
Нервные центры принципы взаимодействия
Иррадиация возбуждения. При сильном и длительном раздражении импульсы, поступившие в нервную систему, не ограничиваются возбуждением одного рефлекторного центра, а распространяются на другие центры. Чем сильнее раздражение, тем больше нервных центров охватывает иррадиация. Возбуждение большого количества различных нервных центров позволяет отобрать из них наиболее нужные и установить между ними новые функциональные связи — условные рефлексы. Большинство двигательных условных рефлексов формируется благодаря иррадиации возбуждения. Излишняя иррадиация возбуждения центральной нервной системы нарушает уравновешенность поведенческих реакций, приводит к расстройству рефлекторной деятельности. Иррадиация возбуждения ограничивается и уравновешивается торможением.
Основными принципами координации являются:
- Принцип сопряженного торможения или реципрокности - базируется на описанном выше одноименном виде торможения и означает возникновение при возбуждении одного центра одновременного торможения другого центра для обеспечения цепных рефлексов и сложной функции. Примером реципрокности может быть регуляция спинным мозгом противоположных по функциональному назначению мышц конечностей. Так, при возбуждении мотонейронов, иннервирующих мышцы сгибатели правой ноги, реципрокно тормозятся мотонейроны мышц разгибателей этой ноги и возбуждаются мотонейроны мышц разгибателей левой ноги. Формирующийся цепной характер рефлексов вслед за этим вызывает возбуждение мотонейронов разгибателей правой ноги и реципрокно — торможение мотонейронов сгибателей правой ноги и возбуждение мотонейронои сгибателей левой ноги. Таким образом, реципрокные взаимоотношения между указанными рефлексами обеспечивают цепной шагательный рефлекс. Реципрокные взаимоотношения имеют место и между рефлексами вдоха и выдоха, когда возбуждение центра вдоха тормозит центр выдоха и наоборот, что обеспечивает ритмичную смену фаз в процессе внешнего дыхания.
- Принцип общего конечного пути - описанный Ч.Шеррингтоном, рассматривался при изложении процесса конвергенции возбуждения.
- Принцип субординации нервных центров - (принцип подчинения) проявляется в виде регулирующего влияния выше расположенных нервных центров на ниже расположенные. Так, двигательные центры головного мозга управляют спинальными мотонейронами. Примером такого влияния является феномен центрального торможения спинальных рефлексов открытый И.М.Сеченовым и получивший название сеченовского торможения. В эксперименте И.М.Сеченова раздражение зрительных бугров лягушки с помощью кристаллика поваренной соли (т.е. раздражение ретикулярной формации среднего мозга) приводило к торможению спинальных двигательных рефлексов, вызываемых погружением лапки лягушки в слабый раствор кислоты. Следовательно, торможение центров спинного мозга явилось следствием возбуждения центров среднего мозга. Прекращение этого тормозного контроля при перерыве цереброспинальных проводящих путей вызывает резкое повышение возбудимости спинальных центров и гиперрефлексию.
- Принцип обратной афферентации - заключается в рецепторном восприятии результатов рефлекторного акта и проведении информации назад в структуры нервного центра, где она обрабатывается и сравнивается с сохраняющимися параметрами возбуждения. Обратная афферентация реализуется в виде положительной или отрицательной обратной связи. Таким образом, с помощью обратной афферентации нервные центры осуществляют непрерывный контроль эффективности, целесообразности и оптимальности рефлекторной деятельности.
Дата добавления: 2015-08-12 ; просмотров: 4180 . Нарушение авторских прав
Свойства нервных центров. К ним относят:
замедленное распространение возбуждения, обусловленное наличием синапсов и многих нейронов на путях передачи возбуждения;
одностороннее распространение возбуждения, обусловленное тем, что основную роль в распространении возбуждения играют химические синапсы, в которых возбуждение передается только от пресинаптических структур на постсинап- тические структуры;
иррадиация возбуждения — распространение возбуждения от активированного нервного центра на соседние и функционально связанные нейронные структуры. Это свойство обусловлено наличием многочисленных ветвлений аксонов, заканчивающихся синаптическими контактами. В результате при активации группы нейронов от них идут возбуждающие влияния на синаптически связанные с ними другие нейронные группы. При прекращении притока возбуждающей импульсации активность нервного центра уменьшается и может наступать концентрация возбуждения — процесс, обратный иррадиации;
суммация возбуждения, проявляющаяся увеличением интенсивности рефлекторной реакции при возрастании длительности раздражения, его силы или площади раздражаемого рецепторного поля. Различают временную и пространственную суммацию. При временной суммации количество активируемых синаптических образований на эфферентном нейроне не изменяется, но увеличивается частота импульсаций, приходящих к каждому синапсу. При пространственной суммации из-за воздействия на новые рецепторы увеличивается количество одновременно активируемых синаптических входов и эфферентный нейрон быстрее и эффективнее возбуждается. В обоих случаях возрастает величина ответной реакции эффекторных структур. Например, увеличивается интенсивность сокращения мышцы, секреции слезной или слюнной железы.
трансформация ритма возбуждений, проявляющаяся изменением частоты импульсаций, генерируемых эфферентными нейронами, по отношению к частоте импульсаций, передаваемых афферентными нейронами. Такое изменение становится возможным благодаря суммации ВПСП, функционированию локальных нейронных цепей и т.д.;
явление последействия, заключающееся в способности нервных центров длительно поддерживать возбуждение после устранения раздражителя, вызвавшего его. Одним из механизмов этого служит возможность длительной циркуляции возбуждения в замкнутых нейронных цепях;
тонус нервных центров, проявляющийся длительным непрерывным возбуждением нервных центров. Такое состояние поддерживается за счет непрерывного притока афферентных импульсаций, влияния биологически активных веществ, доставляемых гуморальным путем, циркуляцией возбуждения в замкнутых нейронных цепях, спонтанной активностью отдельных нейронов;
пластичность нервных центров — способность к перестройке функциональных свойств и изменению выполняемых функций. Например, экспериментально показана способ
но ность диафрагмальных мотонейронов вместо обеспечения ритмических сокращений диафрагмы выполнять функцию регуляции шагательных движений ног;
повышенная утомляемость нервных центров по отношению к утомляемости периферических нервных структур и мышц- В системе нервный центр — нервно-мышечный препарат первое место по утомляемости занимает нервный центр, второе — нервно-мышечный синапс, третье — скелетная мышца и наименее утомляемым оказывается нервное волокно;
высокая чувствительность нервных центров к действию биологически активных веществ и ядов, нарушениям гомеостаза и недостатку кислорода. При прекращении доставки кислорода клетки коры мозга могут сохранять жизнедеятельность не более 5 мин, уменьшение содержания глюкозы в крови ниже 800 мг/л сопровождается гипогликемической комой.
Принципы функционирования, интегративной и координационной деятельности ЦНС. Интегративная деятельность ЦНС проявляется восприятием, объединением и переработкой комплекса поступающей к нервным центрам информации и принятием решений, обеспечивающих регуляцию гомеостаза, а также вегетативных и соматических реакций, направленных на достижение полезного для организма приспособительного результата. Координационной деятельностью называют регуляцию распределения возбуждения и торможения в нейронных структурах, а также взаимодействия нервных центров, обеспечивающие адекватные рефлекторные и произвольно вызываемые реакции.
Принцип рефлекторных регуляций — важнейший принцип функционирования нервной системы.
Рефлексом называют стереотипную ответную реакцию организма на действие раздражителя, осуществляющуюся благодаря регуляторной роли нервных клеток, составляющих рефлекторную дугу.
Абсолютное большинство рефлексов замыкается в головном и спинном мозге. Но есть также рефлексы, рефлекторная дуга которых замыкается вне центральной нервной системы в вегетативных внеорганных ганглиях или даже в пределах одного органа (например, сердца или кишечника).
Рефлекторная дуга включает 5 звеньев. Начальным звеном является рецептор, затем идет афферентный (чувствительный, Центростремительный) нейрон, ассоциативный (вставочный)
нейрон, эфферентный (двигательный, центробежный) нейрон и эффектор. Эффектором может быть любая структура (мышца, железа и т.д.), на которой заканчивается синапсом эфферентный нейрон. Вставочный нейрон может быть один или их может быть много. Они располагаются в нервных центрах.
Следовательно, в образовании рефлекторной дуги участвует как минимум 3 нейрона. Исключение составляет лишь один вид рефлексов так называемые сухожильные рефлексы, рефлекторная дуга которых включает только 2 нейрона: афферентный и эфферентный. Отросток афферентного нейрона входит в спинной мозг через задние корешки и, проникая в передние рога серого вещества, формирует синапс прямо на эфферентном нейроне. Примеры схем двух- и трехнейронной рефлекторной дуги соответственно сухожильного и оборонительного сгибательного рефлекса, вызываемого раздражением кожи, представлены на рис. 6.4.
Область сосредоточения рецепторов, с которых запускается определенный рефлекс, называют рецепторным полем этого рефлекса.
Разрез спинного мозга
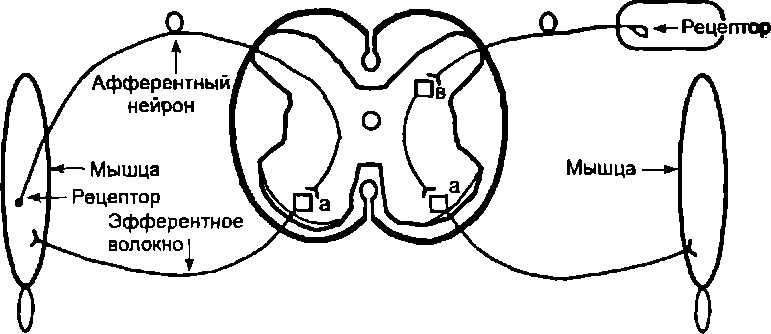
Рис. 6.4. Схема двухнейронной рефлекторной дуги сухожильного рефлекса и трехнейронной рефлекторной дуги сгибательного оборонительного
рефлекса: а — а-мотонейрон, в — вставочный нейрон
Классификация рефлексов: все рефлекторные реакции подразделяют на безусловные и условные. Безусловные врожденные и проявляются при воздействии специфического
раздражителя на строго определенное рецепторное поле. Условные — приобретаются или вырабатываются в процессе жизни. Подробная их характеристика будет дана при изучении высшей нервной деятельности. Здесь же рассмотрим наиболее употребительные варианты классификации безусловных рефлексов.
По биологической значимости рефлекторной реакции выделяют пищевые, оборонительные, половые, ориентировочные, статокинетические рефлексы.
По виду рецепторов различают эстероцептивные, инте- роцептивные, проприоцептивные рефлексы. Среди последних выделяют сухожильные и миотатические рефлексы.
По участию в рефлексе соматических или вегетативных отделов ЦНС и органов эффекторов различают соматические и вегетативные рефлексы. Если эффектор и рецепторное поле рефлекса относятся к соматическим структурам, то такие рефлексы называют соматическими. Вегетативными называют рефлексы, эффектором в которых являются внутренние органы, а эфферентная часть рефлекторной дуги образована вегетативными нейронами. Примером вегетативного рефлекса является рефлекторное замедление сердечной деятельности, вызванное воздействием на рецепторы желудка. Примером соматического рефлекса является сгибание руки в ответ на болевое раздражение кожи.
По уровню замыкания рефлекторной дуги в центральной нервной системе выделяют спинальные, бульварные (замыкающиеся в продолговатом мозге), мезенцефальные и талами- ческие рефлексы.
По количеству нейронов, участвующих в замыкании рефлекторной дуги, и числу центральных синапсов выделяют двухнейронные, трехнейронные, мультинейронные; моносинап- тические, полисинаптические рефлексы.
Благодаря принципу обратной связи (обратной афферентации) формируется замкнутый регуляторный контур. Только замкнутая регуляторная система может обеспечить устойчивое, адекватное регулировние. Благодаря обратной связи осуществляется контроль за исполнением, результатом влияния управляющего сигнала на эффекторные структуры и величиной регулируемого параметра.
Участие обратной связи даже в простейших регуляциях можно рассмотреть на примере реализации сгибательного
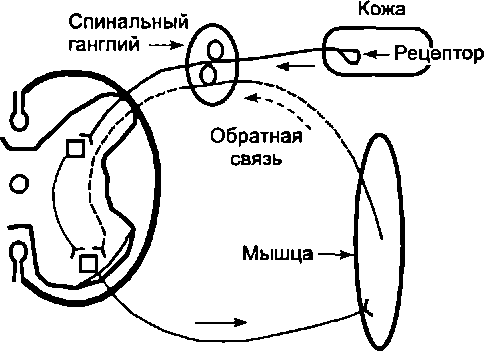
Рис. 6.5. Обратная связь в простейших рефлекторных реакциях
рефлекса, вызываемого прикосновением к коже (рис. 6.5). При рефлекторном сокращении мышцы изменяется активность находящихся в ней рецепторов и частота импульсаций в афферентных волокнах, идущих к мотонейронам. В результате формируется замкнутый контур регулирования, в котором роль канала обратной связи выполняют афферентные волокна, передающие импульсацию в нервные центры от рецепторов мышц, а роль канала прямой связи — эфферентные волокна, идущие к мышцам от мотонейронов. Таким образом, нервный центр (его мотонейроны) получает информацию об изменении состояния мышцы, вызванном передачей импульсов по двигательным волокнам. Благодаря обратной связи формируется своеобразное регуляторное кольцо. Поэтому некоторые авторы даже предлагали вместо термина "рефлекторная дуга" применять термин "рефлекторное кольцо"
Участие обратной связи в механизме регуляции кровообращения, дыхания, температуры тела и поведенческих реакций рассматривается ниже в соответствующих разделах.
Принцип реципрокной связи проявляется во взаимодействии центров-антагонистов по функциональному назначению, например группы мотонейронов, ответственных за сгибание руки, и группы, управляющей мышцами-разгибателями руки. При реципрокной связи возбуждение одного из антагонистических центров приводит к торможению другого. Это осуществляется за счет активации тормозных нейронов на конечном участке проводящих путей, идущих от возбужденного центра к антагонисту.
Принцип конвергенции заключается в схождении, поступлении к одному и тому же нейрону импульсаций от различных нервных центров или рецепторов различных модальностей (различных органов чувств). На основе конвергенции самые разные раздражители могут вызвать однотипную реакцию. Например, сторожевой рефлекс (поворот головы, насторажи- вание) может быть вызван и световым, и звуковым, и тактильным воздействием. Возможность конвергенции импульсаций с разных входов на одни и те же эфферентные нейроны называют принципом общего конечного пути.
Принцип дивергенции утверждает возможность расхождения импульсаций от одного нейрона сразу на многие нейроны. На основе дивергенции происходит иррадиация возбуждения и становится возможным быстрое вовлечение в ответную реакцию многих центров, расположенных на разных уровнях ЦНС.
Принцип доминанты характеризует особенности взаимодействия нервных центров. Доминантный очаг возбуждения обладает стойкой высокой активностью, он подавляет возбуждение в других нервных центрах, подчиняет их своему влиянию, притягивает к себе афферентные импульсации, адресуемые к другим центрам, и усиливает свою активность за счет этих импульсаций. Доминантный центр может длительно находиться в состоянии возбуждения без признаков утомления.
Примером рефлекторной реакции, вызванной доминантным очагом возбуждения, может служить обнимательный рефлекс самца лягушки, проявляющийся в период размножения. Если в этот период положить самца лягушки брюшком на палец, то у животного возникает обнимательный рефлекс. Самец обхватывает и сжимает передними лапками палец. Затем металлическим пинцетом проводят по спинке животного. Это воздействие обычно вызывает оборонительную реакцию (бегства). Но в данных условиях воздействие пинцетом лишь усиливает обнимательный рефлекс, самец сильнее сдавливает палец.
Контрольные вопросы и задания
Какова роль нервной системы в организме ?
Как подразделяют нервную систему ? Какие клетки ее образуют?
Какова роль глиальных клеток в ЦНС?
Каковы структура и функции нейронов и их частей?
Каковы особенности поляризации мембрены нейрона и возбудимости его отделов?
Каковы особенности передачи возбуждения в центральных синапсах? Что такое медиаторы?
7 Перечислите виды и механизмы торможения.
Что такое интеграция возбуждения на нейроне?
Какова классификация нейронов?
Что такое нейронные цепи?
Каковы свойства нервных центров?
Дайте характеристику рефлекса и рефлекторной дуги.
Какова классификация рефлексов?
Каковы принципы осуществления нервных регуляций?
Как соотносится длительность латентных периодов сухожильных и оборонительных рефлексов, вызываемых раздражением кожи? Почему имеются такие различия?
Зарисуйте схему нейронной сети, которая может увеличить частоту эфферентных импульсов по отношению к частоте афферентных.
Какие механизмы обусловливают способность спинальных мотонейронов генерировать частоту возбуждения до 50 имп/с, а вставочных — до 500 имп/с?
Почему при наличии обнимательного рефлекса у самца лягушки ряд раздражителей перестает вызывать оборонительную реакцию, а вместо нее лишь усиливает обнимательный рефлекс?
Принцип доминанты сформулирован А.А Ухтомским, Под доминантой А.А.Ухтомский понимал господствующий очаг возбуждения, определяющий характер текущих реакций центров в данный момент. Этот господствующий центр может возникнуть при длительном действии гуморальных или рефлекторных раздражителей. Для него характерно: а) наличие повышенной возбудимости; б) иннерционность, обусловленная длительными следовыми процессами; в) способность к суммации и сопряженному торможению других центров, функционально несовместимых с деятельностью центров доминантного очага. Доминанта создаëт определенный вектор поведения.
Примером может служить обнимательный рефлекс у лягушки в результате гормональных воздействий. Гормоны в сочетании с раздражением кожи (рецепторы – мозоли пальцев передних конечностей самца) создают суммацию раздражителей.
А.А.Ухтомский считал, что доминанта является общим принципом работы нервных центров, что она связана с возбуждением или активацией многих нервных центров, временно кооперирующихся для выполнения биологически важной функции. Взаимодействие работающих центров на единый ритм активности (усвоение ритма) рассматривается как динамический функциональный орган.
Согласно современным представлениям (теория функциональных систем П.К.Анохина) любой поведенческий акт, в том числе условный рефлекс – результат анализа и синтеза афферентной информации, включающей, прежде всего, возбуждение центральных рецепторов латерального, переднего гипоталамуса, преоптической области, реагирующих на содержание в крови питательных веществ, половых гормонов и других факторов. Доминирующий очаг может возникнуть не только под влиянием гормональных воздействий, но и под влиянием нервных сигналов, изменяющих возбудимость центральных нейронов.
Иннерционность доминанты - это способность доминантного состояния поддерживаться довольно долго, что обусловлено длительностью следовых процессов. Следовое возбуждение в свете открытий современной физиологии может быть обусловлено следующими факторами:
Суммацией взаимодействия возбуждающих потенциалов, вызванных приходящими к нейронам подпороговыми нервными импульсами.
1.Изменением концентрации ионов К + в синаптической щели, которые как деполяризаторы, усиливают вхождение ионов Са² + в пресинаптическое окончание.
2.Следами метаболитов, связанных с медиаторами.
3.Циклическими связями в ЦНС, способными обеспечить следовую самостимуляцию центров (следовые процессы храняться различное время в гиппокампе – часы, сутки – предполагают, что здесь хранятся следовые процессы от пережитых доминат; в спинальных центрах – постсинаптический потенциал длится минуты).
Доминатное состояние как системная реакция может не только суммировать посторонние раздражения, но и может тормозить конкурирующие центры.
Феномен сопряженного торможения конкурирующих центров возможен благодаря динамическому функциональному органу как временной кооперации нервных центров.
Существуют другие теории, объясняющие сопряженное торможение. В 1957 году В.Маунткасл (в опытах использовал кошек, вводя в сенсомоторную кору электроды под углом и перпендикулярно) выдвинул гипотезу колончатой организации коры больших полушарий, согласно которой головной мозг – это совокупность нейронных модулей, связанных многократно. Доминирование одного из модулей и вызывает динамику систем.
Проведение возбуждения через нервные центрыосуществляется тольков одном направлении: с афферентного нейрона на эфферентный. Свойство объясняется строением синапсов – медиаторы выделяются только концевыми аппаратами аксонов и возбуждение распространяется к постсинаптической мембране.
Задержка проведения в синапсах и время рефлекса обусловлена более медленным проведением нервных импульсов через синапсы, т.к. необходимо время для выделения медиатора, его диффузию через синаптическую щель к постсинаптической мембране и возникновения возбуждающего постсинаптического потенциала под действием медиатора. Чем больше сила раздражителя, тем короче время рефлекса.
Иррадиация возбуждения-это способность возбуждения широкой волной разливаться по ЦНС от центра к центру во всех направлениях, что объясняется наличием огромного количества коллатералей. От каждого аксона отходят идут коллатерали к множеству нейронов, от них идет еще большее число коллатералей. Т.о. импульс, пришедший в ЦНС, может иррадиировать по многим направлениям ко многим центрам.
Конвергенции -свойство нервных центров, противоположное иррадиации. Схождение, сужение – к эфферентному нейрону возбуждение проходит по многим путям, т.к. афферентных путей в 4-5 раз больше.
Циркуляция нервных импульсов по замкнутым нейронным цепям
может длительно осуществляться в одном нервном центре благодаря тому, что импульсы, возникнув в одном из нейронов, передаются к другим нейронам, а затем по коллатералям их аксонов возвращаются на первый нейрон.
Утомляемость нервных центров обуславливается высоким обменом веществ, что приводит к нарушением проведения возбуждения в межнейронных синапсах. Уменьшаются запасы медиатора в окончаниях аксона, падает чувствительность к медиатору постсинаптической мембраны, понижаются ресурсы нервной клетки.
Пластичность нервных центров-перемена функций нервного центра в случае, если рабочий орган, с которым данный орган связан, заменить другим.
Определение понятия нервного центра
Нервный центр — это совокупность связанных между собой нейронов, совместно выполняющих определённую функцию путём преобразования входящего возбуждения в выходящее с изменёнными характеристиками.
Данное определение содержит 7 критериев нервного центра, найдите и назовите их.
Суперкраткое определение: Нервный центр - это "полисинаптический трансформатор возбуждения".
Нервный центр - это та нервная структура, которая связывает сенсорную систему с эффекторной и преобразует сенсорное возбуждение в эффекторное или модулирующее.
Нервный центр - это многозначное понятие.
Анатомический подход: нервный центр - это совокупность сходных нервных клеток, выполняющих общую функцию и компактно расположенных в определенном участке ЦНС.
Здесь используется морфологический подход, т.е. нервные центры определяются по строению. Нервные клетки, образующие такой нервный центр, соединены в локальные компактные структуры: нервные узлы (ганглии) в периферической нервной системе или мозговые ядра в центральной нервной системе.
Физиологический подход (по деятельности) : нерный центр - это система взаимосвязанных клеток, которые объединяются для выполнения определённой функции, а физически могут находится в различных местах нервной системы.
Такое определение созвучно понятию "функциональной системы", которое было предложено П.К. Анохиным. Но если функциональная система подразумевает временное объединение нейронов, то нервный центр - это обычно устойчивое образование. По П.К. Анохину различные анатомические нервные центры при необходимости могут на время объединяться в функциональную систему для получения определенного полезного результата.
Нервные центры биорегуляции. В функциональном отношении нервный центр также может представлять собой сложное объединение нескольких анатомических нервных центров, расположенных в разных отделах ЦНС и обусловливающих сложные рефлекторные акты. В этом смысле говорят о "пищевом нервной центре", "болевом нервном центре" и т.п. Это центры регуляции функций организма.
Понятие нерный центр может также пересекаться с понятием доминанты. Доминанта - это устойчивый очаг возбуждения, подчиняющий себе другие очаги . Кроме единичного очага возбуждения в состав доминанты также могут входить и другие связанные с ним очаги. Создатель учения о доминанте А.А. Ухтомский называл такие объединения возбуждённых очагов "созвездиями" ("констелляциями").
Эволюция выбирает путь концентрации нервных элементов и увеличение количества связей между ними. Поэтому из рассеянных диффузно нервных клеток образуются компактные нервные центры.
Линия эволюционного развития структуры нервной системы
1) Отдельные нервные клетки соединены примерно равными связями.
2) Нервные цепи - нервные клетки соединены последовательно более сильными связями, образуя предопределённый путь для движения возбуждения от одного пункта к другому.
3) Нервные сети - нервные клетки соединены в виде решеток с неравноценными связями.
4) Нервные узлы (ганглии) - нервные клетки собраны в компактные структуры, соединёнными между собой продолными связями (коннективами) и поперечными (комиссурами) в виде лестницы.
5) Нервная трубка - нервные клетки расположены в виде сплошных слоёв, закрученных в виде трубы.
6) Нервные ядра - обособленные нейронные структуры из тесно связанных сходных между собой нейронов в составе нервной трубки, специализирующиеся на определённых функциях.
7) Нейронные поля — зоны коры головного мозга. Они состоят из вертикальных колонок, в которые сгруппированы нейроны.
Свойства нервных центров
Наличие входов и выходов для возбуждения . В нервном центре можно различить приносящие (афферентные) входы и выносящие (эфферентные) выходы.
Одностороннее проведение возбуждения. Это свойство отдельного синапса и нервной цепи. В нервном центре может быть множество путей между входами и выходами. За счёт обратных связей возможно возвратное движение возбуждения. Но это происходит внутри нервного центра. А если рассматривать нервный центр целиком, то возбуждение приходит внего по приходящим путям, а выходит по эфферентным выходящим. Таким образом, можно говорить об одностороннем проведении возбуждения нервным центром.
З адержка ( замедление ) проведения возбуждения . В нервных центрах возникает задержка в проведении возбуждения, так называемый латентный (скрытый) период. Задержка обусловлена синаптической передачей возбуждения. Чем больше синапсов участвует в проведении возбуждения, тем более длительной получается задержка.
Суммация возбуждения . Если одновременно подавать возбуждение на несколько входов нервного центра, то на выходе можно получить более сильное возбуждение. Свойством суммации обладает и отдельный нейрон за счёт суммации локальных потенциалов.
Трансформация (преобразование) входящего возбуждения в иное - выходящее . Нервный центр осуществляет изменение, перекодирование поступающих в него потоков импульсов. Трансформация возбуждения - это, пожалуй, самое главное свойство нервного центра. Наиболее известное свойство из этого ряда – трансформация ритма. Нервный центр получает на входе один ритм импульсации, а на выходе дает другой (более медленный или более частый).
Последействие (облегчение) . Это означает, что после возбуждения нервного центра он некоторое время ещё сохраняет повышенную возбудимость. Поэтому последующее возбуждение даёт более сильный эффект и получение эффекта от работы нервного центра облегчено.
Утомляемость и низкая лабильность. Лабильность - это предельная частота импульсации, доступная данной нервной структуре. Нервные центры могут пропускать через себя потоки возбуждения с ограниченной частотой импульсации вследствие задержки передачи возбуждения, которая происходит в многочисленных синапсах. Повышенная утомляемость нервных центров объясняется высокой утомляемостью синапсов и ухудшением метаболизма (обменнных процессов) в нейронах после нагрузки.
Тонус . Это означает, что даже без внешнего воздействия нервный центр сохраняет определённый уровень возбудимости и самостоятельно поддерживает у себя определённый уровень возбуждения.
Чувствительность к кислороду и к действию биологически активных веществ (нейротропных). Это создаёт предпосылки к хеморегуляции - химическому управлению деятельностью нервного центра. Например, усиление или ослабление кровоснабжения изменяет работу нервных центров.
В озбудимость ( возбуждение ) . Это способность нервных центров переходить в более возбуждённое состояние, например, при внешнем воздействии на них (стимуляции) или под влиянием других нервных центров.
Торможение ("тормозимость") . Это способность нервных центров переходить в менее возбуждённое состояние, например, при внешнем воздействии на них или под влиянием других нервных центров.
Иррадиация возбуждения . Это "растекание возбуждения" по нервному центру, распространение возбуждения на новые участки от места его первоначального появления.
Конвергенция (схождение) . Это объединение двух или нескольких входящих потоков возбуждения в один выходящий поток. Т.е. в нервный центр входит больше потоков возбуждения, че выходит из него.
Дивергенция (расхождение) . Это разделение входящего потока возбуждения на несколько выходящих потоков. За счёт дивергенции получается, что в нервный центр входит меньше потоков возбуждения, чем выходит из него.
Окклюзия (запирание) . Это блокирование одним из входящих потоков возбуждения другого входящего потока. В результате выходящий поток возбуждения получается слабее, чем сумма этих входящих потоков.
Индукция (отдача) . Это наведение противоволожного (возбуждённого или тормозного) состояния на другие нервные центры или на себя самого. Для понятия индукции очень важно, что данной структурой наводится именно противоположное состояние, а не то, в котором находится она сама. Так, возбуждённая структура индуцирует торможение, а заторможенная - возбуждение.
Автоматия (спонтанная активность, автономность) нервных центров. Это означает, что даже без внешнего воздействия нервный центр может самостоятельно порождать возбуждение на выходе или поддерживать свой тонус (как бы развлекать сам себя). Объясняется это свойство нервного центра существованием в нём специальных нейронов-пейсмекеров (водителей ритма). В них самопроизвольно возникает возбуждение, независимо от работы их афферентных входов. Таким образом, в нервных центрах может происходить периодическая или постоянная генерация (порождение) нервных импульсов, которые возникают даже при отсутствии входящего возбуждения. Самопроизвольная импульсация пейсмекеров обусловлена колебаниями процессов метаболизма в нейронах и действием на них гуморальных факторов.
Реципрокные (взаимоисключающие) отношения. Это означает, что возбуждение одного нейрона (или центра) подавляет работу другого, связанного с ним, нейрона (или центра).
П ластичность . Это способность перестраивать свою структуру и\или деятельность под влиянием предыдущей деятельности. Пластичность - это одно из важнейших свойств биологических систем, которое отличает их от технических систем.
Адаптация. Нервный центр способен приспосабливаться к новой нагрузке и новым условиям работы.
Компенсаторные возможности. При частичном повреждении нервный центр продолжает свою деятельность за счёт сохранившихся нейронов. Для этого он использует свои способности к пластичности и адаптации.
Основные принципы в работе нервных центров
Принцип общего конечного пути ("воронка Шеррингтона"). Как правило, центры имеют больше афферентных входов, чем эфферентных выходов. Поэтому входящие потоки возбуждения конкурируют за выход, имея общий конечный путь. В итоге количество афферентных входов превышает количество эфферентных выходов.
Принцип обратной связи. Это означает, что последующий элемент (нейрон или центр) в последовательной цепи взаимосвязанных элементов влияет на состояние предыдущего элемента. Обратная связь позволяет произвести отладку взаимодействия между элементами и добиться их оптимального взаимодействия для достижения предельно возможного положительного результата в работе системы, состоящей из этих элементов.
Принцип доминанты. Это означает, что нервный путь или нервный центр наиболее активный получает преимущество по отношению к другим путям или центрам и начинает доминировать, господствовать над ними. Он тормозит их деятельность и перехватывает их возбуждение, чтобы усилить своё.
Принцип иерархии (соподчинения). Это означает, что одни элементы (нейроны и/или центры) подчиняются влиянию других элементов. Как правило, вышерасположенные центры подчиняют себе нижерасположенные центры.
Принцип пластичности. Это означает, что нервный центр перестраивает свою деятельность, приспосабливаясь к наилучшему выполнению своей функции для достижения общего конечного системного результата. Пластичность является важнейшей отличительной особенностью биосистем по сравнению с техническими системами.
Низшие нервные центры
Низшие нервные центры играют важную роль в работе любой сенсорной системы. Они являются одним из необходимых элементов сенсорной системы, по которому это понятие отличается от понятия "анализатор". Нервные центры не просто переключают возбуждение с одних нейронов на другие с помощью вставочных нейронов, т.е. выполняют "релейную" функцию, как это считалось ранее. Важно понять, что нервные центры занимаются трансформацией поступающего в них возбуждения, т.е. его преобразованием, или перекодированием. В результате этой трансформации входящее афферентное возбуждение превращается в выходящее эфферентное , отличающееся от входящего.
Работа (функции) низших нервных центров
1. Трансформация возбуждения , т.е.преобразование входящего потока сенсорного возбуждения в новый поток - выходящий. Выходящий поток может сильно отличаться от входящего, например, в том случае, если он должен управлять мышцами, а не строить нервную модель раздражения в виде сенсорного образа.
Виды трансформации возбуждения в нервном центре
1. Усиление.
2. Ослабление.
3. Блокировка.
4. Изменение паттерна (узора, характера).
5. Контрастирование границ в пространстве. Обычно оно достигается с помощью латерального (бокового) торможения. Латеральное торможение усиливает возбуждение по контуру раздражителя и рецептивного поля и ослабляет возбуждение в центральной области рецептивного поля.
6. Контрастирование границ во времени. Происходит за счёт преобразования тонического (постоянного) возбуждения в кратковременное фазическое. Таким способом отмечаются начало и конец действия раздражителя.
2. Распределение входящих потоков сенсорного возбуждения по выходящим потокам, которые направляются в различные нервные структуры. Эта функция нервного центра наглядно показана с помощью нашей схемы "Пути сенсорного возбуждения".
Виды распределения возбуждения в нервном центре
1. Конвергенция (схождение).
2. Дивергенция (расхождение).
3. Окклюзия (запирание).
4. Иррадиация (распространение).
3. Детекция . С помощью детекции выделяются раздражители с определёнными характеристиками за счёт срабатывания специальных нейронов-детекторов с соответствующими рецептивными полями. На любые другие раздражители, неадекватные для них, такие нейроны-детекторы не срабатывают, т.к. просто не возбуждаются такими стимулами.
Читайте также:
