбуждения в возбудимых тканях.
Мера лабильности – максимальное количество импульсов, которое может воспроизвести ткань в единицу времени в соответствии с ритмом возбуждения.
Лабильность определяется скоростью перемещения ионов в клетку и обратно, и она в свою очередь зависит от скорости изменения проницаемости клеточной мембраны. Чем больше рефрактерная фаза, тем ниже лабильность ткани.
Нервное волокно: функциональное значение отдельных структурных элементов, классификация нервных волокон, механизм проведения возбуждения по мякотным и безмякотным волокнам, закономерности проведения возбуждения по нервному волокну и опыты их доказывающие.
Нервные волокна – отростки нейронов, с помощью которых осуществляется связь нейронов между собой, а также с иннервируемыми клетками и рецепторами.
Оболочку безмиелиновых волокон образуют швановские клетки, в которые погружаются осевые цилиндры нейронов. Оболочку миелиновых волокон образуют в периферической НС леммоциты, а в ЦНС олигодендроциты. Миелиновая оболочка влияет на электрические свойства волокна, способствует увеличению скорости проведения возбуждения, выполняет трофическую функцию (регуляция метаболизма, рост осевого цилиндра)
Плазматическая мембрана определяет пассивные электрические свойства волокна (сопротивление и емкость мембраны). Распределение и концентрация электроуправляемых каналов влияет на возбудимость и проводимость нервного волокна.
Нейрофибрилы и микротрубочки обеспечивают аксональный транспорт.
Появление разности потенциалов между возбужденными и невозбужденными участками мембраны. =>возникновение локальных токов между возбужденными и невозбужденными участками мембраны=>генерация новых ПД в соседних участках волокна.
Законы проведения возбуждения по нервному волокну
Анатомической и физиологической целостности
Изолированное проведение возбуждения
Двустороннее проведения возбуждения
Если в эксперименте нанести раздражение в любой участок нервного волокна, то возбуждение регистрируется как в проксимальном, так и в дистальном участке нерва
Относительная неутомляемость нервного волокна
Нервно-мышечный синапс: доказательства его наличия, назначение отдельных структурных элементов, механизм проведения возбуждения, особенности проведения в синапсе по сравнению с нервным волокном и опыты, их доказывающие.
Синапс – специализированная структура, обеспечивающая передачу сигнала от клетки к клетке с помощью химических веществ или электрического поля.
В состав синапса входит пре и постсинаптическая мембраны и синаптическая щель.
В области пресинаптической мембраны находятся многочисленные везикулы, которые содержат молекулы медиатора ацетилхолина. Везикулы концентрируются в особых областях- активных зонах.
Постсинаптическая мембрана образует постсинаптические складки, на гребнях которых располагаются АХ – рецепторы.
Синаптическая щель заполнена межклеточной жидкостью, в ней располагается базальная пластинка, которая содержит различные белки. Они способствуют плотному сцеплению пре и постсинаптических мембран. Базальная мембрана содержит фермент ацетилхолинэстераза, котрая разрушает высвобожденный ацетилхолин и готовит синапс к проведению очередного сигнала.
Механизм синаптической передачи.
Выброс медиатора в синаптическую щель. Он запускается посредством ПД пресинаптического окончания. Деполяризация мембраны пресинаптического окончания ведет к открытию потенциал управляемых Ca – каналов. Ca входит в нервное окончание согласно электрохимическому градиенту. Са активирует экзоцитозный аппарат пресинапса, что обеспечивает выброс ацетилхолина посредством экзоцитоза в синаптическую щель
Диффузия АХ к постсинаптической мембране, где локализуются N – холинорецепторы.
Возникновение возбуждения в мышечном волокне. В результате взаимодействия АХ с N – холинорецепторами открываются ионные каналы и вследствие преобладания входа Na в клетку происходит деполяризация постсинаптической мембраны.
Удаление АХ из синаптической щели. Осуществляется путем разрушения под действием АХЭ, около 60 % холина захватывается обратно пресинаптическим окончанием, часть АХ рассеивается.
Особенности проведения в синапсе по сравнению с нервным волокном.
Задержка в передаче сигнала к другой клетке. Это время затрачивается на высвобождение медиатора из нервного окончания, диффузию его к постсинаптической мембране и возникновение постсинаптических потенциалов, способных вызвать ПД в мышечном волокне.
Низкая лабильность синапса. Она объясняется тем, что необходимо время для высвобождения медиатора, распространения его до постсинаптической мембраны, возникновение ВПСП, а при более частом раздражении в синапсе возникают длительные ВПСП. Суммируясь они могут вызвать стойкую деполяризацию постсинаптичекой мембраны, что делает невозможным генерацию новых ПД, т.к. ПД может возникнуть только при наличии заряда в клетке.
Одностороннее проведение. Возбуждение передается только от нервного волокна к другой клетке, т.к. пресинаптическое окончание чувствительно только к нервному импульсу, а постсинаптическая мембрана к медиатору.
Неизолированное проведение. Возбуждение рядом расположенных постсинаптических мембран суммируется
Затухание возбуждения в химических синапсах принедостаточном выделении медиатора из пресинаптического окончания в синаптическую щель
Проводимость нервно – мышечного синапса угнетается и стимулируется различными веществами.
Утомляемость синапса. Главная причина – истощение запасов медиатора в пресинаптическом окончании.
7. Скелетная мышца: значение отдельных структурных элементов мышечного волокна, понятие о структурной и функциональной единице изолированной мышцы и двигательного аппарата организма, классификация двигательных единиц, физиологические свойства скелетной мышцы и ее функции
Структурной и функциональной единицей мышцы, является мышечное
волокно, представляющее собой сильно вытянутое многоядерное волокно.
Мышечное волокно имеет оболочку сарколемму. В саркоплазме мышечного
волокна имеются многочисленные ядра, митохондрии, растворимые белки,
капельки жира, гранулы гликогена, фосфатсодержащие вещества, другие
малые молекулы, а также ионы. Там же расположены многочисленные
мембраны. Они включены в состав поперечных Т – трубочек, пересекающих
мышечное волокнои связывающих сарколемму с внутриклеточным
пространством и саркоплазматическим ретикулумом. Последний
представляет собой систему связанных друг с другом систем и канальцев. Т –
система и ретикулум обеспечивают функциональное согласование процессов
возбуждения клеточной мембраны со специфической активностью
сократительного аппарата миофибриллы. Через Т – трубочки может
происходить выделение продуктов обмена в межклеточное пространство и
Внутри мышечного волокна тянется масса миофибрилл, с которыми связана
способность мышц к сокращению.
Двигательная единица – комплекс мотонейрона со всеми его коллатералями и группой иннервируемых им мышечных волокон.
Двигательные единицы подразделяют на быстрые и медленные. Медленные мотонейроны низкопороговые, малоутомляемые. Быстрые мотонейроны высокопороговые, утомляемые, не способны к длительному поддержания высокочастотного разряда.
Физиологические свойства скелетной мышцы.
1.Возбудимость. Способность мышцы отвечать на действие раздражителя самой мышцы или двигательного нерва изменением физиологических свойств и возникновением возбуждения.
2.Проводимость. Способностъ проводить возбуждение, возникшее в каком-либо участке мышечного волокна, по всему волокну.
3.Рефрактерность. Временное снижение возбудимости мышцы, которое возникает в результате возбуждения.
4.Лабильность. Количество возбуждений за единицу времени, зависящее от уровня обменных процессов.
5.Сократимость. Способность изменять свою длину или напряжение при возбуждении. Это основная функция скелетном мышцы. В период относительного покоя скелетные мышцы полностью не расслаблены, а умеренно напряжены. Такое состояние называется мышечным тонусом и объясняется редкими импульсами от двигательных нейронов, которые попеременно возбуждают нейромоторные единицы. При изотоническом сокращении укорачивается мышечное волокно, а напряжение не изменяется; при изометрическом сокращении длина мышцы не изменяется, а напряжение возрастает.
При этом мышцы выполняют следующие функции:
1) обеспечивают определенную позу тела человека;
2) перемещают тело в пространстве;
3) перемещают отдельные части тела относительно друг друга;
4) являются источником тепла, выполняя терморегуляционную функцию.
Нервное волокно - аксон, покрытый оболочками. Различают мякотные и безмякотные нервные волокна. Мякотные нервнее волокна имеют миелиновую оболочку, безмякотные ее лишены.
Мякотные волокна в отличие от безмякотных, помимо оболочки из клеток олигодендроглии, покрывающей осевой цилиндр, имеют жироподобный миелиновый слой. Последний выполняет функции изолятора - препятствует деполяризации нервного волокна. Однако слой миелина не является сплошным: на нем имеются так называемые насечки, или перехваты узла (перехват Ранвье), в которых осевой цилиндр не покрыт миелиновым слоем.
Согласно классификации Гассера-Эрлангера, нервные волокна по строению и скорости проведения возбуждения делятся на три основные группы. Различают волокна группы А - толстые миелиновые, диаметр их 15 мкм, скорость проведения возбуждения от 10 до 150 м/с; волокна группы В - тонкие миелиновые, диаметр 10 мкм, скорость проведения возбуждения до 10 м/с волокна группы С - тонкие безмиелиновые, диаметр 5 мкм, скорость проведения возбуждения до 1 м/с.
Нервное волокно обладает возбудимостью и проводимостью. Для проведения возбуждения необходима анатомическая целостность волокна. Однако не только анатомические, но и физиологические нарушения вызывают прекращение проведения. Нерв может быть целым, но он не будет проводить возбуждение, так как его функции нарушены.
Проведение возбуждения по нерву подчиняется двум основным законам.
1. Закон двустороннего проведения. Нервное волокно обладает способностью проводить возбуждение по двум направлениям: центростремительно и центробежно. Независимо от того, какое это нервное волокно - центробежное или центростремительное, если ему нанести раздражение, то возникшее возбуждение будет распространяться в обе стороны от места раздражения.
2. Закон изолированного проведения. Периферический нерв состоит из большого числа отдельных нервных волокон, которые вместе идут в одном и том же нервном стволе. В нервном стволе одновременно могут проходить самые разнообразные центробежные и центростремительные нервные волокна.
Однако возбуждение, которое передается по одному нервному волокну, не передается на соседние. Благодаря такому изолированному проведению возбуждения по нервному волокну возможны отдельные весьма тонкие движения человека.
То есть каждое волокно изолированно передает импульс мышце, и тем самым центральная нервная система имеет возможность координировать мышечные сокращения. Если бы возбуждение могло переходить на другие волокна, стало бы невозможным отдельное мышечное сокращение, каждое возбуждение сопровождалось бы сокращением самых разнообразных мышц. [4, 249c]
Основным структурным элементом нервной системы является нервная клетка, или нейрон. Через нейроны осуществляется передача информации от одного участка нервной системы к другому, обмен информацией между нервной системой и различными участками тела. В нейронах происходят сложнейшие процессы обработки информации. С их помощью формируются ответные реакции организма (рефлексы) на внешние и внутренние раздражения.
Нейроны разделяются на три основных типа: афферентные, эфферентные и промежуточные нейроны. Афферентные нейроны (чувствительные, или центростремительные) передают информацию от рецепторов в центральную нервную систему.
Афферентный нейрон имеет ложноуниполярную форму, т.е. оба его отростка выходят из одного полюса клетки. Далее нейрон разделяется на длинный дендрит, образующий на периферии воспринимающее образование - рецептор, и аксон, входящий через задние рога в спинной мозг. К афферентным нейронам относят также нервные клетки, аксоны которых составляют восходящие пути спинного и головного мозга. Эфферентные нейроны, (центробежные) связаны с передачей нисходящих влияний от вышележащих этажей нервной системы к нижележащим (например, пирамидные нейроны коры больших полушарий - рис.42) или из центральной нервной системы к рабочим органам (например, в передних рогах спинного мозга расположены тела двигательных нейронов, или мотонейронов, от которых идут волокна к скелетным мышцам; в боковых рогах спинного мозга находятся клетки вегетативной нервной системы, от которых идут пути к внутренним органам). Для эфферентных нейронов характерны разветвленная сеть дендритов и один длинный отросток - аксон. Промежуточные нейроны (интернейроны, или вставочные) - это, как правило, более мелкие клетки, осуществляющие связь между различными (в частности, афферентными и эфферентными) нейронами. Они передают нервные влияния в горизонтальном направлении (например, в пределах одного сегмента спинного мозга) и в вертикальном (например, из одного сегмента спинного мозга в другие - выше или нижележащие сегменты). Благодаря многочисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов.
Функциональное значение различных структурных элементов нейрона. Различные структурные элементы нейрона имеют свои функциональные особенности и разное физиологическое значение. Нервная клетка состоит из тела, или сомы, и различных отростков. Многочисленные древовидно разветвленные отростки дендриты служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток аксон, который передает нервные импульсы дальше - другой нервной клетке или рабочему органу (мышце, железе). Форма нервной клетки, длина и расположение отростков чрезвычайно разнообразны и зависят от функционального назначения нейрона.
Среди нейронов встречаются самые крупные клеточные элементы организма. Размеры их поперечника колеблются от 6-7 мк (мелкие зернистые клетки мозжечка) до 70 мк (моторные нейроны головного и спинного мозга). Плотность их расположения в некоторых отделах центральной нервной системы очень велика. Например, в коре больших полушарий человека на 1 мм 3 приходится почти 40 тыс. нейронов. Тела и дендриты нейронов коры занимают в целом примерно половину объема крови.
В крупных нейронах почти 1/3 - 1/4 величины их тела составляет ядро. Оно содержит довольно постоянное количество дезоксирибонуклеиновой кислоты (ДНК). Входящие в его состав ядрышки участвуют в снабжении клетки рибонуклеиновыми кислотами (РНК) и протеинами. В моторных клетках при двигательной деятельности ядрышки заметно увеличиваются в размерах. Нервная клетка покрыта плазматической мембраной - полупроницаемой клеточной оболочкой, которая обеспечивает регуляцию концентрации ионов внутри клетки и ее обмен с окружающей средой. При возбуждении проницаемость клеточной мембраны изменяется, что играет важнейшую роль в возникновении потенциала действия и передаче нервных импульсов. Аксоны многих нейронов покрыты миелиновой оболочкой, образованной Шванновскими клетками, многократно "обернутыми" вокруг ствола аксона. Однако начальная часть аксона и расширение в месте его выхода из тела клетки - аксонный холмик лишены такой оболочки. Мембрана этой немиелинизированной части нейрона - так называемого начального сегмента - обладает высокой возбудимостью.
Внутренняя часть клетки заполнена цитоплазмой, в которой расположены ядро и различные органоиды. Цитоплазма очень богата ферментными системами (в частности, обеспечивающими гликолиз) и белком. Ее пронизывает сеть трубочек и пузырьков - эндоплазматический ретикулюм. В цитоплазме имеются также отдельные зернышки - рибосомы и скопления этих зернышек - тельца Ниссля, представляющие собой белковые образования, содержащие до 50% РНК. Это белковые депо нейронов, где также происходит синтез белков и РНК. При чрезмерно длительном возбуждении нервной клетки, вирусных поражениях центральной нервной системы и других неблагоприятных воздействиях величина этих рибосомных зернышек резко уменьшается.
В специальных аппаратах нервных клеток - митохондриях совершаются окислительные процессы с образованием богатых энергией соединений (макроэргических связей АТФ). Это энергетические станции нейрона. В них происходит трансформация энергии химических связей в такую форму, которая может быть использована нервной клеткой. Митохондрии концентрируются в наиболее активных частях клетки. Их дыхательная функция усиливается при мышечной тренировке. Интенсивность окислительных процессов нарастает в нейронах более высоких отделов центральной нервной системы, особенно в коре больших полушарий. Резкие изменения митохондрий вплоть до разрушения, а следовательно, и угнетение деятельности нейронов отмечаются при различных неблагоприятных воздействиях (длительном торможении в центральной нервной системе, при интенсивном рентгеновском облучении, кислородном голодании и гипотермии). [2, 110c]
Нервные волокна являются не самостоятельными структурными элементами нервной ткани, а представляют собой комплексные образования, включающие следующие элементы:
1) отростки нервных клеток (осевые цилиндры);
2) глиальные клетки (леммоциты, или шванновские клетки);
3) соединительно-тканную пластинку (вязальную пластинку).
Главной функцией нервных волокон является проведение нервных импульсов. При этом отростки нервных клеток (осевые цилиндры) проводят нервные импульсы, а глиальные клетки (леммоциты) способствуют этому проведению.
По особенностям строения и функции нервные волокна подразделяются на две разновидности:
Строение и функциональные особенности безмиелинового нервного волокна. Безмиелиновое нервное волокно представляет собой цепь леммоцитов, в которую вдавлено несколько (5 – 20) осевых цилиндров. Каждый осевой цилиндр прогибает цитолемму леммоцита и как бы погружается в его цитоплазму. При этом осевой цилиндр окружен цитолеммой леммоцита, а ее сближенные участки составляют мезаксон.
Мезаксон в безмиелиновых нервных волокнах не играет существенной функциональной роли, но является важным структурным и функциональным образованием в миелиновом нервном волокне.
По своему строению безмиелиновые нервные волокна относятся к волокнам кабельного типа. Несмотря на это, они тонкие (5 – 7 мкм) и проводят нервные импульсы очень медленно (1 – 2 м/с).
Строение миелинового нервного волокна. Миелиновое нервное волокно имеет те же структурные компоненты, что и безмиелиновое, но отличается рядом особенностей:
1) осевой цилиндр один и погружается в центральную часть цепи леммоцита;
2) мезаксон длинный и закручен вокруг осевого цилиндра, образуя миелиновый слой;
3) цитоплазма и ядро леммоцитов сдвигаются на периферию и составляют нейролемму миелинового нервного волокна;
4) на периферии расположена базальная пластинка.
На поперечном сечении миелинового нервного волокна видны следующие структурные элементы:
1) осевой цилиндр;
2) миелиновый слой;
4) базальная пластинка.
Поскольку основу любой цитолеммы составляет билипидный слой, то миелиновую оболочку миелинового нервного волокна (закрученный мезаксон) образуют наслоения липидных слоев, интенсивно окрашивающихся в черный цвет осмиевой кислотой.
По ходу миелинового нервного волокна видны границы соседних леммоцитов – узловые перехваты (перехваты Ранвье), а также участки между двумя перехватами (межузловые сегменты), каждый из которых соответствует протяженности одного леммоцита. В каждом межузловом сегменте отчетливо прослеживаются насечки миелина – прозрачные участки, в которых содержится цитоплазма леммоцита между витками мезаксона.
Высокая скорость проведения нервных импульсов по миелиновым нервным волокнам объясняется сальтаторным способом проведения нервных импульсов: скачками от одного перехвата к другому.
Реакция нервных волокон на разрыв или пересечение. После разрыва или пересечения нервного волокна в нем осуществляются процессы дегенерации и регенерации.
Поскольку нервное волокно представляет собой совокупность нервных и глиальных клеток, то после его повреждения отмечается реакция (как в нервных, так и в глиальных клетках). После пересечения наиболее заметные изменения проявляются в дистальном отделе нервного волокна, где отмечается распад осевого цилиндра, т. е. дегенерация отсеченного от тела участка нервной клетки. Леммоциты, окружающие этот участок осевого цилиндра, не погибают, а округляются, пролиферируют и образуют тяж глиальных клеток по ходу распавшегося нервного волокна. При этом эти глиальные клетки фагоцитируют фрагменты распавшегося осевого цилиндра и его миелиновую оболочку.
В перикарионе нервной клетки с отсеченным отростком проявляются признаки раздражения: набухание ядра и сдвиг его на периферию клетки, расширение перинуклеарного пространства, дегрануляцию мембран зернистой ЭПС, вакуолизацию цитоплазмы и др.
В проксимальном отделе нервного волокна на конце осевого цилиндра образуется расширение – колба роста, которая постепенно врастает в тяж глиальных клеток на месте погибшего дистального участка этого же волокна. Глиальные клетки окружают отрастающий осевой цилиндр и постепенно трансформируются в леммоциты. В результате этих процессов происходит регенерация нервного волокна со скоростью 1 – 4 мм в сутки. Осевой цилиндр, подрастая к концевым глиоцитам распавшегося нервного окончания, разветвляется и формирует с помощью глиальных клеток концевой аппарат (двигательное или чувствительное окончание). В результате регенерации нервного волокна и нервного окончания восстанавливается иннервация нарушенного участка (реиннервация), что приводит к восстановлению его функций. Следует подчеркнуть, что необходимым условием регенерации нервного волокна является четкое сопоставление проксимального и дистального участков поврежденного нервного волокна. Это достигается сшиванием концом перерезанного нерва.
Нерв – комплексное образование, состоящее из:
1) нервных волокон;
2) рыхлой волокнистой соединительной ткани, образующей оболочки нерва.
Среди оболочек нерва различают:
1) эндоневрий (соединительную ткань, окружающую отдельные нервные волокна);
2) периневрий (соединительную ткань, окружающую пучки нервных волокон);
3) эпиневрий (соединительную ткань, окружающую нервный ствол).
В названных оболочках проходят кровеносные сосуды, обеспечивающие трофику нервных волокон.
Нервные окончания (или концевые нервные аппараты). Представляют собой окончания нервных волокон. Если осевой цилиндр нервного волокна является дендритом чувствительной нервной клетки, то его концевой аппарат образует рецептор. Если осевой цилиндр является аксоном нервной клетки, то его концевой аппарат образует эффекторное или синаптическое окончание. Следовательно, нервные окончания подразделяются на три основные группы:
1) эффекторные (двигательные или секреторные);
2) рецептурные (чувствительные);
Двигательное нервное окончание – концевой аппарат аксона на поперечно-полосатом мышечном волокне или на миоците. Двигательное нервное окончание на поперечно-полосатом мышечном волокне носит также название моторной бляшки. В нем различают три части:
1) нервный полюс;
2) синаптическую щель;
3) мышечный полюс.
В каждом терминальном ветвлении аксона содержатся следующие структурные элементы:
1) пресинаптическая мембрана;
2) синаптические пузырьки с медиатором (ацетилхолином);
3) скопление митохондрий с продольными кристами.
Мышечный полюс (или полотна моторной бляшки) включает:
1) постсинаптическую мембрану – специализированный участок плазмолеммы миосимпласта, содержащий белки-рецепторы к ацетилхолину;
2) участок саркоплазмы миосимпласта, в котором отсутствуют миофибриллы и содержится скопление ядер и саркосом.
Синаптическая щель – пространство в 50 нм между преи постсинаптическими мембранами, в котором содержится фермент ацетилхолинэстераза.
Рецепторные окончания (или рецепторы). Представляют собой специализированные концевые аппараты дендритов чувствительных нейронов, главным образом псевдоуниполярных нервных клеток спинальных ганглиев и черепных нервов, а также некоторых вегетативных нейринов (клеток II типа Догеля).
Рецепторные нервные окончания классифицируются по нескольким признакам:
1) по локализации:
а) интеророцепторы (рецепторы внутренних органов);
б) экстрорецепторы (воспринимают внешние раздражители: репетиры кожи, органов чувств);
в) проприорецепторы (локализуются в аппарате движения);
2) по специфичности восприятия (по модальности):
г) терморецепторы (тепловые, холодовые);
б) несвободные (инкапсулированные, неинкапсулированные).
РАЗДЕЛ II. ЧАСТНАЯ ГИСТОЛОГИЯ
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Читайте также:
