
Низкая лабильность нервный центр
Нервный центр это совокупность нейронов, расположенных в различных отделах ЦНС и объединенных выполнением одной функции.
Например: центр дыхания, пищеварения и т. д. Нервные центры имеют многоуровневую организацию. Например, регуляция дыхания осуществляется с участием центров спинного мозга, продолговатого, моста, коры. Нейроны в нервном центре связаны синаптически и образуют нейронные сети. Процессы, происходящие в нейронных сетях, обеспечивают определенный уровень активности нервного центра путем:
1) регуляция входа информации;
2) трансформации ритма;
3) ослабления и усиления информации;
4) за счет процессов в нейронных сетях возникает низкая лабильность, быстрая утомляемость и высокая чувствительность к кислороду нервных центров.
Регуляция ввода информации осуществляется благодаря наличию нейронных сетей с конвергенцией и дивергенцией.
Конвергенция – это процесс схождения импульсов по многим афферентным путям на одном нейроне. Так, на мотонейроне сходятся сигналы от афферентных волокон, от различных нисходящих трактов, сходятся аксоны от возбуждающих и тормозных нейронов. Благодаря конвергенции на нейроне происходят процессы пространственной суммации.
Механизм пространственной суммации. На нейроне суммируются ВПСП и ТПСП, возникающие в различных синапсах. Если преобладает активность возбуждающих синапсов и суммарная величина ВПСП будет достаточной для возникновения ПД, то нейрон будет в возбужденном состоянии. Если преобладает активность тормозных синапсов и суммарные тормозные потенциалы подавляют активность возбуждающих синапсов или снижают величину суммарного возбуждающего потенциала, то нейрон заторможен.
Временная суммация. Этот процесс не связан с конвергенцией и заключается в суммировании ВПСП и ТПСП, возникающих в одном синапсе. Поэтому частые, но слабые сигналы, суммируясь, могут вызывать рефлекторный ответ или наоборот, затормозить его.
Роль конвергенции в деятельности нервного центра. Благодаря тому, что некоторые нейроны могут оказаться общими для различных рефлекторных дуг возникает явление окклюзии. Суть явления заключается в том, что рефлекторный ответ, возникающий при одновременном раздражении двух рецептивных полей оказывается меньше суммы рефлекторных ответов при раздельном раздражении этих же рецептивных полей.
Благодаря конвергенции возникает, и облегчение рефлекторной деятельности при одновременном раздражении различных рецептивных полей.
Вследствие конвергенции возбуждающих и тормозных путей на нейронах нервного центра рефлекторный ответ может быть заторможен при активации другого рецептивного поля. Дивергенция – это способность нейрона устанавливать, многочисленные связи с другими нейронами.
Благодаря процессу дивергенции одна и та же информация может поступать в различные нервные центры, что обеспечивает иррадиацию возбуждения и торможения в ЦНС. В нормальных условиях иррадиации возбуждения препятствует деятельность тормозных нейронов.
Трансформация ритма заключается в том, что информация, выходящая из нервного центра отличается по частоте и ритму от приходящей к нему афферентной информации. Возможно как учащение, так и урежение импульсации.
Ослабление сигналов. Такое явление может происходить при длительной работе нервного центра. В его синапсах развивается синаптическая депрессия. Проявляется в снижении постсинаптических потенциалов и связана со стойкой деполяризацией постсинаптической мембраны при длительной работе синапса. Возможно это нейронный коррелят привыкания нервных центров.
Усиление сигналов осуществляется двумя путями:
1) путем посттетанической потенциации. Ответ на слабый сигнал усиливается, если этот сигнал поступает после предварительного ритмического раздражения. Механизм этого явления заключается в том, что ритмическое раздражение привело к накоплению ионов кальция в пресинаптическом окончании. В результате этого слабый сигнал вызвал увеличенное выделение медиатора и большую величину ВПСП на нейроне.
2) второй механизм, усиливающий поступающий сигнал – реверберация.
Реверберация – это циркуляция импульсов по замкнутым нейронным сетям. На этом механизме основана кратковременная память, обучение.
Низкая лабильность нервных центров.
Лабильность – максимальное количество импульсов, которое ткань может генерировать в единицу времени синхронно с раздражением. Чувствительные нервы до 1000 имп./сек., двигательные нервы до сотни импульсов. Таким образом, нервный центр имеют самый низкую лабильность. Связано это с функциональными возможностями центральных синапсов. Утомляемость нервных центров проявляется в постепенном снижении и прекращении рефлекторного ответа при продолжительном раздражении афферентных центров. Нервные центры имеют самую высокую утомляемость.
Механизм легкой утомляемости нервных центров связан с синаптическими процессами: снижением количества легко доступного медиатора, снижением чувствительности постсинаптической мембраны к медиатору, снижением активности энзимов синапса.
Нервные центры характеризуются высокой чувствительностью к дефициту кислорода, что связано с высокой интенсивностью обменных процессов.
Дата добавления: 2016-09-20 ; просмотров: 626 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Интеграция нервных процессов и рефлекторная деятельность ЦНС, лежащие в основе приспособительных реакций организма, во многом определяется общими свойствами нервных центров:
односторонним проведением возбуждения,
замедленным проведением возбуждения,
способностью к иррадиации,
способностью к суммации,
способность к тонической активности,
повышенной чувствительностью к недостатку питательных веществ и кислорода.
Одностороннее проведение возбуждения – это способность нервных центров проводить возбуждение только в одном направлении - от афферентов к эфферентам.
Если раздражать электрическим током афферент, то в эфферентных нервных волокнах возникает серия ПД. Однако, при раздражении эфферента, в афферентных волокнах возбуждение не возникает. Одностороннее проведение сигналов обусловлено возможностью передачи возбуждения в центральных химических синапсах только от пресинаптической мембраны к постсинаптической.
Схема эксперимента, доказывающего одностороннее проведение возбуждения в нервном центре

Замедленное проведение возбуждения - это свойство нервных центров, передавать возбуждение с низкой скоростью.
Между моментом поступления нервного импульса к окончанию аксона и формированием ВПСП на постсинаптической мембране проходит около 0,5 мс. Время от поступления нервного импульса к окончанию аксона до формирования ВПСП называется синаптической задержкой.
Низкая лабильность - это свойство нервных центров, характеризующееся относительно небольшой частотой импульсации, которую они способны воспроизвести без искажения ритма стимуляции афферента.
Лабильность нервных центров не превышает 50-70 ПД/с. Это обусловлено синаптической задержкой передачи возбуждения в центральных химических синапсах.
Повышенная утомляемость - это свойство нервных центров, которое проявляется в постепенном снижении или полном прекращении рефлекторного ответа при продолжительном раздражении афферентных волокон.
Снижение работоспособности нервных центров связано с нарушением передачи возбуждения в межнейронных химических синапсах и обусловлено повышенной утомляемостью центральных химических синапсов.
Иррадиация – это свойство нервного центра передавать возбуждение в ЦНС на другие нервные центры при достаточно сильном воздействии раздражителя на рефлексогенную зону.
Благодаря распространению возбуждения на несколько нервных центров рефлекторная реакция приобретает генерализованный характер. Иррадиации препятствуют процессы торможения, которые в нормальных условиях ограничивают распространение возбуждения в ЦНС.
При заболеваниях зубов афферентная импульсация от них существенно возрастает, что приводит к генерализованному распространению возбуждения. Иррадиация возбуждения при интенсивной зубной боли (например, при пульпите), приводит к тому, что пациент не в состоянии точно указать больной зуб.
В основе иррадиации возбуждения в ЦНС лежит морфологический принцип дивергенции (дихотомического ответвления аксонов нейронов), который заключается в способности клеток нервного центра устанавливать многочисленные синаптические контакты с нейронами других нервных центров.
Схема иррадиации возбуждения в ЦНС

1) афферент, 2) коллатерали, 3) вставочные нейроны, 4) эфферентные нейроны.
Способность к суммации - это свойство нервных центров формировать рефлекторную реакцию при действии на рецепторы раздражителей, которые являются подпороговыми для нервного центра.
Различают два вида суммации: последовательную (временную) и одновременную (пространственную). Последовательная суммация наблюдается при взаимодействии возбуждений, которые поступают в нервный центр с коротким интервалом времени друг за другом по одним и тем же афферентам.
Если на рецептор действует один пороговый для рецептора и афферента, но подпороговый для возбуждения нервного центра раздражитель, то разрядная деятельность в области аксонного холмика центрального нейрона, а значит и рефлекторный ответ, не возникают. Однако, при последовательном действии серии таких раздражителей, в центральном синапсе увеличивается выделение медиатора. Вследствие этого на постсинаптической мембране происходит суммация ВПСП и начинается циркуляция местных ионных токов, сила которых достаточна для генерации в области аксонного холмика серии эфферентных ПД, а значит и для возникновения ответной рефлекторной реакции эффектора.
Примером проявления подобной суммации может служить чихание, которое возникает рефлекторно в результате длительной стимуляции рецепторов слизистой оболочки носа, накопившейся слизью, частицами пыли или другими раздражающими факторами.
Пространственная суммация возникает при взаимодействии возбуждений, одновременно приходящих в нервный центр по нескольким афферентам. При одновременном действии раздражителя на несколько рецепторов одного рецептивного поля, сигналы от которых конвергирует к одному нейрону, увеличивается выделение медиатора в центральных синапсах. Вследствие этого происходит суммация ВПСП на постсинаптической мембране, и начинают циркулировать местные ионные токи, сила которых достаточна для генерации серии эфферентных ПД в области аксонного холмика, что приводит к рефлекторному ответу исполнительного органа.
Последействие - это способность нервного центра продлевать рефлекторный ответ после прекращения раздражения рецепторов.
Известны два основных механизма последействия: кратковременный и долговременный. Кратковременное рефлекторное последействие связано со следовой деполяризацией постсинаптической мембраны нейрона после прекращения раздражения афферента. После прекращения раздражения рецепторов на постсинаптической мембране центрального синапса сохраняется следовая деполяризация в виде ВПСП, который поддерживает высокую возбудимость эфферентного нейрона. В это время за счет теплового движения молекул в синаптическую щель может самопроизвольно выделиться нескольких квантов медиатора, что приводит к образованию миниатюрного ВПСП, который суммируется со следовым потенциалом. В результате этого продолжается циркуляция местных ионных токов и поддерживается разрядная деятельность эфферентного нейрона, а следовательно и рефлекторная реакция.
За счет кратковременного механизма разрядная деятельность нейрона после прекращения раздражения афферента продлевается на несколько десятков мс. По мере уменьшения амплитуды ВПСП импульсная активность нейрона постепенно уменьшается, а затем прекращается совсем.
Долговременное рефлекторное последействие обусловлено реверберацией - циркуляцией возбуждения по замкнутым нейронным цепям в нервных центрах.
Схема организации нейронной ловушки

1) афферент, 2) эфферентный нейрон, 3) коллатераль, 4) вставочные нейроны.
Трансформация ритма - это свойство нервных центров изменять частоту проходящих через них импульсов.
Частота импульсации при прохождении через нервный центр может увеличиваться или уменьшаться. Способность нервных центров увеличивать частоту проходящих через них импульсов, называют мультипликацией ритма. Выделяют два основных механизма мультипликации: кратковременный и долговременный.
Кратковременная мультипликация, как и последействие, связана с суммацией после одиночного стимула на постсинаптической мембране миниатюрного и следового ВПСП.
Долговременная мультипликация обусловлена распространением возбуждения по дихотомически разветвляющимся коллатералям аксонов афферентов и вставочных нейронов, которые затем конвергируют (сходятся) к одному эфферентному нейрону. Вследствие схождения к эфференту нескольких вставочных нейронов, которые связаны с одним афферентом, нервный центр на одиночное раздражение афферента отвечает серией эфферентных ПД.
Схема нейронной организации долговременной мультипликации

1) афферент, 2) коллатерали, 3) вставочные нейроны, 4) эфферентный нейрон.
Пластичность - это способность нервных центров изменять свое функциональное назначение в зависимости от особенностей условий деятельности.
Пластичность обеспечивает изменение эффективности и направленности связей между нервными клетками. Она является рабочим механизмом обучения. Наибольшей пластичностью обладает кора больших полушарий головного мозга. При повреждении какой-либо зоны коры через некоторое время другие ее участки могут брать на себя выполнение функции утраченного отдела.
Способность к тонической активности заключается в постоянной минимальной импульсной активности нервных центров, которая поддерживается за счет афферентных сигналов от рефлексогенных зон. Тоническая активность нервных центров обеспечивает тонус большинства исполнительных органов и их постоянную готовность к деятельности даже в состоянии относительного покоя.
Повышенная чувствительность к недостатку питательных веществ и кислорода обусловлена высоким уровнем метаболизма нервной ткани. Специфичность обмена веществ в нейронах состоит в исключительности использования в качестве питательного материала глюкозы, тогда как другие клетки используют также различные моносахариды, жирные кислоты и аминокислоты. Кроме того, в нейронах преобладает окислительное фосфорилирование, требующее непрерывной доставки О2.
При относительно небольшой массе, составляющей не более 2% от веса тела, мозг человека потребляет до 20% всего поступившего в организм кислорода и 17% глюкозы. Интенсивность потребления глюкозы настолько велика, что в нейронах не успевают образоваться запасы гликогена. Поэтому уменьшение доставки глюкозы к ЦНС быстро влечет за собой нарушение функции нервных центров.
Интенсивность потребления нервными клетками кислорода составляет около 50 мл в мин. Потребляя такое большое количество О2, мозг и особенно кора больших полушарий высокочувствительны к его недостатку. Поэтому уже через 5-7 секунд после прекращения кровообращения в мозге человек теряет сознание, а через 5-6 минут нервные клетки больших полушарий подвергаются необратимым биологическим изменениям.
Центры ствола мозга менее чувствительны к недостатку кислорода: их функция может восстановиться после 20 минут полного прекращения кровообращения. Центры спинного мозга еще более выносливы. Их функция восстанавливается даже через 30 минут после полного прекращения притока крови.
При гипотермии - понижении температуры тела, вследствие снижения уровня обмена веществ, ЦНС дольше переносит недостаток О2 и питательных веществ. Гипотермия при температуре тела 25-28 0 С повышает устойчивость коры больших полушарий к кислородной недостаточности с 5 до 15-20 минут.
1. Односторонняя передача возбуждения в синапсе ( от пре к постсинаптической мембране ). В обратном направлении передача невозможна, ибо медиатор содержится только в пресинаптических бляшках, а рецептор к нему только на постсинаптической мембране.
2. Синаптическая задержка. В синапсе передача возбуждения задерживается на 0,6 – 0,8 сигм ( 1 сигма=0,001 сек.). Время уходит на выделение медиатора, транспорт его через синаптическую щель, контакт с рецептором и суммацию ВПСП.
3. Суммация ВПСП.
А. Последовательная, когда последователь по времени суммируются ВПСП.
Б. Пространственная (одновременная). На одном нейроне может контактировать несколько аксонов. Из каждого одновременно выделится по кванту АХ, которые сразу вызовут падение мембранного потенциала в нейроне до критического уровня (Рис.30).
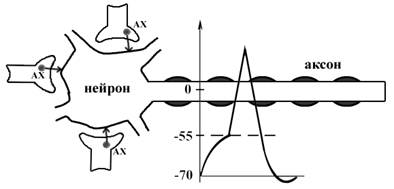
Рис. 30. Схема одновременной суммации ВПСП.
4. Высокая утомляемость нервного центра.
При длительном действии раздражителя расходуется АХ в пресинаптической области и не возбуждается постсинаптическая мембрана. Восстанавливается работоспособность утомлённого нервного центра за счёт отдыха. Он может быть активным и пассивным. Пассивный отдых. Когда ничего не делая ожидается время синтеза достаточного количества АХ в пресинаптической бляшке. Активный отдых. Для этого необходимо заняться другим видом работы и подключить к возбуждению параллельную рефлекторную дугу. От неё по коллатерали возбуждение подойдёт к утомлённому нервному центру и выбросит в синапс недостающее количество АХ. ВПСП одновременно будет суммироваться на нейроне из нескольких синапсов. Этот вид отдыха более продуктивен, восстановление трудоспособности наступает быстрее, чем при пассивном отдыхе (Рис.31).
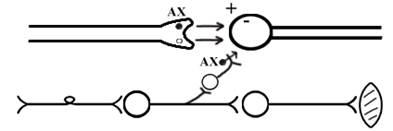
Рис. 31. Механизм активного отдыха.
5. ТРАНСФОРМАЦИЯ ритма. Нервный центр может изменять число нервных импульсов, подошедших к нему в область пресинаптичеческих бляшек, увеличивая или уменьшая частоту (трасформируя). Поэтому трансформация может быть как поышающая, так и понижающая. ПОНИЖАЮЩАЯ – возникает за счёт суммации ВПСП (Рис.32).
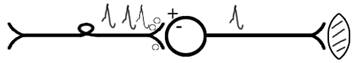
Рис.32. Механизм понижающей трансформации.
ПОВЫШАЮЩАЯ трансформация за счёт возвратных коллатералей, исходящих от нервного центра. Из окончаний собственных коллатералей выделяется АХ, который вызовет дополнительные импульсы возбуждения в нервном центре ( Рис.33).
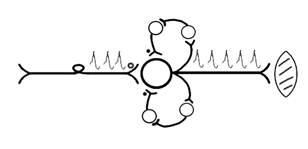
Рис. 33. Механизм повышающей трансформации в нервном центре.
6. ПОСЛЕДЕЙСТВИЕ – способность нервного центра генерировать возбуждение после действия раздражителя. Это может быть связано с действием гуморальных раздражителей, способных вызывать деполяризацию или с наличием возвратных коллатералей. По ним импульсы возвращаются к нейрону и его возбуждают (Рис.34).
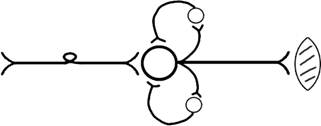
Рис.34. Механизм последействия в нервном центре.
7. ВЫСОКАЯ ЧУВСТВИТЕЛЬНОСТЬ нервного центра к биологически активным веществам (БАВ). В крови существуют соединения, способные увеличивать проницаемость мембраны нейрона к ионам натрия, вызывая деполяризацию. При небольшой их концентрации возникает частичная деполяризация, что определяет тонус нейрона и его готовность к ответной реакции. Это важно для поддержания гомеостаза в организме. При высокой концентрации БАВ в крови может возникнуть самовозбуждение нейронов без действия раздражителей.
Нервный центр обладает высокой чувствительностью к недостатку кислорода. Нейроны коры головного мозга способны существовать без кислорода не более 3-5 минут и этим определяется длительность клинической смерти. Нейроны нижележащих отделов ЦНС могут существовать без кислорода несколько дольше.
1. НИЗКАЯ ЛАБИЛЬНОСТЬ нервного центра. Лабильность – это функциональная активность. К нервному центру подходит до 500 импульсов, а он может пропустить 100-120 импульсов. Это связано с последовательной суммацией ВПСП, когда частота импульсов теряется. Низкая лабильность предохраняет рабочий орган от перегрузок.
ТЕМА 7: Торможение в центральной нервной системе.
Варианты организации тормозных процессов в ЦНС.
1. Реципрокное торможение (от лат. reciprocus - взаимный) было открыто английским физиологом Ч. Шеррингтоном и российским физиологом Н.Е. Введенским. Этот вид торможения основан на том, что одни и те же афферентные пути, через которые осуществляется возбуждение одной группы нервных клеток, обеспечивают через вставочные нейроны торможение других групп нейронов. Например, при возбуждении болевых рецепторов кожи конечности сигнал от ноцицепторов с участием афферентного нейрона поступает в спинной мозг, где переключается на альфа-мотонейрон мышц-сгибателей и одновременно на тормозной нейрон, который тормозит активность альфа-мотонейрона мышц-разгибателей. Реципрокное торможение характерно как для спинного мозга, так и для головного. Экспериментально доказано, что нормальное выполнение каждого естественного двигательного акта основано на использовании реципрокного торможения.
2. Возвратное, или антидромное, торможение наблюдается в отношении альфа-мотонейронов спинного мозга. При возбуждении альфа-мотонейрона нервный импульс направляется к мышечным волокнам, возбуждая их. Одновременно по коллатерали, идущей к тормозному нейрону (клетка Реншоу), импульс возбуждает эту тормозную клетку, которая в свою очередь вызывает торможение возбужденного ранее альфа-мотонейрона. Таким образом, альфа-мотонейрон, активируясь, через систему тормозного нейрона сам себя (возвратно, или антидромно) затормаживает. Чем выше активность альфа-мотонейрона, тем выраженнее тормозное влияние клеток Реншоу на этот нейрон (это проявляется в снижении частоты генерации потенциалов действия альфа-мотонейроном).
4. Тоническое торможение отражает наличие постоянного тормозного влияния одной структуры на другую. Примером такого постоянного тормозного влияния являются тормозные нейроны коры больших полушарий, которые угнетают нейроны ретикулярной формации ствола мозга, нейроны таламуса и лимбической системы.
5. Общее центральное торможение открыто в 1937 И.С. Беритовым (Словарь физиологических терминов, 1987). Это нервный процесс, который развивается при любой рефлекторной деятельности. Он захватывает почти всю ЦНС, включая центры головного мозга. Такое торможение проявляется раньше, чем возникнет какая-либо двигательная реакция. Оказалось, что общее центральное торможение может проявляться при такой малой силе раздражителя, при которой двигательный эффект еще отсутствует. По мнению И.С. Беритова, такое торможение обеспечивает концентрацию возбуждения в определенных группах вставочных и двигательных нейронов, препятствуя возникновению других рефлекторных или поведенческих актов, которые могли бы возникнуть под влиянием раздражителей. Важную роль в формировании общего центрального торможения, вероятно, играет желатинозная субстанция спинного мозга. Не исключено, что общее
центральное торможение играет важную роль в создании целостной поведенческой деятельности, а также в обеспечении избирательного возбуждения определенных рабочих органов.
Пессимальное торможение как вариант вторичного торможения. Этот вид торможения развивается в возбуждающих синапсах в результате сильной и длительной деполяризации постсинаптической мембраны приходящими сюда высокочастотными импульсными потоками (подобно пессимуму Введенского в нервно-мышечном синапсе либо католической депрессии Вериго), что приводит к аккомодационным изменениям и снижению возбудимости (повышению порога возбуждения) в постсинаптической мембране, делающим ее неактивной. В частности, этот вид торможения лежит в основе пессимального торможения условнорефлекторной деятельности в коре больших полушарий.
Торможение вслед за возбуждением как вариант вторичного торможения. Этот вид торможения развивается достаточно часто, так как возникает всякий раз на фоне следовой гиперполяризации мембраны нейрона после очередного его возбуждения. Для него характерен сравнительно кратковременный период существования, так как он определяется лабильностью нейрона, т.е. скоростью восстановления исходного уровня мембранного потенциала после генерации очередного потенциала действия.
Нервный центр - совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт.
Известно, что группы клеток, регулирующих одну и ту же функцию, могут располагаться в разных отделах ЦНС. При этом различно функциональное значение не только нейронов, лежащих в разных отделах ЦНС, но и нейронов одного и того же отдела. И. П. Павлов ввел понятие ядра и рассеянных элементов центра. При повреждении ядра центра происходит глубокое нарушение функции, а нарушение функции, возникающее при повреждении рассеянных элементов (нейронов расположенных к периферии от ядра), легко компенсируется.
Расположение клеток одного и того же центра в разных отделах ЦНС ярко прослеживается на центре речи. Эту функцию обеспечивают нервные клетки, регулирующие двигательную активность речевых мышц языка и губ, мышц гортани, расположенные в продолговатом мозге. Высшая регуляция речевой функции осуществляется клетками, лежащими в височной, лобной и теменной областях коры больших полушарий. При повреждении теменной области человек теряет способность понимать речь, хотя и хорошо ее слышит. При повреждении лобных долей - нарушается двигательная речевая функция.
Кроме того, одни и те же нейроны ЦНС могут участвовать в регуляции разных функций. Так, клетки центра глотания, расположенные в продолговатом мозге, обеспечивают последовательные сокращения мышц языка, неба, глотки, гортани и пищевода. Но поднятие мягкого неба происходит и при акте рвоты. Следовательно, нервные клетки, регулирующие движение мышц мягкого неба входят в состав и центра глотания и центра рвоты.
Для нормального осуществления определенной функции необходима целостность всех отделов ЦНС, регулирующих эту функцию.
В тех случаях, когда говорят о свойствах нервных центров, имеют в виду не такое широкое его определение, а лишь свойства нейронов, связанные с особенностями их строения и механизмом передачи возбуждения в синапсах.
Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований.
1. Одностороннее проведение возбуждения. В ЦНС - в ее нервных центрах, внутри рефлекторной дуги и нейронных цепей возбуждение, как правило, идет в одном направлении - от пресинаптической мембраны к постсинаптической, т.е. вдоль рефлекторной дуги от афферентного нейрона к эфферентному. Это связано со свойствами синапсов: для химических - с выработкой медиаторов в пресинаптической части синапса, диффузией их через синаптическую щель к хеморецепторам постсинаптической мембраны, а для большинства электрических синапсов - с полупроводниковыми свойствами их синаптических мембран. В целом это организует деятельность ЦНС и является одним из принципов координационной деятельности ЦНС.
2. Замедление проведения возбуждения в нервных центрах, или центральная задержка. Замедление проведения возбуждения по нервным центрам получило название центральной задержки. Она обусловлена медленным проведением нервных импульсов через синапсы, так как затрачивается время на следующие процессы: выделение медиатора из пресинаптических везикул, трансфузия его через синаптическую щель к постсинаптической мембране и генерация возбуждающего постсинаптического потенциала (ВПСП). При этом истинная синаптическая задержка (до начала генерации ВПСП) составляет 0,5 мс, а вместе со временем генерации ВПСП она достигает 1,5-2,5 мс.
3. Суммация возбуждения и суммация торможения. Принято выделять два вида суммации - временную и пространственную.
Временная, или последовательная, суммация проявляется в том, что в области постсинаптической мембраны происходит суммация следов возбуждения во времени, т.е. на нейроне в области его аксонного холмика происходит интеграция событий, разыгрывающихся на отдельных участках мембраны нейрона на определенном отрезке времени. Например, если с определенным интервалом к нейрону 1 в точку А через возбуждающие синапсы приходят потенциалы действия (ПД) от возбуждающего нейрона 2, то на постсинаптической мембране нейрона 1 будут генерироваться ВПСП. Если эти ВПСП не достигают критического уровня деполяризации, то ПД на аксонном холмике не возникает. Если же частоту следования подпороговых импульсов от нейрона 1 увеличить, то на аксонном холмике может произойти суммация ВПСП, при которой суммарное изменение мембранного потенциала достигнет критического уровня деполяризации и нейрон 1 будет генерировать ПД, т.е. возбудится. Это явление носит название временной, или последовательной, суммации, так как в этом случае происходит суммация следов возбуждения во времени.
Пространственная суммация возбуждения проявляется в суммировании на аксонном холмике нейрона 1 постсинаптических потенциалов, которые возникают одновременно в различных точках этого нейрона (А, В.С и т.д.) в ответ на приходящие от нейронов 2,3, 4 и т.д. потенциалы действия. Даже если каждый в отдельности из нейронов 2, 3, 4 и т.д. вызывает лишь подпороговые ВПСП, при синхроннном их появлении они будут способны довести мембранный потенциал в области аксонного холмика нейрона до критического уровня деполяризации и тем самым вызывать возбуждение нейрона 1.
В ЦНС имеет место сочетание двух видов суммации возбуждения (временной и пространственной). Все сказанное в полной мере относится и к явлению суммации торможения - одновременная генерация тормозных по-стинаптических потенциалов в отдельных точках нейрона, где локализованы тормозные синапсы, либо последовательное увеличение величины ТПСП в одной точке нейрона может вызывать выраженное повышение мембранного потенциала в области аксонного холмика и тем самым снизить его возбудимость (явление пространственной и временной суммации торможения). Учитывая, что как правило нейроны ЦНС имеют огромное число синаптических входов (до 10 000), в том числе возбуждающих и тормозных, то можно утверждать, что временная и пространственная суммация возбуждающих и тормозных постсинаптических потенциалов является тем важнейшим процессом, который в конечном итоге определяет состояние нейрона (покой, возбуждение, торможение).
4. Явление окклюзии (или закупорки). Это явление было открыто Ч. Шеррингтоном. Оно отражает эффект взаимодействия между собой двух импульсных потоков, при котором имеет место взаимное угнетение рефлекторных реакций - суммарная ответная реакция (рефлекс), вызываемая одновременным воздействием двух потоков, меньше, чем сумма двух реакций, возникающих при действии каждого из этих двух потоков в отдельности. Согласно Ч. Шеррингтону, явление окклюзии объясняется перекрытием синаптических полей, образуемых афферентными звеньями двух взаимодействующих рефлексов. В связи с этим при одновременном поступлении двух афферентных посылок ВПСП вызывается каждым из них отчасти в одних и тех же нейронах. В целом, Ч. Шеррингтон считал, что явление окклюзии, или закупорки, отражает характерный для ЦНС принцип конвергенции - схождения афферентных путей на одном теле эфферентного нейрона. Явление окклюзии используют в физиологических экспериментах для определения общего звена для двух путей распространения импульсов. Если имеется общее звено, то одновременное раздражение двух путей с максимальной интенсивностью вызывает ответ меньшей интенсивности, чем сумма ответов, получаемых при раздельном раздражении этих путей стимулами той же интенсивности.
5. Явление облегчения. Это явление, которое по своему внешнему проявлению противоположно окклюзии. Оно проявляется в том, что при совместном раздражении рецептивных полей двух рефлексов наблюдается усиление реакций организма на действие двух раздражителей одновременно. Иначе говоря, суммарная реакция выше суммы реакции при изолированном раздражении каждого из этих рецептивных полей. Явление облегчения объясняется тем, что часть общих для обоих рефлексов нейронов возбуждается лишь при совместном действии двух потоков импульсов, в то время как при изолированном действии эти нейроны в силу низкой возбудимости, не активируются. Подобно явлению окклюзии, явление облегчения демонстрирует наличие конвергентных процессов в ЦНС.
6. Трансформация ритма возбуждения. Это одно из свойств нейрона как компонента нейронной цепи, которое обнаруживается в процессе проведения возбуждения по нейронным цепям. Трансформация ритма возбуждения заключается в способности нейрона изменять ритм приходящих импульсов. Особенно четко проявляется свойство трансформации ритма при раздражении афферентного волокна одиночными импульсами. На такой импульс нейрон отвечает пачкой импульсов. Трансформация ритма возбуждения может происходить 1) за счет возникновения длительного ВПСП, на фоне которого генерируется подряд несколько спайков (подобное явление характерно, например, для тормозных клеток Реншоу); 2) за счет следовых колебаний мембранного потенциала, которые могут возникнуть в ответ на приходящий импульс - если величина этих колебаний всякий раз достигает критического уровня деполяризации, то каждому такому колебанию будет соответствовать возникновение вторичного ПД.
Трансформация ритма возбуждения проявляется и в противоположном феномене - частота приходящих к нейрону импульсов выше, чем частота генерации ПД при ответе нейрона на эти импульсы. В этом случае урежение импульсации связано с более низкой лабильностью нейрона-приемника, которая, в свою очередь обусловлена большой длительностью фазы следовой гиперполяризации этого нейрона.
Высокая утомляемость нервных центров коррелирует с характерной для нейрона и, особенно, для нейронных объединений низкой лабильностью. Если нервное волокно, являясь периферическим отростком нейрона, способно генерировать до 1000 потенциалов действия в 1 с, то для нейрона предельный ритм возбуждения составляет не более 50-100 Гц, а для нейронных объединений - не более 50 Гц. С одной стороны, низкая лабильность нейронов обусловлена наличием длительной следовой гиперполяризации, а с другой особенностями синаптической передачи.
Помимо высокой утомляемости для нервных центров характерна также высокая чувствительность к гипоксиии, т.е. низкому содержанию кислорода и к ряду нейротропных веществ: нервным ядам, наркотикам, алкоголю, ганг-лиоблокаторам, антидепрессантам, психостимуляторам, транквилизаторам. Все эти факторы по тем или иным причинам существенно нарушают деятельность отдельных нейронов, входящих в нейронное объединение (нервный центр), а также нарушают деятельность нейронного объединения в целом. В процессе эволюции были выработаны механизмы защиты, позволяющие создать стабильную среду для деятельности нейронов. Одним из них является гематоэнцефалический барьер, строго регулирующий транспорт различных веществ из крови в мозг. Однако возможности этого барьера небезграничны.
9. Тонус нервных центров. Для многих нейронных объединений, или нервных центров, характерна фоновая активность, т.е. генерация нервных импульсов с определенной частотой на протяжении длительного времени. Такая активность обусловлена не наличием в составе данного объединения нейрона-пейсмкера (фоновоактивного нейрона), а постоянным возбуждением афферентного нейрона благодаря непрерывному раздражению сенсорных рецепторов. Например, тонус двигательных центров поддерживается непрерывным потоком импульсов от проприорецепторов - чувствительных нервных окончаний, заложенных в самих мышцах. Слабое возбуждение от центров по эфферентным волокнам передается мышцам, которые всегда находятся в состоянии некоторого сокращения. Перерезка афферентных или эфферентных волокон приводит к потере мышечного тонуса. Тоническая активность характерна для многих центров, в том числе для центров, регулирующих деятельность сердца и сосудов. В целом, тонус нервных центров обеспечивает постоянную (но разной интенсивности в зависимости от состояния на периферии) импульсацию к соответствующим периферическим системам, а также постоянное межцентральное взаимодействие.
Существуют синаптические, мембранные, молекулярные и морфологические механизмы пластичности. Это означает, что в основе пластичности нейронных объединений (нервных центров) могут лежать изменения на уровне одного элемента, входящего в объединение (например, в целом, нейрона, или его отдельной области), либо одновременно во многих его элементах. Основным фундаментом, позволяющим реализовать свойство пластичности, очевидно, следует считать наличие у каждого нейрона в
отдельности огромного числа синаптических связей, а также возможность изменения синтетических процессов внутри каждого нейрона.
Экспериментальной моделью пластичности нейронных объединений является постетаническая потенциация, т.е. длительное повышение возбудимости нейрона после его кратковременной высокочастотной (30-50 Гц в течение 1-2 с) стимуляции.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Читайте также:
