
Норадреналин и парасимпатическая нервная система
Вегетативная нервная система поддерживает гомеостаз. ВНС управляет такими висцеральными функциями, как кровообращение, пищеварение и выделение, главным образом без условного или сознательного контроля. ВНС также модулирует функцию эндокринных желез, регулирующих метаболизм. ВНС имеет сенсорные и моторные компоненты и разделяется на симпатическую и парасимпатическую системы. Первые нейроны симпатической системы расположены в промежуточных рогах тораколюмбального отдела спинного мозга; синапс со вторым набором нейронов находится в параили превертебральном симпатическом ганглии. В парасимпатической системе первые нейроны расположены либо в черепно-мозговом нерве, в автономных ядрах, либо в промежуточном роге сакрального отдела спинного мозга; синапс со вторым набором нейронов находится или в автономном ганглии (в случае черепно-мозговых нервов), или в эффекторной ткани непосредственно. ВНС имеет три главных компонента:
• афферентный (центростремительный, чувствительный);
• центральный объединяющий;
• эфферентный.
Афферентный компонент несет информацию от нейрональных физиологических рецепторов, расположенных в концах центростремительных нервов, к спинному мозгу и более высоким областям ЦНС. Большая часть этой информации обрабатывается в пределах гипоталамуса и других нижележащих областей мозга. После обработки соответствующий сигнал посылается от ЦНС вниз по эфферентным нервам к исполнительным органам (см. рис. 8.1, 8.9), названным так потому, что они отвечают на деятельность в ЦНС.
На основе различий анатомии и медиаторов эфферентную часть ВНС подразделяют на три системы:
• парасимпатическую (холинергическую);
• симпатическую (адренергическую);
• неадренергическую нехолинергическую (НАНХ).
Ацетилхолин — нейромедиатор холинергической системы. Ацетилхолин — нейромедиатор, высвобождаемый из пресинаптического окончания в автономном ганглии и в окончаниях нервов в исполнительном органе. Рецепторами для ацетилхолина служат холинорецепторы, которые подразделяют на мускариновые и никотиновые.
Норадреналин — нейромедиатор адренергической системы. Другая важная составляющая ВНС — адренергическая система. До сих пор неизвестно, какой нейромедиатор использовался в этой системе первоначально — эпинефрин или норэпинефрин. Сейчас известно, что за исключением надпочечников, которые секретируют эпинефрин (адреналин), нейроме-диатором в адренергической системе является норэпинефрин.
Ацетилхолин — ганглионарный медиатор для холинергической и адренергической систем. Эфферентные нервы и для холинергической, и для адренергической систем происходят из соответствующих частей ствола мозга и спинного мозга. Эфферентные нервы образуют синапс в ганглии, расположенном вне органа, где основным нейромедиатором является АХ:
• в адренергической системе ганглии находятся в цепочке вблизи спинного мозга, известной как паравертебральная симпатическая цепочка;
• в холинергической системе ганглий обычно располагается внутри или вблизи эффекторного органа.
Несмотря на явное анатомическое различие, оба типа ганглиев используют АХ как основной ганглионарный нейромедиатор, активирующий никотиновые рецепторы.
Нейромедиаторы могут модулировать собственное высвобождение. Нейромедиаторы могут модулировать собственное высвобождение. Нейромедиаторы могут активировать пресинаптические рецепторы на нейроне, что ингибирует высвобождение самих нейромедиаторов.
Учебное видео анатомии вегетативной нервной системы (ВНС)
- Вернуться в оглавление раздела "фармакология"
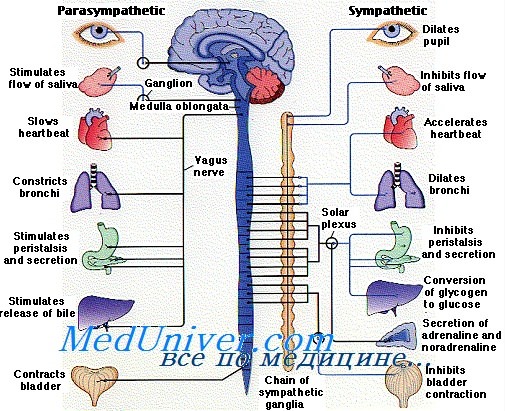
Частями вегетативной системы являются симпатическая и парасимпатическая нервная система, причем последняя оказывает непосредственное влияние и тесно взаимосвязана с работой сердечной мышцы, частотой сокращения миокарда. Локализуется она частично в головном и спинном мозге. Парасимпатическая система обеспечивает расслабление и восстановление организма после физических, эмоциональных нагрузок, однако не может существовать отдельно от симпатического отдела.
Что такое парасимпатическая нервная система
Отдел отвечает за функциональность организма без его участия. Например, парасимпатические волокна обеспечивают дыхательную функцию, регулируют сердцебиение, расширяют кровеносные сосуды, контролируют естественный процесс пищеварения и защитные функции, обеспечивают другие важные механизмы. Парасимпатическая система необходима человеку, чтобы организм расслабился после физической нагрузки. При ее участии снижается тонус мышц, приходит в норму пульс, сужается зрачок и сосудистые стенки. Это происходит без участия человека – произвольно, на уровне рефлексов
Основные центры этой автономной структуры – головной и спинной мозг, где сосредоточены нервные волокна, обеспечивающие максимально быструю передачу импульсов для работы внутренних органов, систем. С их помощью можно контролировать артериальное давление, проницаемость сосудов, сердечную деятельность, внутреннюю секрецию отдельных желез. Каждый нервный импульс отвечает за определенную часть тела, которая при его возбуждении начинает реагировать.
- Клизма от глистов
- Кольпоскопия шейки матки, что это такое
- Диетическое овсяное печенье
Все зависит от локализации характерных сплетений: если нервные волокна находятся в области таза, то отвечают за физическую активность, а в органах пищеварительной системы – за секрецию желудочного сока, перистальтику кишечника. Строение вегетативной нервной системы имеет следующие конструктивные отделы с уникальными функциями для всего организма. Это:
- гипофиз;
- гипоталамус;
- блуждающий нерв;
- эпифиз.
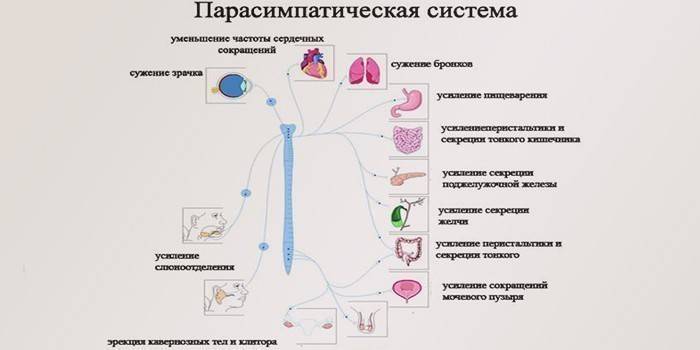
Так обозначены главные элементы парасимпатических центров, а дополнительными структурами считаются следующие:
- нервные ядра затылочной зоны;
- крестцовые ядра;
- сердечные сплетения для обеспечения толчков миокарда;
- подчревное сплетение;
- поясничное, чревные и грудные нервные сплетения.
Симпатическая и парасимпатическая нервная система
Парасимпатический отдел вегетативной нервной системы отвечает за бесперебойную работу сердечно-сосудистой, мочеполовой и пищеварительной систем, при этом имеет место парасимпатическая иннервация печени, щитовидки, почек, поджелудочной железы. Функции разные, а влияние на органический ресурс комплексное. Если симпатический отдел обеспечивает возбуждение внутренних органов, то парасимпатический – помогает восстанавливать общее состояние организма. Если возникает дисбаланс двух систем, больной нуждается в лечении.
- Потайной шов вручную
- Как ухаживать за фиалками
- Фитнес дома - видео с упражнениями для похудения. Видеоуроки домашнего фитнеса
Где расположены центры парасимпатической нервной системы
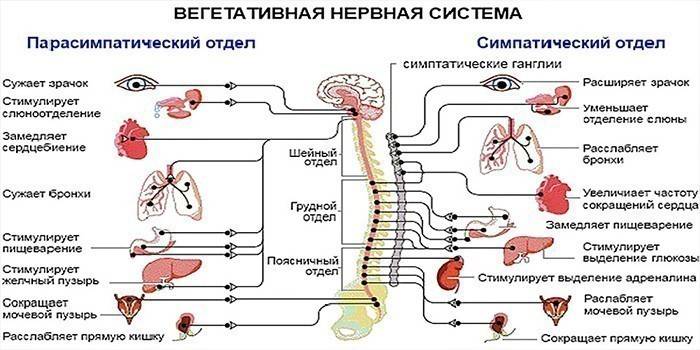
Симпатическая нервная система конструктивно представлена симпатическим стволом в два ряда узлов с обеих сторон от позвоночника. Внешне структура представлена цепочкой из нервных комочков. Если затронуть элемент так называемого расслабления, парасимпатическая часть вегетативной нервной системы локализуется в спинном и головном мозге. Итак, от центральных отделов из головного мозга импульсы, которые возникают в ядрах, идут в составе черепно-мозговых нервов, от крестцовых отделов – в составе тазовых внутренностных нервов, достигают органов малого таза.
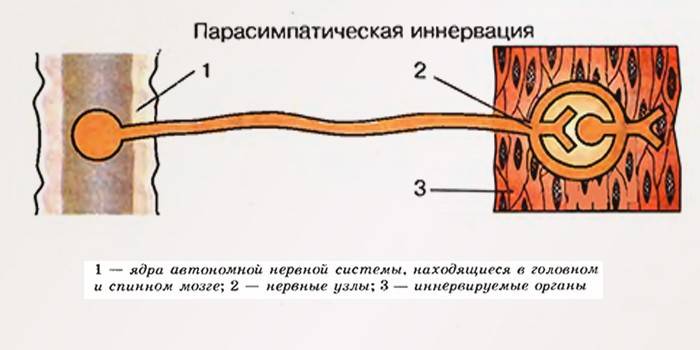
Функции парасимпатической нервной системы
Парасимпатические нервы отвечают за естественное восстановление организма, нормальное сокращение миокарда, тонус мышц и продуктивное расслабление гладких мышц. Парасимпатические волокна отличаются локальным действием, но в итоге действуют сообща – сплетениями. При локальном поражении одного из центров, страдает вегетативная нервная система в целом. Влияние на организм комплексное, а врачи выделяют следующие полезные функции:
- расслабление глазодвигательного нерва, сужение зрачка;
- нормализация кровяной циркуляции, системного кровотока;
- восстановление привычного дыхания, сужение бронхов;
- снижение артериального давления;
- контроль важного показателя глюкозы в крови;
- сокращение частоты сердечных сокращений;
- замедление прохождения нервных импульсов;
- снижение глазного давления;
- урегулирование работы желез пищеварительной системы.
Кроме того, парасимпатическая система помогает сосудам головного мозга и половых органов расширяться, а гладким мышцам прийти в тонус. С ее помощью происходит естественное очищение организма за счет таких явлений, как чиханье, кашель, рвота, походы в туалет. К тому же, если начинают проявляться симптомы артериальной гипертонии, важно понимать, что за сердечную деятельность отвечает вышеописанная нервная система. Если одна из структур – симпатическая или парасимпатическая выходят из строя, необходимо предпринимать меры, поскольку они тесно связаны между собой.
Болезни
Прежде чем использовать те или иные медицинские препараты, делать исследования, важно правильно диагностировать заболевания, связанные с нарушенной работой парасимпатической структуры головного и спинного мозга. Проблема со здоровьем проявляется стихийно, она способна поразить внутренние органы, повлиять на привычные рефлексы. В основе могут лежать следующие нарушения организма любого возраста:
- Циклический паралич. Болезнь спровоцирована цикличными спазмами, сильным повреждением глазодвигательного нерва. Заболевание возникает у пациентов разного возраста, сопровождается дегенерацией нервов.
- Синдром глазодвигательного нерва. В такой непростой ситуации зрачок может расширяться без воздействия потока света, чему предшествует повреждение афферентного участка дуги зрачкового рефлекса.
- Синдром блокового нерва. Характерный недуг проявляется у пациента незначительным косоглазием, незаметным для простого обывателя, при этом глазное яблоко направлено внутрь или вверх.
- Травмированные отводящие нервы. При патологическом процессе одновременно сочетаются в одной клинической картине косоглазие, раздвоение зрения, выраженный синдром Фовиля. Патология затрагивает не только глаза, но и лицевые нервы.
- Синдром троичного нерва. Среди основных причин патологии врачи выделяют повышенную активность болезнетворных инфекций, нарушение системного кровотока, поражение корково-ядерных путей, злокачественные опухоли, перенесенная черепно-мозговая травма.
- Синдром лицевого нерва. Наблюдается очевидный перекос лица, когда человеку произвольно приходится улыбаться, при этом испытывая болезненные ощущения. Чаще это осложнение перенесенного заболевания.
Симпатическая часть вегетативной нервной системы состоит из центрального и периферического отделов. Центральный отдел образуют клетки боковых рогов спинного мозга на уровне всех его грудных и верхних трех поясничных сегментов. Периферический отдел представлен нервными волокнами и симпатическими нервными узлами (ганглиями). Нервные узлы подразделяются на две группы:
околопозвоночные (паравертебральные), расположенные двумя цепочками по бокам от позвоночника и образующие правый и левый симпатические стволы, и предпозвоночные (превертебральные) — узлы периферических нервных сплетений, лежащие в грудной и брюшной полостях
Основным медиатором симпатических синапсов является норадреналин и такие синапсы называютадренергическими.Рецепторы, связывающие адренергический медиатор получили названиеадренорецепторов.Различают два типа адренорецепторов — альфаи бета,каждый из которых делят на два подтипа — 1 и 2. Небольшая часть симпатических синапсов использует медиатор ацетилхолин и такие синапсы называют холинергическими,а рецепторы — холинорецепторами.Холинергические синапсы симпатической нервной системы обнаружены в потовых железах. В адренергических синапсах кроме норадреналина в существенно меньших количествах содержатся адреналин и дофамин, также относящиеся к катехоламинам, поэтому медиаторное вещество в виде смеси трех соединений раньше называли симпатином.
Норадреналин (НА), из группы катехоламинов, производное аминокислоты тирозина Постганглионарные окончаниясимпатическойвегетативной нервной системы. α1- , α2-, β1-, β2-адренорецепторы (все метаботропного типа) Возбуждение α1- и β1- рецепторов оказывает возбуждающее действие, а α2- и β2-рецепторв - тормозящее.
Отсюда - набор симпатических реакций:
1) усиление сокращений сердца,
2) сужение сосудов мышц,
3) расширение бронхов,
4) ослабленние перистальтики кишечника,
5) расширение зрачков.
Обычно в одном органе содержится несколько типов адренорецепторов. При этом α-рецепторы преобладают в сосудах кожи и сфинктерах ЖКТ, β1-рецепторы - в сердце, β2-рецепторы - в бронхах, кишечнике, матке, сосудах скелетных мышц.
6. Характеристика парасимпатического отдела вегетативной нервной системы его медиаторы
Центрами парасимпатического отдела автономной нервной системы являются ядра, находящиеся в среднем мозге (IIIпара черепно-мозговых нервов), продолговатом мозге (VII, IXи Х пары черепно-мозговых нервов) и крестцовом отделе спинного мозга (ядра тазовых внутренних нервов). От среднего мозга отходят Преганглионарные волокна парасимпатических нервов, которые входят в состав глазодвигательного нерва. Из продолговатого мозга выходят Преганглионарные волокна, идущие в составе лицевого, языкоглоточного и блуждающих нервов. От крестцового отдела спинного мозга отходят Преганглионарные парасимпатические волокна, которые входят в состав тазового нерва.
При возбуждении парасимпатических нервов тормозится работа сердца отрицательные хроно-, ино-, дромо- и батмотропное действия),повышается тонус гладкой мускулатуры бронхов, в результате чего уменьшается их просвет, сужается зрачок, стимулируются процессы пищеварения (моторика и секреция), обеспечивая тем самым восстановление уровня питательных веществ в организме, происходит опорожнение желчного пузыря, мочевого пузыря, прямой кишки. Действие парасимпатической нервной системы направлено на восстановление и поддержание постоянства состава внутренней среды организма, нарушенного в результате возбуждения симпатической нервной системы. Парасимпатическая нервная система выполняет в организметрофотропную функцию.
К этому отделу относятся интрамуральные системы всех полых висцеральных органов, обладающих собственной автоматической двигательной активностью: сердце, бронхи, мочевой пузырь, пищеварительный тракт, матка, желчный пузырь и желчные пути.
Внутриорганный отдел имеет все звенья рефлекторной дуги: афферентный, вставочный и эфферентный нейроны, которые полностью находятся в органе и нервных сплетениях внутренних органов (например, ауэрбаховском и мейснеровском). Преганглионарные волокна выделяютацетилхолининорадреналин,постганглионарные —АТФиаденозин, ацетилхолин, норадреналин, серотонип, дофамин, адреналин, гистамини т.д. Главная роль принадлежит АТФ и аденозину.
Ацетилхолин (АХ), низкомолекулярное вещество с зарядом +1
Дата добавления: 2019-11-25 ; просмотров: 466 ;
Содержание
В соматической нервной системе нервные волокна тянутся к скелетной мышце, коже и органам чувств, а также от них. Соматические нервные волокна обычно реагируют на стимулы из окружающей среды, как, например, в случае рефлекса отдергивания. Большая часть активности соматической нервной системы происходит сознательно и под волевым контролем. Напротив, вегетативная, или автономная, нервная система (ВНС) в основном занята регуляцией кровообращения и деятельности внутренних органов. Она отвечает на изменяющиеся условия среды ортостатической реакцией, запуском реакций с целью регуляции внутренней среды организма. Основные виды деятельности вегетативной нервной системы не подлежат волевому контролю.
Вегетативная и соматическая нервные системы, в основном на периферии, разделены анатомически и функционально (А), но тесно контактируют в центральной нервной системе (ЦНС). Периферическая вегетативная нервная система является эфферентной (двигательной), но большинство вегетативных нервов несут также афферентные (сенсорные) нейроны. Они называются также висцеральными афферентными нейронами, потому что проводят сигналы от висцеральных (внутренних) органов, таких как пищевод, желудочно-кишечный тракт, печень, легкие, сердце, артерии и мочевой пузырь. Некоторые из них называются по нерву, который они сопровождают, например афферентный нейрон блуждающего нерва.
Активность вегетативной нервной системы обычно регулируется рефлекторной дугой, которая имеет афферентный сегмент (висцеральные и/или соматические афферентные нейроны) и эфферентный сегмент (автономные и/или соматические афферентные нейроны). Афферентные волокна проводят стимулы от кожи (болевые, или ноцицептивные, стимулы) и болевых рецепторов, от механорецепторов и хеморецепторов в таких органах, как легкие, желудочно-кишечный тракт, желчный пузырь, сосудистая система и гениталии. Вегетативная система содержит автономные эфферентные волокна, которые проводят рефлекторный ответ на афферентную информацию, индуцируя сокращение гладких мышц в таких органах, как глаза, легкие, пищеварительный тракт и желчный пузырь, и влияя на работу сердца и желез. Примеры действия соматической нервной системы - афферентные стимулы от кожи и органов чувств (например, реакция на световые стимулы) и эфферентные импульсы к скелетной мускулатуре (например, кашель и рвота).
Простые рефлексы могут быть внутри органа (например, в желудке), но сложные (комплексные) рефлексы идут через контролирующие вегетативные центры в ЦНС, в основном в спинном мозге (А). Эти центры контролируются гипоталамусом, активность которого связана с вегетативной нервной системой. Кора головного мозга -самый высокоорганизованный нервный центр, связывающий вегетативную нервную систему с другими системами.
Периферическая ВНС состоит из симпатического и парасимпатического отделов (А), которые в большинстве случаев действуют независимо. Вегетативные центры симпатического отдела расположены в грудном и поясничном отделах спинного мозга, а центры парасимпатического отдела - в стволовом отделе головного мозга (глаза, железы и органы, иннервируемые блуждающим нервом) и в крестцовом отделе спинного мозга (мочевой пузырь, нижняя часть толстой кишки и половые органы) (А). Преганглионарные волокна обоих отделов вегетативной нервной системы тянутся от центров к ганглиям, где заканчиваются на постганглионарных нейронах.
Преганглионарные симпатические нейроны, начинающиеся в спинном мозге, заканчиваются или в околопозвоночной ганглионарной цепи, в шейном или брюшном ганглии, или в так называемых терминальных ганглиях. Передача стимула от преганглионарных к постганглионарным нейронам является холинергической, т. е. опосредована высвобождением нейромедиатора ацетилхолина. Стимуляция постганглионарными симпатическими волокнами всех эффекторных органов, кроме потовых желез, является адренергической, т. е. опосредована высвобождением норадреналина.
Парасимпатические ганглии расположены вблизи или внутри эффекторных органов. Синаптическая передача в парасимпатических ганглиях и эффекторных органах является холинергической (А).
Большинство органов иннервируется симпатическими и парасимпатическими нервными волокнами. Однако ответы органов на активность этих двух систем могут быть или антагонистическими (как в сердце), или комплементарными (в половых органах).
Мозговое вещество надпочечников - это и ганглий, и эндокринная железа одновременно. Преганглионарные симпатические волокна в мозговом веществе надпочечников высвобождают ацетил-холин, что ведет к выделению в кровоток адреналина (и некоторого количества норадреналина).
Ацетилхолин (АЛ) служит нейромедиатором не только двигательной концевой пластинки и в центральной нервной системе, но и в вегетативной нервной системе, ВНС , где проявляет активность
- во всех преганглионарных волокнах ВНС
- во всех парасимпатических постганглионарных нервных окончаниях
- в некоторых симпатических постганглионарных нервных окончаниях (потовые железы).
Синтез ацетилхолина. Ацетилхолин (АХ) синтезируется в цитоплазме нервных окончаний, а ацетилкофермент А (ацетил-КоА) - в митохондриях. Реакция ацетил-КоА + холин катализируется холинацетилтрансферазой, которая синтезируется в соме и достигает нервных окончаний при помощи аксоплазматического транспорта. Поскольку холин должен быть удален из внутриклеточной среды при помощи переносчиков, этот этап является лимитирующей стадией в процессе синтеза ацетилхолина.
Высвобождение ацетилхолина. Пузырьки у пресинаптического нервного окончания высвобождают содержимое в синаптическую щель, когда концентрация Са2+ в цитоплазме возрастает в ответ на потенциал действия (ПД) (А). Адреналин и норадреналин могут ингибировать высвобождение ацетилхолина путем стимуляции пресинаптических a2-адренорецепторов. В постганглионарных парасимпатических волокнах ацетилхолин блокирует свое собственное высвобождение при помощи связывания с пресинаптическими ауторецепторами (М-рецепторами; см. ниже), как показано на рис. Б.
Ацетилхолин связывается с постсинаптическими холинергическими рецепторами, или холинорецепторами, в вегетативных ганглиях и органах, иннервируемых парасимпатическими волокнами, например в сердце, в гладкой мускулатуре (глаз, бронхов, мочеточников, мочевого пузыря, гениталий, кровеносных сосудов, пищевода и желудочно-кишечного тракта), в слюнных и слезных железах, а также в симпатически иннервируемых потовых железах. Холинорецепторы бывают никотинергическими (N) или мускаринергическими (М). N-холинорецепторы (никотинергические) стимулируются алкалоидом никотином, а М-холинорецепторы (мускаринергические) - алкалоидом яда грибов мускарином.
NN-холинорецепторы вегетативных ганглиев, специфичные для нервов (А), отличаются от специфичных для мышц NM-холинорецепторов на двигательной концевой пластинке тем, что образованы разными субъединицами. Они сходны тем, что и те и другие являются ионотропными рецепторами, т. е. они действуют одновременно как холинорецепторы и как катионные каналы. Связывание ацетилхолина ведет к быстрому притоку ионов Na+ и Са2+ и к раннему (быстрому) возбуждающему постсинаптическому потенциалу действия (ВПСП), который запускает постсинаптический потенциал действия (ПД), как только превышается пороговый потенциал (А, левая часть).
М-холинорецепторы (М1-М5) воздействуют на синаптическую передачу не прямо, а посредством G-белков (метаботропные рецепторы).
М—холинорецепторы находятся в основном в вегетативных ганглиях (А), в ЦНС, в клетках желез внешней секреции. Они активируют фосфолипазу Сβ (ФЛ-Сβ) с помощью Gq-белков в постганглионарных нейронах, в результате чего высвобождаются инозитолтрифосфат (ИТФ) и диацилглицерол (ДАГ) (вторичные мессенджеры). Это стимулирует приток Са2+ и поздний возбуждающий постсинаптический потенциал (ВПСП) (А, средняя часть рисунка). Передача синаптического сигнала модулируется поздним ВПСП, а также котрансмиттерными пептидами, запускающими пептидергический ВПСП или ингибирующий постсинаптический потенциал СИПСП1 (А, правая часть).
М2-холинорецепторы расположены в сердце и функционируют в основном опосредованно, через Gj-белки. Gj-белок открывает специфические К+-каналы, локализованные в основном в синусном узле, атриовентрикулярном узле и в клетках предсердия, таким образом оказывая отрицательный хронотропный и дромотропный эффекты на сердце (Б). Gj-белки также ингибируют аденилатциклазу, снижая таким образом приток Са2+ (Б).
М3-холинорецепторы расположены в основном в гладких мышцах. Как и М1-холинорецепторы (А, средняя часть рисунка), М3-холинорецепторы запускают сокращение путем стимуляции входа Са2+ в клетку. Однако они также могут индуцировать релаксацию путем активации Са2+-зависимой NO-син-тазы, например в клетках эндотелия.
Конец действия ацетилхолина достигается путем опосредованного ацетилхолинэстеразой расщепления молекулы ацетилхолина в синаптической щели. Примерно 50% высвобождаемого холина вновь поглощается пресинаптическими нервными окончаниями (Б).
Антагонисты. Атропин блокирует все М-холинорецепторы, тогда как пирензепин, метоктрамин, гекса-гидрозиладифенидол и тропикамид селективно блокируют M1-, М2-, М3- и М4-холинорецепторы соответственно, тубокурарин- NM-холинорецепторы, а триметафан- NM-холинорецепторы.
Некоторые нейроны могут образовывать (при участии ферментов) L-допа или L-дофа (L-дигидрооксифенилаланин) из аминокислоты L-тирозина. L-допа - предшественник дофамина, норадреналина и адреналина - трех природных катехоламинов, которые синтезируются в указанной последовательности. Дофамин является финальным продуктом синтеза в нейронах, содержащих только ферменты, необходимые на первой стадии синтеза (это декарбоксилаза ароматических L-аминокислот). Дофамин используется в качестве медиатора дофаминергическими нейронами в ЦНС и вегетативными нейронами, иннервирующими почки.
Норадреналин (НА) синтезируется в том случае, если присутствует также второй фермент (дофамин-β-карбоксилаза). В большинстве симпатических постганглионарных нервных окончаниях и в норадре-нергических центральных нейронах норадреналин служит нейромедиатором наряду с комедиаторами -аденозинтрифосфатом (АТФ), соматостатином и нейропептидом Y (НП-Y).
В мозговом веществе надпочечников (см. ниже) и в адренергических нейронах продолговатого мозга N-метилтрансфераза фенилэтаноламина превращает норадреналин в адреналин (А).
Окончания немиелинизированных симпатических постганглионарных нейронов являются шишковидными (узловатыми), или варикозными (А). Эти узлы представляют собой синаптические контакты, хотя и не всегда близкие, с эффекторными органами. Они также служат участками синтеза и хранения норадреналина. L-Тирозин (А1) активно поглощается нервными окончаниями и превращается в дофамин. При адренергической стимуляции эта стадия ускоряется зависимым от протеинкиназы А (ПК-A; А2) фосфорилированием соответствующего фермента. Это приводит к увеличению запасов дофамина. Дофамин помещается в хромаффинные везикулы, где превращается в норадреналин (АЗ). Норадреналин, будучи конечным продуктом, ингибирует дальнейший синтез дофамина (отрицательная обратная связь).
Высвобождение норадреналина. Норадреналин высвобождается в синаптическую щель после поступления в нервные окончания потенциала действия и инициации притока Са2+ (А4).
Адренергические рецепторы, или адренорецепторы (В). Различают четыре основных типа адренорецепторов (а1, а2, β1 и β2) в зависимости от их аффинности к адреналину и норадреналину, а также к многочисленным агонистам и антагонистам. Все адренорецепторы реагируют на адреналин; однако норадреналин оказывает слабое действие на β2-адренорецепторы. Изопротеренол (изопреналин) активирует только β-адренорецепторы, а фентоламин блокирует только а-адренорецепторы. Активность всех адренорецепторов опосредована G-белками.
Выделяют различные подтипы (a1A, а1B. а1D) a1-адренорецепторов (Б1). Их расположение и функции; ЦНС (усиление симпатической активности), слюнные железы, печень (усиление гликогенолиза), почки (действие на порог высвобождения ренина) и гладкая мускулатура (сигнал к сокращению артериол, матки, различных протоков, бронхиол, мочевого пузыря, желудочно-кишечных сфинктеров и расширителя зрачка).
Активация а1-адренорецепторов (Б1), опосредованная Gq-белками и фосфолипазой Сβ (ФЛ-Сβ), ведет к формированию вторичных мессенджеров: инозитолтрифосфата (ИТФ), который увеличивает внутриклеточную концентрацию Са2+, и диацилглицерола (ДАГ), который активирует протеинкиназу С (ПК-С). Опосредованная Gj-белками активация a1-адренорецепторов также приводит к активации Са2+-зависимых К+-каналов. Результирующая утечка К+ гиперполяризует и расслабляет мишень - гладкую мышцу (например, в желудочно-кишечном тракте).
Различают три подтипа (a2A, а2B, а2C) a2-адренорецепторов (Б2). Их расположение и функции: ЦНС (снижение симпатической активности; например, использование а2-агониста клонидина для снижения кровяного давления), слюнные железы (уменьшение слюноотделения), островковые клетки поджелудочной железы (снижение секреции инсулина), жировые клетки (снижение липолиза), тромбоциты (увеличение агрегации) и нейроны (пресинаптические ауторецепторы, см. ниже). Активированные а2-адренорецепторы (Б2) связываются с G1 белком и ингибируют (посредством аi-субъединицы Gj-белков) аденилатциклазу (синтез цАМФ) и в то же время при помощи βу-субъединицы Gj-белков увеличивают вероятность открывания зависимых от потенциала К+-каналов (гиперполяризация). Так как аг-адренорецепторы сопряжены с Go-белками, они также ингибируют зависимые от потенциала Са2+-каналы (снижая [Са2+]внутр ).
Все β-адренорецепторы сопряжены с GS-белком и их aS-субъединицы высвобождают цАМФ как вторичный мессенджер. Затем цАМФ активирует протеинкиназу А (ПК-A), которая фосфорилирует различные белки, что зависит от типа клеток-мишеней.
Норадреналин и адреналин действуют через β1-адренорецепторы (БЗ) и открывают посредством цАМФ и протеинкиназы А Са2+-каналы L-типа в мембранах клеток сердца. Это увеличивает [Са2+]внутр. и таким образом производит положительные хронотропные, дромотропные и инотропные эффекты. Активированные Gs-белки могут также напрямую увеличивать вероятность открывания зависимых от потенциала Са2+-каналов в сердце (БЗ). В почках базальная секреция ренина увеличивается посредством β1-адренорецепторов.
Активация β2-адренорецепторов адреналином (БД) увеличивает уровень цАМФ, таким образом снижая [Са2+]внутр. (механизм этого процесса пока не ясен). Это расширяет бронхиолы и кровеносные сосуды в скелетных мышцах и расслабляет мышцы матки, различных протоков и желудочно-кишечного тракта. Дальнейшие эффекты активации β2-адренорецепторов - увеличенная секреция инсулина и гликогенолиз в печени и мышцах, а также пониженная агрегация тромбоцитов.
Адреналин также увеличивает высвобождение норадреналина в норадренергических волокнах посредством активации пресинаптических β2-адренорецепторов (Д2, А5).
Продукция тепла увеличивается при помощи β3-адренорецепторов клеток бурого жира.
Норадреналин в синаптической щели инактивируется при помощи (А6а-г):
- диффузии норадреналина из синаптической щели в кровь;
- вненейронного поглощения адреналина (в сердце, железах, гладких мышцах, нейроглии и печени) и последующей внутриклеточной деградации норадреналина посредством катехоламин-O-метилтрансферазы (КОМТ) и моноаминоксидазы (МАО);
- активного вторичного поглощения норадреналина (70%) пресинаптическим нервным окончанием. Некоторое количество поглощенного норадреналина входит во внутриклеточные везикулы (АЗ) и используется повторно, а остальная часть инактивируется при помощи МАО;
- стимуляции пресинаптических а2-адренорецепторов (ауторецепторов А6г, 7) норадреналином синаптической щели, который ингибирует дальнейшее высвобождение норадреналина.
Пресинаптические а2-адренорецепторы также можно обнаружить в холинергических нервных окончаниях, например в желудочно-кишечном тракте (снижение подвижности) и предсердии (отрицательный дромотропный эффект), а пресинаптические М-холинорецепторы присутствуют в норадренергических нервных окончаниях. Их взаимодействие позволяет в некоторой степени регулировать периферическую нервную систему. Мозговое вещество надпочечников
После стимуляции преганглионарных симпатических волокон (холинергическая передача) 95% всех клеток в мозговом веществе секретирует в кровь при помощи экзоцитоза гормон адреналин (А), а другие 5% - высвобождают норадреналин (НА). Синтез норадреналина в мозговом веществе аналогичен таковому в норадренергических нейронах (см. выше), но большая часть норадреналина покидает везикулы и превращается в цитоплазме в адреналин. Специальные везикулы, называемые хромаффинными тельцами, после этого активно запасают адреналин и готовятся к высвобождению его и комедиаторов: знкефалина, нейропептида Y путем экзоцитоза.
При реакциях организма на стресс секреция адреналина (и некоторого количества норадреналина) мозговым веществом существенно увеличивается - ответ на физический, ментальный или эмоциональный стресс. Таким образом, клетки, не иннервированные симпатически, активируются и при стрессовой реакции. Адреналин увеличивает высвобождение норадреналина из нейронов посредством пресинаптических β2-адренорецепторов (А2). Секреция адреналина мозговым веществом (опосредованная увеличением симпатической активности) стимулируется некоторыми триггерами, например физической работой, холодом, теплом, тревогой, гневом (стрессом), болью, дефицитом кислорода или снижением кровяного давления. Например, при тяжелой гипогликемии ( Нехолинергические и неадренергические нейромедиаторы Править
У человека гастринвысвобождающий пептид (ГВП) и вазоактивный интестинальный (кишечный) пептид (ВИП) служат комедиаторами в преганглионарных симпатических волокнах; а нейропептид Y (НП-Y) и соматостатин - в постганглионарных волокнах. Постганглионарные парасимпатические волокна используют в качестве комедиаторов нейропептиды энкефалин, вещество Р и/или НП-Y.
Первичной целью преганглионарной секреции пептидов, по-видимому, является регуляция постсинаптических нейронов. Существует весомое доказательство того, что АТФ (аденозинтрифосфат), НП-Y и ВИП также функционируют как независимые нейромедиаторы в вегетативной нервной системе. ВИП и ацетилхолин часто присутствуют одновременно (но в нескольких разных везикулах) в парасимпатических волокнах кровеносных сосудов, экзокринных железах и потовых железах. В желудочно-кишечном тракте ВИП (вместе с оксидом азота) индуцирует истощение слоя круговых мышц и мышц сфинктера, а также (вместе с комедиаторами динорфином и галанином) увеличивает желудочную секрецию. Oксид азота (N0) высвобождается из нитроергических нейронов.
Читайте также:
