
Нервные рецепторы в мышцах
Содержание
- 1 Специфические рецепторы поперечнополосатых мышц
- 1.1 Мышечные веретена
- 1.2 Сухожильные органы Гольджи
- 2 Читайте также
Специфические рецепторы поперечнополосатых мышц [ править | править код ]
Помимо свободных нервных окончаний (ноцицепторов) и рецепторов соединительнотканных футляров в мышцах и области мышечно-сухожильных соединений имеется два типа важных рецепторов глубокой чувствительности:
- мышечные веретена;
- сухожильные органы Гольджи.
Эти рецепторы относятся к группе проприорецепторов, которые играют важную роль в выполнении движений и поддержании позы. Проприорецепторы передают информацию о позе тела в пространстве и расположении конечностей относительно туловища (чувство положения). Кроме этого, они регистрируют движения в суставах и их скорость (чувство движения) и могут оценивать приложенное мышечное усилие (чувство силы). Эта информация далее направляется вместе с импульсами от органов чувств и равновесия в высшие центры моторной деятельности (в том числе в мозжечок). Эти импульсы передают основную информацию для контроля движений и коррекции поддержания позы и целенаправленных движений.
Мышечные веретена расположены по всей мышце. Их название обусловлено их веретенообразной формой. Количество мышечных веретен в мышце варьирует от 40 до 500 (Lindel, 2006). Как уже было отмечено, они лежат вдоль мышечных волокон (экстрафузальных волокон, вне веретена) и имеют 5-10 мм в длину и 0,2 мм в толщину (Kolster, 2006). Мышечные веретена состоят из соединительнотканной капсулы, поверхностной слой которой соответствует эндомизию, и 10-20 так называемых интрафузальных мышечных волокон (внутри веретена). Концы веретена прикрепляются к соединительнотканной оболочке мышцы или к сухожилию. Интрафузальные волокна только на концах имеют актиновые и миозиновые филаменты, позволяющие им сокращаться, а в центре волокно способно лишь к растяжению.
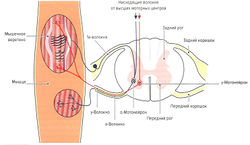
Несократительный растяжимый центр интрафузального волокна окружен толстым спиралевидным миелинизированным нервным окончанием типа 1а (Аа-волокна) (кольцеспиральное окончание) (рис. 1.6, а). Диаметр la-волокон составляет 10-20 мкм. Данные нервные волокна являются самыми быстрыми в организме человека — скорость распространения импульса составляет 60-120 м/с. Сократительные концы интрафузальных волокон иннервируются эфферентными у-волокнами (Ay-волокна, у-мотонейроны). Диаметр этих волокон составляет 4-8 мкм, а скорость передачи импульса — 20-50 м/с. Около 50 % всех мотонейронов передних рогов одного сегмента спинного мозга представлены у-мотонейронами (Cabri, 1999).
Растяжение центральной части интрафузальных волокон приводит к раздражению кольцеспиральных нервных окончаний мышечного веретена (рецептор растяжения). На конце афферентных волокон возникает потенциал действия, который затем передается в ЦНС. Центральная часть веретена растягивается либо при растяжении всей мышцы, либо при напряжении сократительных концов интрафузальных волокон при активации у-мотонейронов. При одновременном действии этих двух факторов возможно как взаимное усиление, так и уменьшение их действия. Таким образом, мышечные веретена способны регистрировать как изменение длины мышцы, так и скорость ее изменения, а также косвенно ее изменять. Действуя через различные контуры регуляции, мышечные веретена выполняют несколько функций.
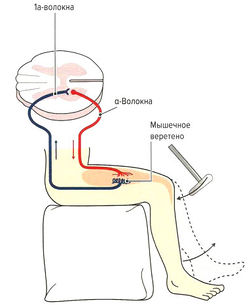
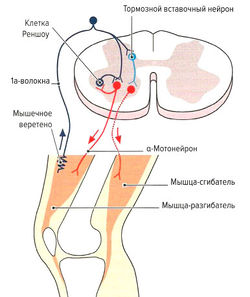
- Защита мышцы от перерастяжения за счет рефлекса растяжения (моносинаптический рефлекс). Функцией моносинаптического рефлекса растяжения является поддержание мышечной длины. Любое растяжение мышцы, например при ударе по сухожилию или мышце (рис. 1.6, б) или при изменении позы туловища, влияет на натяжение мышечных веретен. В результате резко повышается частота импульсации по 1а-афферентным нервным волокнам, идущим через задние спинномозговые корешки в серое вещество спинного мозга и через один синапс (моносинаптически) связываются с а-мотонейронами этой же мышцы (проприоцептивный рефлекс) (рис. 1.6, б). а-Мотонейроны по а-волокнам вызывают сокращение экстрафузальных волокон натянутой мышцы. Поскольку рецептор и эффектор находятся в одном и том же органе, скорость данного рефлекса очень высока (около 30 мс). Непроизвольные изменения длины мышцы могут сразу корректироваться на уровне спинного мозга без вовлечения высших моторных центров. Одновременно с этим через тормозной вставочный нейрон (ингибирующий la-нейрон) происходит ингибирование мотонейронов мышц-антагонистов (полисинаптический рефлекс) (реципрокное ингибирование антагонистов) (рис. 1.6, в). Благодаря этому растянутая мышца не преодолевает сопротивление антагонистов и предотвращается аналогичная реакция антагонистов при их растяжении (Weiss, 2000). После сокращения растянутой мышцы импульсация от la-волокон снижается. Дополнительно вследствие активации а-мотонейронов через коллатеральные связи и тормозные вставочные нейроны (клетки Реншоу) происходит обратное ингибирование самих а-мотонейронов (рис. 1.6, в). Эта отрицательная связь останавливает распространение сигнала и обеспечивает кратковременность сокращения мышцы, поэтому, несмотря на постоянные внешние раздражители (растяжение мышцы), возможно поддержание ее постоянной длины. Кроме этого, остановка рефлекторного ответа обеспечивается сухожильными органами Гольджи, которые активируются при мышечном сокращении и последующем растяжении сухожилия (аутогенное ингибирование) (см. рис. 1.7, б). В заключение необходимо отметить, что возбудимость мотонейронов контролируется высшими центрами головного мозга. При повреждении этих центров или отходящих от них нервных путей (например, при параплегии) развивается аномальное повышение рефлексов (см. рис. 1.6, а).
- Регуляция активного мышечного тонуса. Высшие моторные центры в головном мозге через так называемые экстрапирамидные пути активируют у-мотонейроны и через мышечные веретена регулируют длину мышц и, следовательно, активный мышечный тонус. Активация у-мотонейронов вызывает напряжение сократительных концов интрафузальных волокон, что растягивает центр мышечных веретен и приводит к активации la-волокон. Как уже было отмечено, импульс затем передается через задние рога в передние рога спинного мозга и возбуждает а-мотонейроны. а-Мотонейроны, в свою очередь, вызывают сокращение волокон скелетных
мышц, что опосредованно уменьшает натяжение центральной части веретен. Эта рефлекторная реакция продолжается до тех пор, пока 1а-волокна не будут регистрировать натяжение (у-петля). При нормальной позе (сидя, стоя или при ходьбе) в большинстве мышц, особенно в работающих против силы тяжести, регистрируется постоянная невысокая активность афферентных волокон мышечных веретен, что позволяет поддерживать определенный активный мышечный тонус. Мышцы, действующие против силы тяжести, например мышца, разгибающая позвоночник, в положении стоя находятся в постоянном небольшом натяжении, т. к. туловище под влиянием силы тяжести наклоняется несколько кпереди. За счет постоянной активации у-мотонейронов достигается дополнительное растяжение средней части мышечных веретен, что вызывает активацию высших моторных центров для поддержания мышечного тонуса. Благодаря постоянной фоновой афферентной импульсации во время обычных движений мышечные веретена могут воспринимать как укорочение, так и растяжение мышцы. При усилении или уменьшении активности у-мотонейронов изменяется чувствительность мышечных веретен и регулируется их активность при конкретных позе и движении. Таким образом, у-мотонейроны устанавливают в каждой конкретной ситуации оптимальную рабочую область мышечных веретен, и они могут регистрировать мышечную длину и скорость ее изменения. Это позволяет устанавливать и поддерживать постоянный активный мышечный тонус, а следовательно, поддерживать равновесие и положение суставов и определенную позу (чувство позы и опоры).
- Выполнение точных движений и регуляция тонкой моторики. Возможность регулировать чувствительность мышечных веретен, особенно вместе с произвольными движениями, играет большую роль. При произвольных движениях, например при разгибании в коленном суставе, происходит помимо активации а-мотонейронов и повышение активности у-мотонейронов. За счет а-у-коактивации устанавливается оптимальная длина мышечных волокон, чтобы поддерживать их чувствительность во время всего движения. При сокращении регистрируется растяжение мышцы и становятся возможными очень точные движения. Крохме того, в научной среде продолжаются дискуссии, являются ли точные движения конечностями исключительно результатом изменения у-активности. Супраспинальные нервные центры могут изменять у-активность в у-петле (изменение частоты импульсации по 1а-волокнам) и таким образом изменять активность связанных с ними а-мотонейронов (Weiss, 2005).

Запомните: Чем больше мышечных веретен в мышце, тем более точные и дифференцированные движения возможны при ее сокращении. Например, в мышцах кисти и глазного яблока находится большое количество проприорецепторов (Kolster, 2005).
Лекция 2
Функционирование рецепторного аппарата мышц
При изучении анатомии и физиологии (А.С. Солодков, Е.Б. Сологуб, 2001) вы изучали двигательную сенсорную систему. Одним из отделов этой системы являются проприорецепторы, расположенные в мышцах, сухожилиях и суставных сумках. В мышцах расположены мышечные веретена, в сухожилиях – сухожильные органы Гольджи. В суставных сумках расположены рецепторы суставов.
Еще в XIX веке В. Кюне обнаружил в скелетных мышцах структуры, напоминающие веретено. Затем, в начале XX века Нобелевский лауреат Чарльз Скотт Шеррингтон показал, что эти структуры служат чувствительными рецепторами. Мышечные веретена рассеяны по всем скелетным мышцам. Концы их обычно прикрепляются к мышечным волокнам параллельно. Каждое веретено покрыто капсулой, которая расширятся в центре и образует ядерную сумку. Внутри веретена содержатся интрафузальные мышечные волокна. Эти волокна в 2-3 раза тоньше обычных (экстрафузальных) волокон скелетных мышц.
Интрафузальные волокна подразделяются на два типа:
- Длинные и толстые (диаметр 20-25 мкм)[1], которые информируют ЦНС о динамическом компоненте движения – скорости изменения длины мышцы. Таких волокон в мышечном веретене не более двух.
- Короткие и тонкие (диаметр 10–12 мкм), которые информируют ЦНС о статическом компоненте движения – текущей длине мышцы. Таких волокон в мышечном веретене от 2 до 12.
Нервно-сухожильные веретена (рецепторы Гольджи) открыл в 1903 году Камилло Гольджи. Впоследствии за эти исследования ему была присуждена Нобелевская премия. Рецепторы Гольджи располагаются в месте перехода мышечных волокон в сухожилия. Их длина составляет 0,5-1,0 мм, а диаметр – 0,1-0,2 мм. Отдельный нервный аксон несет афферентные импульсы в спинной мозг и называется аксоном Ib. Он начинается в виде веточек, проходящих между коллагеновыми волокнами сухожилия. Когда мышечные волокна сокращаются, коллагеновые волокна сухожилия натягиваются и сжимают нервные веточки, которые начинают импульсировать (рис.2.1). Таким образом, в результате последовательного крепления сухожильных органов к мышечным волокнам они возбуждаются при укорочении возбужденной мышцы. Сухожильные рецепторы возбуждаются в 1,5 – 8 раз более эффективно при мышечном сокращении, нежели при пассивном растяжении.
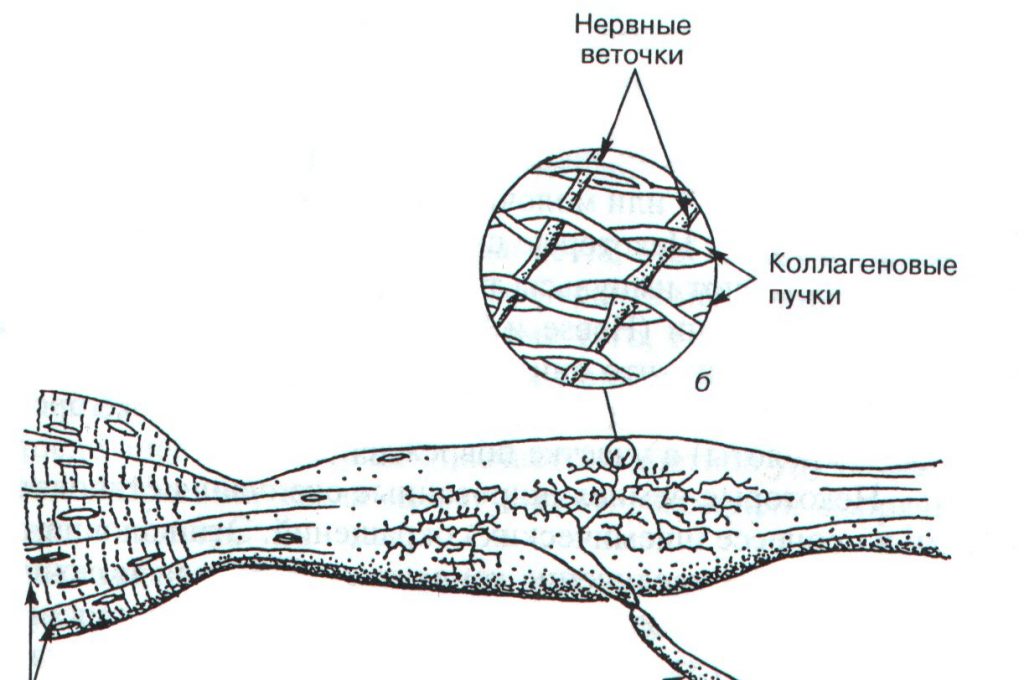
Рис. 2.1. Строение сухожильного органа Гольджи (А.Дж. Мак-Комас, 2001)
Суставные рецепторы подразделяются на несколько типов в зависимости от их реакции на амплитуду, скорость и направление движения в суставе.
Тельца Руффини находятся в капсуле сустава и воспринимают направление и скорость изменения межзвенного угла. Частота их импульсации возрастает с увеличением скорости изменения суставного угла.
Тельца Паччини посылают в ЦНС информацию о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы посылают в ЦНС информацию о значениях межзвенных углов, то есть о положении сустава. Их импульсация продолжается в течение всего периода сохранения межзвенного угла, и она тем больше, чем больше изменения угла.
Афферентация уровня А основана на импульсации мышечных веретен (длина и скорость сокращения мышцы) и рецепторов Гольджи (уровень возбуждения мышцы при ее укорочении). Эта информация очень слабо осознается ЦНС, то есть, по гипотезе Н.А.Бернштейна, этот уровень почти никогда не бывает ведущим.
Афферентация уровня В опирается на информацию, поступающую от суставных рецепторов. Это уровень выступает как ведущий в ряде физических упражнений, таких как наклоны тела вперед и назад, а также циклические движения. Сигналы от суставных рецепторов хорошо осознаются.
2. Самсонова А.В. Биомеханика мышц: учебно-методическое пособие /А.В.Самсонова, Е.Н. Комиссарова / Под ред А.В.Самсоновой .- СПб., 2008.- 217 с.
3. Самсонова А.В. Моторная и сенсорная функции мышц в биомеханике локомоций: монография /А.В.Самсонова; СПбГУФК им. П.Ф.Лесгафта, 2007.- 152 с.
Большинство обычных сенсорных рецепторов (химических, температурных или механических) деполяризуется в ответ на стимул (такая же реакция, как и у обычных нейронов), деполяризация ведёт к высвобождению медиатора из аксонных окончаний. Однако существуют исключения: при освещении колбочки потенциал на её мембране возрастает — мембрана гиперполяризуется: свет, повышая потенциал, уменьшает выделение медиатора.
Существуют следующие виды рецепторов:
| Природа раздражителя | Тип рецептора | Место расположения и комментарии |
| • электрическое поле | • ампула Лоренцини en:Ampullae of Lorenzini | |
| • атмосферное давление | • барорецептор | |
| • химическое вещество | • хемосенсор | |
| • влажность | • гидрорецептор | |
| • механическое напряжение | • механорецептор | |
| • повреждение тканей | • ноцирецептор | В большинстве тканей с разной частотой. Болевые рецепторы — свободные нервные окончания, немиелинизированные. |
| • осмотическое давление | • осморецептор | |
| • свет | • фоторецептор | |
| • положение тела | • проприоцептор | |
| • температура | • терморецептор | |
| • электромагнитное излучение | • электромагнитные рецепторы |
Рецепторы кожи
- Болевые рецепторы.
- Тельца Пачини — капсулированные рецепторы давления в округлой многослойной капсуле. Располагаются в подкожно-жировой клетчатке. Являются быстроадаптирующимися (реагируют только в момент начала воздействия), то есть регистрируют силу давления. Обладают большими рецептивными полями, то есть представляют грубую чувствительность.
- Тельца Мейснера — рецепторы давления, расположенные в дерме. Представляют собой слоистую структуру с нервным окончанием, проходящим между слоями. Являются быстроадаптирующимися. Обладают малыми рецептивными полями, то есть представляют тонкую чувствительность.
- Диски Меркеля — некапсулированные рецепторы давления. Являются медленноадаптирующимися (реагируют на всей продолжительности воздействия), то есть регистрируют продолжительность давления. Обладают малыми рецептивными полями.
- Рецепторы волосяных луковиц — реагируют на отклонение волоса.
- Окончания Руффини — рецепторы растяжения. Являются медленноадаптирующимися, обладают большими рецептивными полями.
Рецепторы мышц и сухожилий
- Мышечные веретена — рецепторы растяжения мышц, бывают двух типов:
- с ядерной сумкой
- с ядерной цепочкой
- Сухожильный орган Гольджи — рецепторы сокращения мышц. При сокращении мышцы сухожилие растягивается и его волокна пережимают рецепторное окончание, активируя его.
Рецепторы связок
В основном представляют собой свободные нервные окончания (Типы 1, 3 и 4), меньшая группа — инкапсулированные (Тип 2). Тип 1 аналогичен окончаниям Руффини, Тип 2 — тельцам Паччини.
Рецепторы сетчатки глаза
Сетчатка содержит палочковые (палочки) и колбочковые (колбочки) фоточувствительные клетки, которые содержат светочуствительные пигменты. Палочки чуствительны к очень слабому свету, это длинные и тонкие клетки, сориентированные по оси прохождения света. Все палочки содержат один и тот же светочуствительный пигмент. Колбочки требуют намного более яркого освещения, это короткие конусообразные клетки, у человека колбочки делятся на три вида, каждый из которых содержит свой светочуствительный пигмент — это и есть основа цветового зрения.
Под воздействием света в рецепторах происходит выцветание — молекула зрительного пигмента поглощает фотон и превращается в другое соединение, хуже поглощающее свет волн (этой длины волны). Практически у всех животных (от насекомых до человека) этот пигмент состоит из белка, к которому присоединена небольшая молекула, близкая к витамину A. Эта молекула и представляет собой химически трансформируемую светом часть. Белковая часть выцвевшей молекулы зрительного пигмента активирует молекулы трансдуцина, каждая из которых деактивирует сотни молекул циклического гуанозинмонофосфата, участвующих в открытии пор мембраны для ионов натрия, в результате чего поток ионов прекращается — мембрана гиперполяризуется.
Чуствительность палочек такова, что адаптировавшийся к полной темноте человек способен увидеть вспышку света такую слабую, что ни один рецептор не может получить больше одного фотона. При этом палочки не способны реагировать на изменения освещённости когда свет настолько ярок, что все натриевые поры уже закрыты.
Литература
Wikimedia Foundation . 2010 .
- Нервно-секреторный эффектор
- TimeZero
НЕРВНАЯ СИСТЕМА — НЕРВНАЯ СИСТЕМА. Содержание: I. Эмбриогенез, гистогенез и филогенез Н.с. . 518 II. Анатомия Н. с. 524 III. Физиология Н. с. 525 IV. Патология Н.с. 54? I. Эмбриогенез, гистогенез и филогенез Н. е.… … Большая медицинская энциклопедия
НЕРВНАЯ СИСТЕМА — сложная сеть структур, пронизывающая весь организм и обеспечивающая саморегуляцию его жизнедеятельности благодаря способность реагировать на внешние и внутренние воздействия (стимулы). Основные функции нервной системы получение, хранение и… … Энциклопедия Кольера
Вегетативная нервная система — Симпатический (показан красным) и парасимпатический (показан синим) отделы автономной нервной системы Вегетативная нервная система (от лат. vegetatio возбуждение, от лат. vegetativus … Википедия
Центральная нервная система — основная часть нервной системы животных и человека, состоящая из скопления нервных клеток (нейронов) и их отростков; представлена у беспозвоночных системой тесно связанных между собой нервных узлов (ганглиев), у позвоночных животных и… … Большая советская энциклопедия
Нервная ткань — В этой статье не хватает ссылок на источники информации. Информация должна быть проверяема, иначе она может быть поставлена под сомнение и удалена. Вы можете … Википедия
рецептор — ▲ источник ↑ сигнал, для (кого), нервная система рецепторы элементы нервной системы, воспринимающие раздражения; приемники информации. экстерорецептор, экстероцептор. интерорецептор. механорецепторы. терморецепторы. хеморецепторы реагируют на… … Идеографический словарь русского языка
РЕЦЕПТОР — В наиболее общих терминах специализированная нервная летка или ее часть, которая преобразует физические стимулы в потенциалы рецептора. То есть клетка, чувствительная к определенной форме стимуляции и надежно претерпевающая определенную модель… … Толковый словарь по психологии
эфферентная система — (от лат. efferens (efferennis) выносящий) нервные пути, включающие центробежные нервные волокна (см. нейрон), проводящие нервные импульсы от нервных центров (кора головного мозга, спинальные ядра и др.) к периферическим устройствам (см.… … Большая психологическая энциклопедия
Иммунная система — Лимфоцит, компонент иммунной системы человека. Изображение сделано сканирующим электронным микроскопом Иммунная система подсистема, существующая у позвоночных животных и объединяющая органы и ткани, которые … Википедия
РЕФЛЕНСЫ — (от лат. reflexio отражение>, автоматические двигательные реакции в ответ на внешнее раздражение. Термин Р. заимствован из области физ. явлений и имеет в виду аналогию между нервной системой, отражающей раздражение в форме двигательной реакции, и … Большая медицинская энциклопедия
Теория по нормальной физиологии на тему: Синапс. Физиология мышечных волокон. В данной статье рассматривается механизм синаптической передачи, НМС…
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Синапс — это специфическое место контакта двух возбудимых систем (клеток) для передачи возбуждения.
По способу передачи сигналов:
- механические,
- химические,
- электрические.
По виду медиатора: холинэргические и др.
Нервно-мышечный синапс (НМС) — химический, передача с помощью медиатора ацетилхолина.
Синонимы к слову НМС:
- Нервно-мышечное соединение;
- Моторная концевая пластинка.

Аксоны нервных клеток на своих окончаниях теряют миелиновую оболочку , ветвятся, и концевые веточки аксона утолщаются. Это пресинаптическая терминаль или бляшка или пуговка , которая погружается в углубление на поверхности мышечного волокна.
Покрывающая концевую веточку аксона поверхностная мембрана называется пресинаптической мембраной , т.е. это мембрана, покрывающая поверхность синаптической бляшки (терминали аксона).
Мембрана, покрывающая мышечное волокно в области синапса, называется постсинаптической мембраной , или концевой пластинкой. Она имеет извитую структуру, образуя многочисленные складки, уходящие вглубь мышечного волокна, за счет чего увеличивается площадь контакта.

На постсинаптической мембране находятся белковые структуры — рецепторы , способные связывать медиатор . В одном синапсе количество рецепторов достигает 10-20 млн.
Между пре- и постсинаптическими мембранами находится синаптическая щель , размеры ее в среднем 50 нм, она открывается в межклеточное пространство и заполнена межклеточной жидкостью .
В синаптической щели находится мукополисахаридное плотное вещество в виде полосок, мостиков и содержится фермент ацетилхолинэстераза .
В пресинаптической терминали находится большое количество пузырьков или везикул , заполненных медиатором — химическим веществом посредником, осуществляющим передачу возбуждения.
В нервно-мышечном синапсе медиатор — ацетилхолин (АХ).

АХ синтезируется из холина и уксусной кислоты (ацетил-коэнзима А) с помощью фермента холинэстеразы. Эти вещества перемещаются из тела нейрона по аксону к пресинаптической мембране. Здесь в пузырьках происходит окончательное образование АХ.
3 фракции медиатора:
- Первая фракция — доступная — располагается рядом с пресинаптической мембраной.
- Вторая фракция — депонированная — располагается над первой фракцией.
- Третья фракция — диффузно рассеянная — наиболее удаленная от пресинаптической мембраны.
Механизм синаптической передачи



Ионы Ca вызывают образование специального белкового комплекса , который включает в себя везикулу и структуры, расположенные непосредственно около пресинаптической мембраны.
Они связаны между собой так называемыми белками экзоцитоза.
Часть белков расположена на везикулах (синапсин, синаптотагмин, синаптобревин), а часть — на пресинаптической мембране (синтаксин, синапсо-ассоциированный белок). Данный комплекс получил название секретосома .



Излитию содержимого пузырька в щель способствует белок синаптопорин , формирующий канал, по которому идет выброс медиатора.
Квант медиатора — количество молекул, содержащихся в одной везикуле.
На 1 ПД выбрасывается 100 квантов АХ.





На постсинаптической мембране возникает потенциал концевой пластинки (ПКП). Он является аналогом локального ответа (ЛО).
Потенциал действия на постсинаптической мембране не возникает ! Он формируется на соседней мембране мышечного волокна.
- связывание с рецептором,
- разрушение ферментов (ацетилхолинэстеразой),
- обратное поглощение в пресинаптическую мембрану,
- вымывание из щели и фагоцитоз.
События в синапсе :
- ПД приходит к терминали аксона;
- Он деполяризует пресинаптическую мембрану;
- Ca2+ входит в терминаль, что приводит к выделению АХ;
- В синаптическую щель выделяется медиатор АХ;
- Он диффундирует в щель и связывается с рецепторами постсинаптической мембраны;
- Меняется проницаемость постсинаптической мембраны для ионов Na+;
- Ионы Na+ проникают в постсинаптическую мембрану и уменьшают ее заряд — возникает потенциал концевой пластинки (ПКП) .
На самой постсинаптической мембране ПД возникнуть не может, так как здесь отсутствуют потенциалзависимые каналы, они являются хемозависимыми!
- ПКП суммируются и достигают КУД на соседнем участке мышечного волокна, что приводит к возникновению ПД и его распространению по мышечному волокну (около 5 м/с).
Достигнув пороговой величины, то есть КУД, ПКП возбуждает соседнюю (внесинаптическую) мембрану мышечного волокна за счет местных круговых токов.

Особенности проведения возбуждения в нервно-мышечном синапсе
Одностороннее проведение возбуждения — только в направлении от пресинаптического окончания к постсинаптической мембране.
Суммация возбуждения соседних постсинаптических мембран.
Синаптическая задержка — замедление в проведении импульса от нейрона к мышце составляет 0,5-1 мс. Это время затрачивается на секрецию медиатора, его диффузию к постсинаптической мембране, взаимодействие с рецептором, формирование ПКП, их суммацию.
Низкая лабильность — она составляет 100-150 имп/с для сигнала, что в 5-6 раз ниже лабильности нервного волокна.
Чувствительность к действию лекарственных веществ, ядов, БАВ, выполняющих роль медиатора.
Утомляемость химических синапсов — выражается в ухудшении проводимости вплоть до блокады в синапсе при длительном функционировании синапса. Главная причина утомляемости — исчерпание запасов медиатора в пресинаптическом окончании.
Законы проведения возбуждения по нервам:
- Закон функциональной целостности нерва.
- Закон изолированного проведения возбуждения.
- Закон двустороннего проведения возбуждения.
В зависимости от скорости проведения возбуждения нервные волокна подразделяются на 3 группы: A, B, C. В группе A выделяют 4 подгруппы: альфа, бетта, гамма и сигма.
Физиология мышечных волокон
- скелетная (40-50% массы тела),
- сердечная (менее 1%),
- гладкая (8-9%).
Физиологические свойства скелетных мышц:
- Возбудимость — способность отвечать на действие раздражителя возбуждением.
- Проводимость — способность проводить возбуждение из места его возникновения к другим участкам мышцы.
- Лабильность — способность мышцы сокращаться в соответствии с частотой действия раздражителя (200-300 Гц для скелетной мышцы).
- Сократимость — для мышцы является специфическим свойством — это способность мышцы изменять длину или напряжение в ответ на действие раздражителя.
Физические свойства скелетных мышц:
- Растяжимость — способность мышцы изменять длину под действием растягивающей силы.
- Эластичность — способность мышцы восстанавливать первоначальную длину или форму после прекращения действия растягивающей силы.
- Силы мышц — способность мышцы поднять максимальный груз.
- Способность мышцы совершать работу.
Режимы сокращения:
- Изотонический,
- Изометрический,
- Ауксотонический.
Изотонический режим — сокращение мышцы происходит с изменением ее длины без изменения напряжения (тонуса) (напр.: сокращение мышц языка).
Изометрический режим — длина постоянная, увеличивается степень мышечного напряжения (тонуса) (напр.: при поднятии непосильного груза).
Ауксотонический режим — одновременно изменяется длина и напряжение мышцы (характерен для обычных двигательных актов).
Механизм сокращения поперечно-полосатых мышц
Любая скелетная мышца состоит из мышечных волокон, которые, в свою очередь, состоят из множества тонких нитей — миофибрилл , расположенных продольно. Каждая миофибрилла состоит из протофибрилл — нитей сократительных белков: миозина (миозиновая протофибрилла), актина (актиновая протофибрилла).
Кроме сократительных белков в миофибрилле имеются два регуляторных белка: тропомиозин и тропонин .
Миозиновые волокна соединены в толстый пучок, от которого в торону актиновых нитей отходят поперечные мостики. У каждого мостика выделяют шейку и головку.
Нить актина располагается в виде 2 скрученных ниток бус. На ней имеются актиновые центры.

Тропомиозин в виде спиралей оплетает поверхность актина, закрывая в покое ее центры. Одна молекула тропомиозина контактирует с 7 молекулами актина.
Тропонин образует утолщение на конце каждой нити тропомиозина.
Под влиянием возникшего в мышечном волокне ПД из саркоплазматического ретикулума (СПР — депо Ca2+) высвобождаются ионы Ca. Кальций связывается с тропонином, который смещает тропомиозиновый стержень, что приводит к открытию актиновых центров.

В результате, к актиновым центрам присоединяются головки поперечных миозиновых мостиков.
Процесс расслабления происходит в обратной последовательности с использованием энергии АТФ за счет функционирования кальциевого насоса.
При отсутствии повторного импульса ионы Ca не поступают из СПР. В результате отсутствия Ca-тропонинового комплекса, тропомиозин возвращается на свое прежнее место, блокируя актиновые центры актина. Актиновые протофибриллы легко скользят в обратном направлении благодаря эластичности мышцы, и мышца удлиняется (расслабляется).
Гладкие мышцы
Гладкие мышцы — это мышцы, формирующие слой стенок полых внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток без поперечной исчерченности за счет хаотичного расположения миофибрилл.
Особенности гладких мышц:
- Иннервируются волокнами вегетативной нервной системы (ВНС);
- Обладают низкой возбудимостью:
- Обладают низкой величиной МП (мембранного потенциала) — -50 — -60 мВ из-за более высокой проницаемости для ионов Na+
- ПД (потенциал действия) отличается меньшей амплитудой и большей длительностью. Он формируется в основном за счет ионов Ca2+
- Медленная проводимость:
Клетки в гладких мышцах функционально связаны между собой посредством щелевидных контактов — нексусов, которые имеют низкое электрическое сопротивление. За счет этих контактов ПД распространяется с одного мышечного волокна на другое, охватывая большие мышечные пласты, и в реакцию вовлекается вся мышца.
Гладкие мышцы способны осуществлять относительно медленные ритмические и длительные тонические сокращения.
Медленные ритмические сокращения обеспечивают перемещение содержимого органа из одной области в другую.
Длительные тонические сокращения, особенно сфинктеров полых органов, препятствуют выходу из них содержимого.
Это способность сохранять приданную им при растяжении или деформации форму. Благодаря пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии.
Особенность гладких мышц, отличающая их от скелетных. Благодаря автоматии гладкие мышцы могут сокращаться в условиях отсутствия иннервации . Важную роль в этом играет растяжение.
Растяжение является адекватным раздражителем для гладкой мускулатуры. Сильное и резкое растяжение гладких мышц вызывает их сокращение.
Сравнительная характеристика скелетных и гладких мышц:
Читайте также:
