
Нервное волокно шванновская клетка
Шванновские клетки (иначе миелоциты или нейролеммоциты) относятся к нейроглии периферической нервной системы, где сопровождают длинные отростки нейронов в качестве вспомогательных структур. В функциональном плане они являются аналогами олигодендроцитов, присутствующих в ЦНС. Шванновские клетки находятся около аксонов, образуя оболочки периферических нервных путей.
Миелоциты впервые были охарактеризованы в 1838 году немецким физиологом Шванном, в честь которого и получили название.
Общая характеристика
Наряду с мантийными глиоцитами, леммоциты являются основными элементами периферической глии и очень похожи на сопровождающие аксоны олигодендроциты. Однако, между ними все же есть несколько отличие — в первую очередь в том, где находятся шванновские клетки. Последние сопровождают волокна ПНС, а олигодендроциты находятся в сером и белом веществе центральной нервной системы. Тем не менее, в некоторых классификациях клетки периферической глии считаются разновидностями олигодендроглии.
Отличие шванновских клеток заключается также в том, что они охватывают только один аксон, а олигодендроцит — сразу несколько. По типу формируемой оболочки нейролеммоциты бывают двух видов — миелиновые и немиелиновые, которые образуют периферические волокна соответствующих видов.
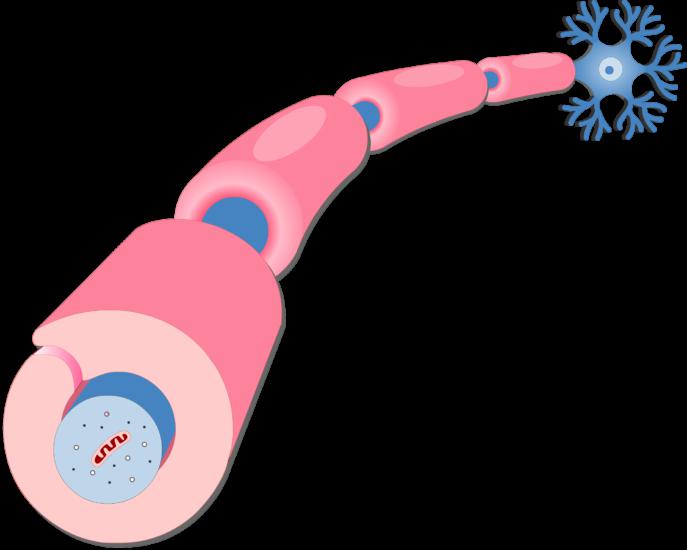
Миелоциты располагаются вдоль проводящего цилиндра. Шванновские клетки как бы оплетают волокно, образуя покрытые оболочкой сегменты, между которыми находятся перехваты Ранвье.
Особенности строения
К цитологическим особенностям леммоцитов можно отнести:
- плохо выраженный синтетический аппарат (ЭПС и пластинчатый комплекс);
- слабо развитые митохондрии;
- темноокрашенные ядра.
Длина шванновской клетки варьирует от 0,3 до 1,5 мм.
Функции
Клетки Шванна играют вспомогательную роль в поддержании работы нервного волокна. При этом они выполняют 5 основных функций:
- опорную — сеть леммоцитов формируют поддерживающую структуру для нейронов и их отростков;
- трофическую — из леммоцитов в отростки поступают различные питательные вещества;
- регенеративную — леммоциты участвуют в восстановлении поврежденных нервных волокон;
- защитную — сформированные вокруг осевых цилиндров нервные отростки обеспечивают дополнительную устойчивость к повреждениям;
- изоляционную (только для миелинизированных волокон) -- слой миелина предотвращает выход электрического сигнала за пределы конкретного нервного отростка.
Огромную роль клетки Шванна играют в восстановлении поврежденных нервных волокон. При разрыве аксона леммоциты сначала фагоцитируют поврежденные частицы, а затем размножаются и образуют мостик, соединяющий соседние концы отростка. Затем внутри этого канала вновь образуется осевой цилиндр.
Учёные из Института Солка США выявили способность тромбина разрушительно влиять на нервные клетки, а заодно и совершенно новую функцию миелинизирующих клеток периферической нервной системы. Их работа, опубликованная в журнале PLOS Genetics, показывает, что клетки Шванна блокируют действие свёртывающего белка на нейроны и препятствует их гибели. Открытие авторов может стать полезным в исследовании нейродегенеративных заболеваний периферической нервной системы. Например, бокового амиотрофического склероза.

Слева: нервно-мышечные синапсы в клетках обычного типа. В центре — то же без шванновских клеток. Справа: со шванновскими клетками и при отсутствии тромбина. Сredit: Salk Institute.
Шванновские клетки располагаются вдоль длинного отростка нейрона и создают электроизолирующий слой – миелиновую оболочку. Они выполняют опорную функцию, участвуют в питании нервной клетки и помогают передавать нервное возбуждение по аксонам. Новое исследование авторов выявляет ещё одну очень важную роль клеток Шванна: защиту аксонов от разрушения тромбином. Как оказалось, белок свертывающей системы крови может приводит нейрон к гибели.
Чтобы подтвердить влияние тромбина, исследователи рассмотрели модель соединения нейрона и мышцы с отсутствующим или не функционирующим тромбином. В результате у мышей наблюдалась меньшая дегенерация нервного волокна, что соответствует выводам учёных о разрушительном действии тромбина.
Исследование авторов открывает новый фактор влияния на жизнь нервной клетки. Теперь учёные нацелены на изучение механизма действия тромбина, а также других разрушающих ферментов. Полученные сведения будут особенно ценными в изучении проблемы нервно-мышечных заболеваний.
Текст: Екатерина Заикина
Glial cells maintain synapses by inhibiting an activity-dependent retrograde protease signal
By Thomas W. Gould, Bertha Dominguez, Fred de Winter, Gene W. Yeo, Patrick Liu, Balaji Sundararaman, Thomas Stark, Anthony Vu, Jay L. Degen, Weichun Lin, Kuo-Fen Lee
Российские ученые разработали технологию внутриутробного наблюдения за развитием миелина в мозге плода. Благодаря этому специалисты смогут сделать выводы о динамике и аномалиях структурного развития мозга…


Credit: A. Alvarez-Prats and T. Balla, Eunice Kennedy Shriver National Institute of Child Health and Human Development/NIH На этой фотографии, сделанной где-то в недрах Национальных…




Наш мозг – огромный мегаполис, дорожная инфраструктура которого напоминает связи и проводящие пути; по ним с огромной скоростью и частотой подобно спорткарам проносятся сигналы, а…

Перед вами нервный мост. Это новая ткань, образующаяся между концами разрезанного нерва. Она состоит из макрофагов и мигрирующих шванновских клеток, в норме образующих миелиновую изоляцию…
К периферическим нервам относят черепные и спинномозговые нервы, соединяющие центральную нервную систему (ЦНС) с периферическими органами и тканями. Спинномозговые нервы формируются при слиянии вентральных (передних) и дорсальных (задних) нервных корешков в месте их выхода из позвоночного канала. Задние нервные корешки образуют утолщения — спинальные ганглии (или задние корешковые ганглии). Спинномозговые нервы относительно короткие — их длина составляет менее 1 см. Проходя через межпозвоночное отверстие, спинномозговые нервы делятся на вентральную (переднюю) и дорсальную (заднюю) ветви.
Задняя ветвь обеспечивает иннервацию мышц, выпрямляющих позвоночник, а также кожи туловища в этой области. Передняя ветвь иннервирует мышцы и кожу передней части туловища; кроме того, от нее отходят чувствительные волокна к париетальной плевре и париетальной брюшине.
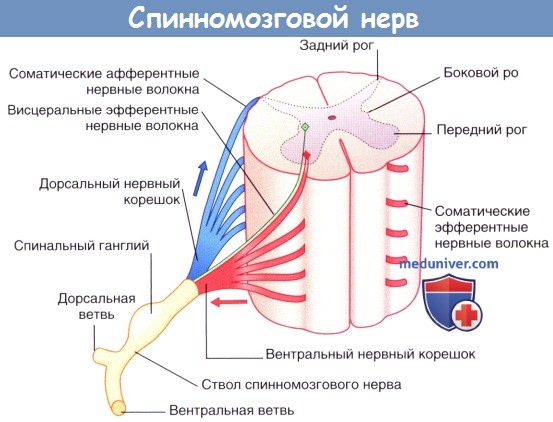
Грудной сегмент спинного мозга и нервные корешки.
Стрелками указано направление проведения импульса. Зеленым цветом обозначено симпатическое нервное волокно.
В состав спинномозгового нерва входят соматические эфферентные нервные волокна, направляющиеся к скелетной мускулатуре туловища и конечностей, и соматические афферентные нервные волокна, проводящие возбуждение от кожи, мышц и суставов. Кроме того, в спинномозговом нерве расположены висцеральные эфферентные и, в некоторых случаях, афферентные вегетативные нервные волокна.
Общие принципы внутреннего строения периферических нервов схематично изображены на рисунке ниже. Только лишь по строению нервных волокон невозможно определить, являются они двигательными или чувствительными.
Периферические нервы окружены эпиневрием — внешним слоем, состоящим из плотной неравномерной соединительной ткани и располагающимся вокруг пучков нервных волокон и сосудов, кровоснабжающих нерв. Нервные волокна периферических нервов могут переходить из одного пучка в другой.
Каждый пучок нервных волокон покрыт периневрием, представленным несколькими отчетливо различимыми эпителиальными слоями, связанными плотными щелевидными соединениями. Отдельные шванновские клетки окружены эндоневрием, образованным ретикулярными коллагеновыми волокнами.
Менее половины нервных волокон покрыто миелиновой оболочкой. Немиелинизированные нервные волокна расположены в глубоких складках шванновских клеток.
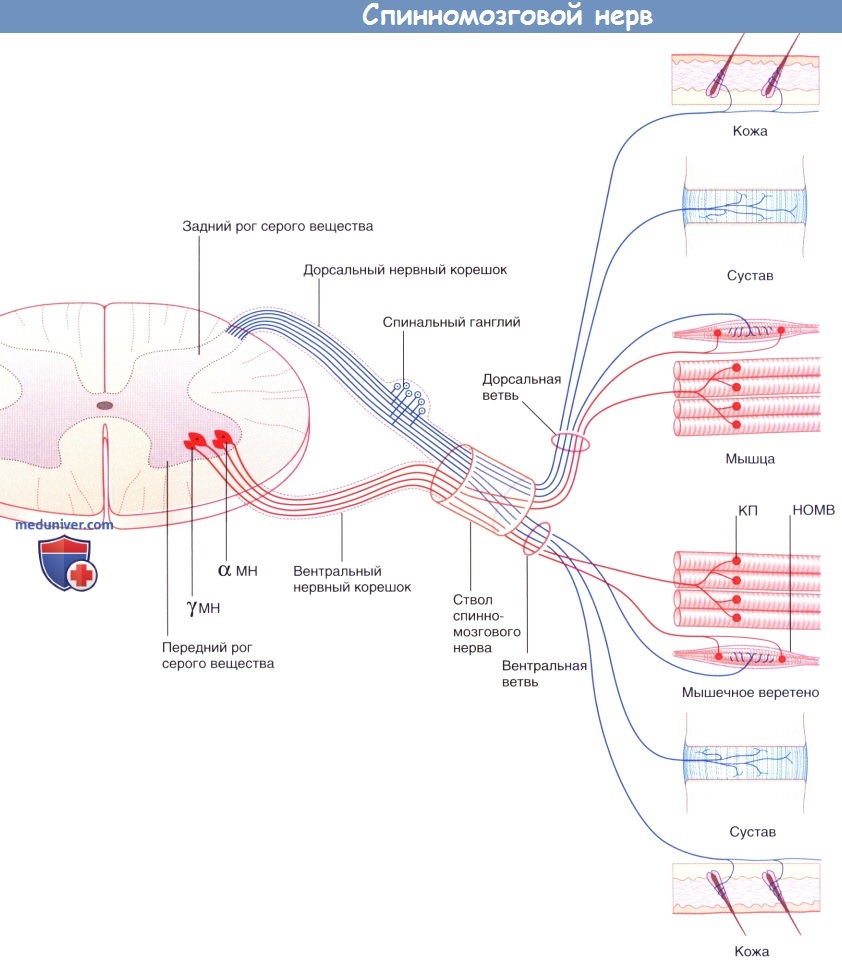
Строение грудного спинномозгового нерва. Обратите внимание: на рисунке не указан симпатический компонент.
КП — концевая пластинка двигательного нерва на мышце; НОМВ — нервное окончание мышечного веретена; МН — мультиполярный нейрон.
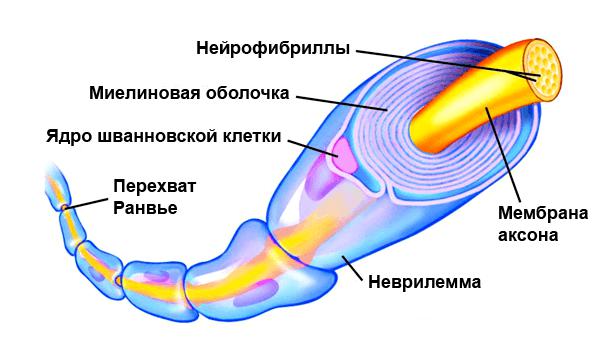
а) Образование миелина. Шванновские клетки (леммоциты) — представители нейроглиальных клеток периферической нервной системы. Эти клетки образуют непрерывную цепочку вдоль периферических нервных волокон. Каждая шванновская клетка миелинизирует участок нервного волокна длиной от 0,3 до 1 мм. Видоизменяясь, шванновские клетки образуют в спинальных и вегетативных ганглиях сателлитные глиоциты, а в области нервно-мышечных соединений — клетки телоглии.
В области конечных участков миелинизированных сегментов аксона по обеим сторонам от перехватов Ранвье (промежутков между конечными участками соседних шванновских клеток) расположены паранодальные карманы.
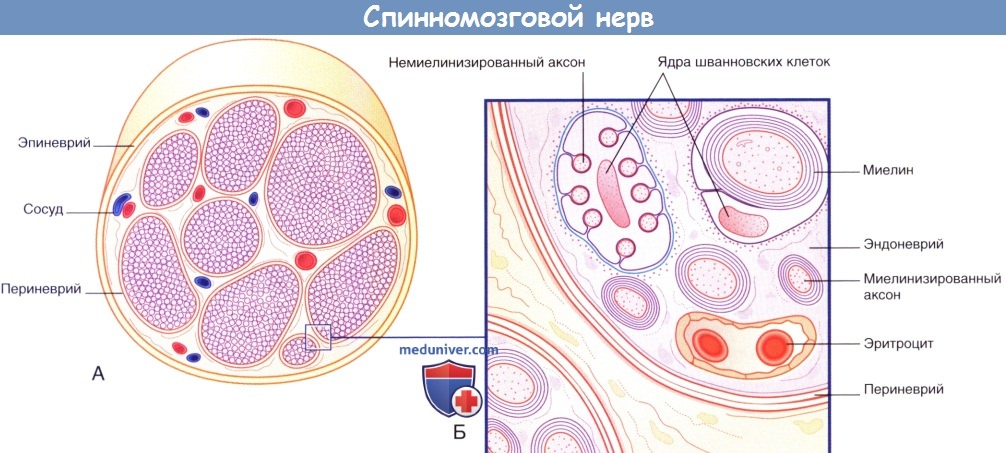
Поперечный срез нервного ствола.
(А) Световая микроскопия. (Б) Электронная микроскопия. 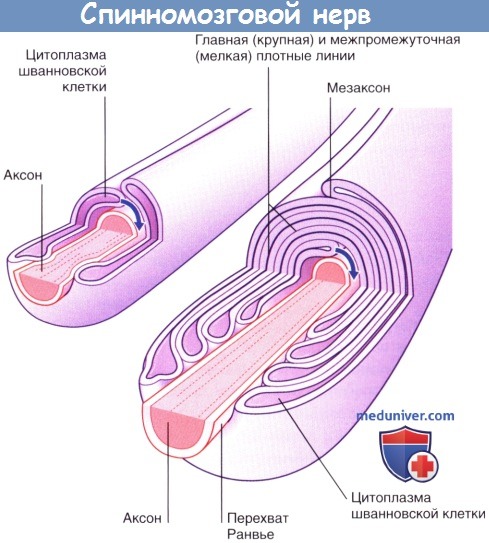
Миелинизация в периферической нервной системе.
Стрелками указано направление накручивания цитоплазмы шванновской клетки.
С точки зрения физиологии периферические нервные волокна классифицируют по скорости проведения нервных импульсов, а также по другим критериям. Двигательные нервные волокна разделяют на типы А, В и С в соответствии с уменьшением скорости проведения импульсов. Чувствительные нервные волокна разделяют на группы I—IV по такому же принципу. Однако на практике эти классификации взаимозаменяемы: так, например, немиелинизированные чувствительные нервные волокна относят не к типу С, а к группе IV.
Подробная информация о диаметрах и местах локализации периферических нервных волокон представлена в таблицах ниже.

На электронно-микроскопическом изображении показаны миелинизированное периферическое нервное волокно и окружающая его шванновская клетка. На рисунках ниже представлена группа немиелинизированных нервных волокон, погруженных в цитоплазму шванновской клетки и продемонстрирован участок перехвата Ранвье аксона ЦНС.
в) Резюме. Стволы спинномозговых нервов проходят в межпозвоночных отверстиях. Эти структуры образуются при соединении вентральных (двигательных) и дорсальных (чувствительных) нервных корешков и разделяются на смешанные вентральные и дорсальные ветви. Нервные сплетения конечностей представлены вентральными ветвями.
Периферические нервы покрыты эпиневральной соединительной тканью, пучковидной периневральной оболочкой и эндоневрием, образованным коллагеновыми волокнами и содержащим шванновские клетки. Миелинизированное нервное волокно включает аксон, миелиновую оболочку и цитоплазму шванновской клетки — нейролемму. Миелиновые оболочки формируются шванновскими клетками и обеспечивают сальтаторное проведение импульсов со скоростью, прямо пропорциональной диаметру нервного волокна.

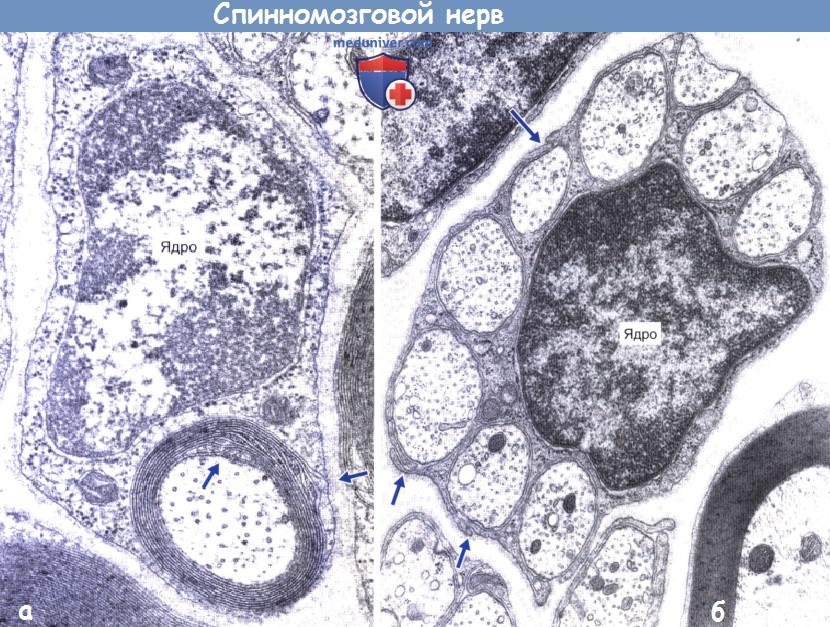
а - Миелинизированное нервное волокно. Десять слоев миелина окружают аксон от внешнего к внутреннему мезаксону шванновской клетки (указано стрелками). Базальная мембрана окружает шванновскую клетку.
б - Немиелинизированные нервные волокна. Девять немиелинизированных волокон погружены в цитоплазму шванновской клетки. Мезаксоны (некоторые указаны стрелками) визуализируются при полном погружении аксонов.
Два неполностью погруженных аксона (сверху справа) покрыты базальной мембраной шванновской клетки. 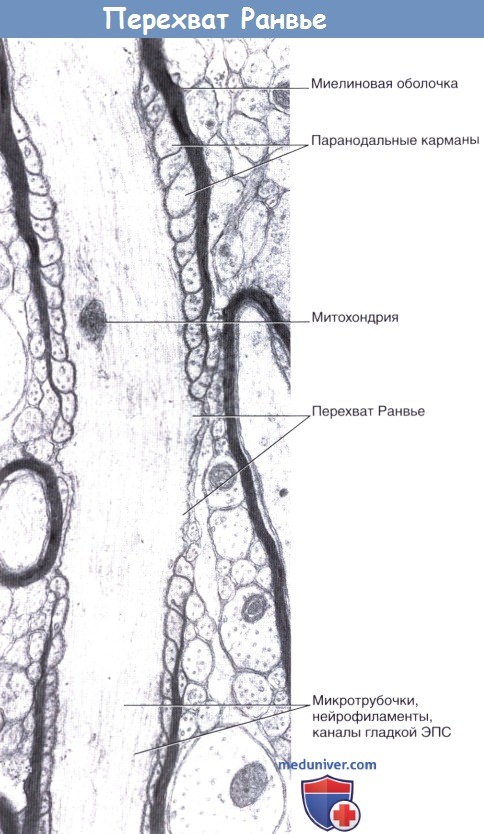
Область перехвата Ранвье ЦНС. Доходя до области перехвата Ранвье, миелиновая оболочка сужается и заканчивается, закручиваясь в области паранодальных карманов цитоплазмы олигодендроцита.
Длина области перехвата Ранвье составляет около 10 нм; на этом участке отсутствует базальная мембрана.
Микротрубочки, нейрофиламенты и удлиненные цистерны гладкой эндоплазматической сети (ЭПС) формируют продольные пучки. 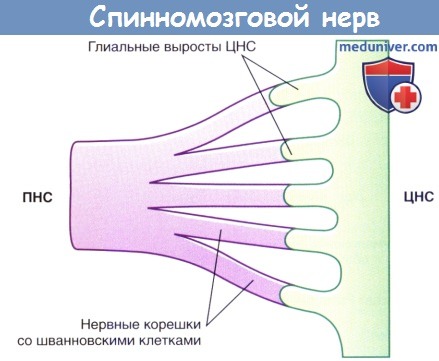
Область перехода центральной нервной системы (ЦНС) в периферическую нервную систему (ПНС).
Редактор: Искандер Милевски. Дата публикации: 12.11.2018
Состоят из отростка нервной клетки, покрытого оболочкой, которая формируется олигодендроцитами. Отросток нервной клетки (аксон или дендрит) в составе нервного волокна называется осевым цилиндром.
- безмиелиновое (безмякотное) нервное волокно,
- миелиновое (мякотное) нервное волокно.
Безмиелиновые нервные волокна
Находятся преимущественно в составе вегетативной нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи, в которых на определенном расстоянии друг от друга видны овальные ядра. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько (10—20) осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в смежное. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. При электронной микроскопии безмиелиновых нервных волокон видно, что по мере погружения осевых цилиндров в тяж неиролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне
29. Миелиновые нервные волокна
1) внутренний, более толстый, — миелиновый слой,
2) наружный, тонкий, состоящий из цитоплазмы, ядер нейролеммоцитов и нейролеммы.
Миелиновый слой содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии — насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы видны участки волокна, лишенные миелинового слоя, — узловатые перехваты, или перехваты Ранвье, т.е. границы между соседними леммоцитами.
Отрезок волокна между смежными перехватами называется межузловым сегментом.
В процессе развития аксон погружается в желобок на поверхности нейролеммоцита. Края желобка смыкаются. При этом образуется двойная складка плазмолеммы нейролеммоцита — мезаксон. Мезаксон удлиняется, концентрически наслаивается на осевой цилиндр и образует вокруг него плотную слоистую зону — миелиновый слой. Цитоплазма с ядрами отодвигается на периферию – образуется наружная оболочка или светлая Шванновская оболочка (при окраске осмиевой кислотой).
Осевой цилиндр состоит из нейроплазмы, продольных параллельных нейрофиламентов, митохондрий. С поверхности покрыт мембраной – аксолеммой, обеспечивающей проведение нервного импульса. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Нервный импульс в миелиновом нервном волокне проводится как волна деполяризации цитолеммы осевого цилиндра, "прыгающая" (сальтирующая) от перехвата к следующему перехвату со скоростью до 120 м/сек.
В случае повреждения только отростка нейроцита регенерация возможна и протекает успешно при наличии определенных для этого условий. При этом, дистальнее места повреждения осевой цилиндр нервного волокна подвергается деструкции и рассасывается, но леммоциты при этом остаются жизнеспособными. Свободный конец осевого цилиндра выше места повреждения утолщается - образуется "колба роста", и начинает расти со скоростью 1 мм/день вдоль оставшихся в живых леммоцитов поврежденного нервного волокна, т.е. эти леммоциты играют роль "проводника" для растущего осевого цилиндра. При благоприятных условиях растущий осевой цилиндр достигает бывшего рецепторного или эффекторного концевого аппарата и формирует новый концевой аппарат.
30. шванновские клетки (леммоциты) — вспомогательные клетки нервной ткани, которые формируются вдоль аксонов периферических нервных волокон. Создают, а иногда и разрушают, электроизолирующую миелиновую оболочку нейронов. Выполняют опорную (поддерживают аксон) и трофическую (питают тело нейрона) функции. Описаны немецким физиологом Теодором Шванном в 1838 году и названы в его честь.
Каждое периферическое нервное волокно одето тонким цитоплазматическим слоем — невролеммой или шванновской оболочкой. Волокно является миелинизированным, если между ним и цитоплазмой шванновской клетки имеется значительный слой миелина. Если волокна лишены миелина, то они называются немиелинизированными безмякотными. Шванновские клетки могут осуществлять волнообразные движения, что, вероятно, способствует транспортировке различных веществ по отросткам нервных клеток.
С нарушением работы шванновских клеток связаны такие нервные заболевания, как синдром Гийена — Барре, болезнь Шарко-Мари, шванноматозис и хроническая воспалительная демиелинизирующая полинейропатия. Демиелинизация в основном происходит из-за ослабления двигательных функций шванновских клеток, в результате чего они оказываются не способны образовывать миелиновую оболочку.
Особенности распространения возбуждения по безмиелиновым волокнам:
1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением.
2. Возбуждение распространяется с небольшой скоростью.
3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна).
По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров.
Однако низкая скорость распространения возбуждения и его затухание не всегда выгодно организму. Поэтому природой был выработан еще один дополнительный механизм распространения возбуждения.
32. Механизмы проведения возбуждения в миелиновых волокнах. Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. В миелинизированномволокне токи проводятся только в зонах, не покрытых миелином (перехватах Ранвье). В этих участках генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением.
При действии раздражителя пороговой силы на мембрану миелинового волокна в области перехвата Ранвье изменяется проницаемость для ионов Nа + , которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны, что ведет к возникновению круговых токов. Этот ток идет через межтканевую жидкость к соседнему перехвату, где происходит смена заряда. Таким образом, возбуждение перепрыгивает с одного участка на другой. Обратное движение возбуждения невозможно так как участок, через который оно прошло, находится в фазе абсолютной рефрактерности.
Особенности распространения возбуждения по миелиновым волокнам:
2. Возбуждение распространяется с большой скоростью.
3. Возбуждение распространяется без декримента.
По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции.
Сальтаторное проведение
(лат. saltatorius, от salto — скачу, прыгаю)
Сальтаторное распространение возбуждения в миелинизированном нервном волокне от перехвата к перехвату [стрелками показано направление тока, возникающего между возбуждённым (А) и соседним покоящимся (Б) перехватом].
34. Проведение нервного импульса, передача сигнала в виде волны возбуждения в пределах одного нейрона и от одной клетки к другой. П. н. и. по нервным проводникам происходит с помощью электротонических потенциалов и потенциалов действия, которые распространяются вдоль волокна в обоих направлениях, не переходя на соседние волокна (см. Биоэлектрические потенциалы, Импульс нервный). Передача межклеточных сигналов осуществляется через синапсы чаще всего с помощью медиаторов, вызывающих появление потенциалов постсинаптических. Нервные проводники можно рассматривать как кабели, обладающие относительно низким осевым сопротивлением (сопротивление аксоплазмы — ri) и более высоким сопротивлением оболочки (сопротивление мембраны — rm). Нервный импульс распространяетсявдоль нервного проводника посредством прохождения тока между покоящимися и активными участками нерва (локальные токи). В проводнике по мере увеличения расстояния от места возникновения возбуждения происходит постепенное, а в случае однородной структуры проводника экспоненциальное затухание импульса, который в 2,7 раза уменьшается на расстоянии l = (константа длины). Так как rm и riнаходятся в обратном отношении к диаметру проводника, то затухание нервного импульса в тонких волокнах происходит раньше, чем в толстых. Несовершенство кабельных свойств нервных проводников восполняется тем, что они обладают возбудимостью. Основное условие возбуждения — наличие у нервов потенциала покоя. Если локальный ток через покоящийся участок вызовет деполяризацию мембраны, достигающую критического уровня (порога), это приведёт к возникновению распространяющегося потенциала действия (ПД). Соотношение уровня пороговой деполяризации и амплитуды ПД, обычно составляющее не менее 1: 5, обеспечивает высокую надёжность проведения: участки проводника, обладающие способностью генерировать ПД, могут отстоять друг от друга на таком расстоянии, преодолевая которое нервный импульс снижает свою амплитуду почти в 5 раз. Этот ослабленный сигнал будет снова усилен до стандартного уровня (амплитуда ПД) и сможет продолжить свой путь по нерву.
Нервные волокна – это отростки нейронов, окруженные оболочками. Различают два вида нервных волокон:
Безмиелиновыенервные волокна имеют следующие компоненты:
− осевые цилиндры (отростки нейронов). Их в составе одного волокна несколько,
− шванновская оболочка, которая образована глиальными (шванновскими) клетками.
Под электронным микроскопом можно заметить, что осевые цилиндры располагаются в складках цитолеммы шванновских клеток, т.е. вне клеток. В связи с этим следует знать термин – мезаксон. Это сближенные края цитолеммы шванновских клеток при образовании складок. Место расположения безмиелиновых нервных волокон – это постганглионарные волокна вегетативной нервной системы. Скорость проведения нервных импульсов по безмиелиновым нервным волокнам – от 0,5 до 2 м/сек.
Миелиновые нервные волокна
Под световым микроскопом миелиновые нервные волокна представляют из себя однородные тяжи, которые на определенном расстоянии друг от друга имеют сужения (перехваты Ранвье) (Рис. 17).
Компоненты миелинового нервного волокна:
− миелиновая оболочка и
Осевой цилиндр (отросток нейрона) в составе миелинового нервного волокна – один. Как указывалось ранее, он имеет оболочку (аксолемма) и цитоплазм (аксоплазма).
Миелиновая оболочка. Чтобы понять строение миелиновой оболочки следует проследить развитие миелиновой оболочки. Вначале осевой цилиндр погружается в складку цитолеммы шванновских клеток, которые затем начинают закручиваться вокруг осевого цилиндра. При этом мезаксон (сдвоенная цитолемма шванновских клеток) ложится в виде слоев вокруг осевого цилиндра, образуя миелиновую оболочку. Таким образом, миелиновая оболочка – это наслоенные друг на друга листки мезаксона, т.е. наслоенные друг на друга липопротеидные мембраны.

- Осевой цилиндр.
- Миелиновый слой
б. Перехват Ранвье
в. Насечка миелина
Шванновская оболочка. Это наружная оболочка нервного волокна, которая содержит цитоплазму и ядра шванновских клеток.
Перехват Ранвье это место стыка (соединения) двух соседних шванновских клеток. Эти участки имеют особенности строения:
− в них отсутствует миелиновая оболочка; осевой цилиндр здесь покрыт только шванновской оболочкой,
− в этом месте отмечается скопление митохондрий.
Перехваты Ранвье – это функционально активные участки миелинового нервного волокна, а именно, передача нервных импульсов в нем совершается скачкообразно, от одного перехвата до другого.
Скорость проведения нервных импульсов в миелиновых нервных волокнах – от 3 до 100 м/сек.
Миелиновые нервные волокна – это основной вид волокон. Все нервные волокна в составе центральной и периферической нервной системы (за исключением постганглионарных волокон вегетативной нервной системы) по строению миелиновые.
Периферические нервы
Состоят из миелиновых (в основном) нервных волокон. Миелиновые нервные волокна, кроме миелиновой и шванновской, имеют оболочку – неврилемма, которая имеет в своем составе: базальную мембрану и сеть коллагеновых волокон. Между отдельными нервными волокнами прослойки рыхлой соединительной ткани – это эндоневрий. Соединительнотканные прослойки между пучками нервных волокон – периневрий и общая соединительнотканная оболочка нервного ствола – эпиневрий.
Нервные окончания
− чувствительные нервные окончания,
− двигательные нервные окончания и
Чувствительные нервные окончания (рецепторы). Определения:
1. Рецептор – это специализированный нервный аппарат, воспринимающий действия раздражителей,
2. (по И.П. Павлову), рецепторы – это трансформаторы энергии раздражителей в нервные импульсы.
Физиологическая классификация рецепторов:
− механорецепторы – стимулируются механическими движениями органов или их частей,
− барорецепторы – воспринимают изменения давления,
− хеморецепторы – стимулируются различными химическими веществами,
− терморецепторы – воспринимают колебания температуры.
Морфологическая классификация чувствительных нервных окончаний. По строению их делят на: свободные и несвободные. Несвободные в свою очередь делят на: неинкапсулированные и инкапсулированные.
Свободные нервные окончания – это голые ветвления осевого цилиндра, т.е. они (ветвления) свободны от оболочек (миелиновой и шванновской) (Рис. 18). Отличаются большой чувствительностью. Их много в составе эпителия, особенно в составе разновидностей многослойного эпителия. Пример. Роговица глаза отличается высокой чувствительностью, из-за наличия между клетками эпителия большого количества свободных чувствительных нервных окончаний.

- Слой шиповатых клеток
- Рыхлая соединительная ткань
- Терминали
- Нервные волокна
Несвободные нервные окончания. В них концевые ветвления осевого цилиндра покрыты оболочками.
Несвободные неинкапсулированныенервные окончания – когда концевые ветвления осевого цилиндра покрыты оболочкой из клеток нейроглии. Таких рецепторов много в составе эпителия и соединительных тканей.
Несвободные инкапсулированные нервные окончания. Их разновидности: колбы Краузе, тельца Мейснера, тельца Фатер – Пачини. По строению они сходны. Рассмотрим их строение на примере телец Фатер – Пачини (Рис. 19). В средне части тельца располагается концевое ветвление осевого цилиндра. Оно окружено видоизмененными глиальными клетками, образующими внутреннюю капсулу тельце. Внутренняя капсула окружена наружной капсулой, состоящей из наслоенных друг на друга соединительнотканных пластинок. Тельца Фатер – Пачини воспринимают давление на орган, т.е. функционально механорецепторы.

1.Секреторные отделы железы
2, 3. Пластинка соединительной ткани
б. Осевой цилиндр
4. Миелиновое нервное волокно.
Двигательные нервные окончания передают нервные импульсы на рабочие органы и ткани. В скелетных мышцах они называются нервно-мышечные окончания.
Синапс. Это место контакта двух нейронов. В зависимости от того, какие части двух нейронов контактируют, различают следующие виды синапсов:
− аксо-соматические, когда аксон первого нейрона контактирует с телом второго,
− аксо-дендритические, когда аксон первого нейрона контактирует с дендритов второго,
− аксо-аксональные, когда аксон первого нейрона контактирует с аксоном второго.
Синапсы имеют две части:
− пресинаптический, полюс и
Пресинаптический полюс – это концевое расширение аксона первого нейрона; содержит скопления митохондрий и пресинаптические пузырьки, имеющие в своем составе медиатор (ацетилхолин или норадреналин) и пресинаптическую мембрану (часть аксолеммы концевого расширения).
Постсинаптический полюс – это прилежащая часть второго нейрона. Имеет постсинаптическую мембрану.
Между пре- и постсинаптической мембраной располагается синаптическая щель. Основное свойство синапса – одностороннее проведение нервных импульсов от пресинаптического полюса в постсинаптический.
Читайте также:
