
Окклюзия в цнс это
Благодаря конвергенции координация рефлекторной деятельности осуществляется с реализацией таких принципов, как облегчение и окклюзии.
Облегчение - превышение эффекта одновременного действия двух относительно слабых афферентных возбуждающих входов в ЦНС над суммой их раздельных эффектов. Объясняется это суммацией ВПСП с увеличением амплитуды в КРД в группе мотонейронов, в которых ВПСП достигают только значений подпороговых при разрешении активации входов.
Окклюзия - уменьшение эффекта одновременного возбуждения двух сильных афферентных входов, по сравнению с суммой эффектов при раздельном их раздражении.
Причина окклюзии заключается в том, что эти афферентные входы в результате конвергенции адресуются к общим "пороговых" мотонейронов и каждый может создать в них такое же надпороговая возбуждения, как и оба входа вместе.
Доминанта, по А. А. Ухтомским, - господствующий очаг возбуждения, который усиливается импульсами, поступающими из других центров, тормозя при этом остальные.
Доминанта формируется под влиянием адекватных стимулов извне или из внутренней среды, а также под влиянием гуморальных агентов, например, гормонов. Доминантными становятся те центры, которые обеспечивают удовлетворение жизненно необходимых потребностей. Доминантой может быть пищевая, половая, защитная потребность, определяет поведение в определенный момент. Для доминантного центра характерны следующие особенности: повышенная возбудимость, инертность возбуждения, пространственная суммация, устойчивость и продолжительность возбуждения, затормаживание других центров.
Доминанта в своем формировании проходит несколько стадий:
■ начальная - иррадиация (генерализация) возбуждения, которая характеризуется распространением возбуждения на многие другие центров;
■ концентрация возбуждения - происходит в центре биологически значимой в настоящее время реакции.
Отрицательная сторона доминанты - ее консервативность. Разрушить доминанту трудно: для этого необходимо создать новую, еще более сильную доминанту.
Пластичность - способность изменять реактивность под влиянием последовательных раздражений. Это свойство является необходимым условием для обеспечения нервных механизмов обучения.
Процессы обучения сопровождаются микроструктурными изменениями в нервных синапсах, особенно в области аксонного бугорка и на дендритах. Происходят и медиаторные сдвиги. Проявлением пластичности также способность нервных центров принимать на себя функцию утраченного или поврежденного центра. Именно благодаря этому свойству при повреждении или исключении частей мозга выполнения функций утраченных центров обеспечивают другие центры.
Структурной основой пластичности после исключения центра, во-первых, может быть восстановление нервных связей (на замену утраченным) благодаря формированию спраутинг - разветвления аксонных коллатералей, отходящие от противоположной стороны (от другой, неповрежденной полушария). Во-вторых, определенное значение имеет переобучение нейронов, в частности синапсов с явлениями "проторения" нервного пути по типу посттетанического потенциации путем изменений в пресинаптических мембранах и мобилизации большего количества медиатора, а также изменений аксотранспорту, следовательно и трофических процессов.
Кора полушарий - самый молодой и самый отдел ЦНС - обладает максимальной пластичностью и обеспечивает пластичность других мозговых центров. Нейрофизиологические эксперименты вооружают современную восстановительную неврологию и нейрохирургию новыми подходами к реабилитации (обучение языку после поражения моторного центра Брока, переобучение после протезирования).
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ ОБ ИНТЕГРАТИВНОЙ ДЕЯТЕЛЬНОСТИ МОЗГА
Интегративная деятельность ЦНС сводится к подчинению и объединения всех функциональных элементов организма в целостную систему, которая обладает определенной направленностью деятельности.
В осуществлении интегративной функции участвуют разные уровни ЦНС. Первый уровень интеграции осуществляется в результате взаимодействия ВПСП и ТПСП, генерируемые при активации синаптических входов нейрона. В том случае, когда возбуждающие и тормозные входы активируются одновременно, возникает суммирование синаптических потенциалов противоположной полярности и мембранный потенциал может отдаляться от того КРД, при котором в клетках низкопороговых зоны возникает ПД. Конвергенция возбуждающих и тормозных входов на мембране нейрона определяет частоту импульсов, генерируемых и выступает в качестве универсального фактора интегративной деятельности нервной клетки.
Второй уровень интеграции заключается в координационном деятельности нейронов элементарных нервных сетей и зависит от специфики морфологических отношений в ЦНС. Имеются в виду рассмотрены дивергенция, конвергенция, принцип общего конечного пути и принцип обратной связи.
Сложность задач, выполняемых нервной системой, приводит к высокой специализации нейронных объединений - нервных центров (третий уровень), деятельность которых связана с осуществлением различных функций. Функционирование ЦНС в целом осуществляется благодаря деятельности значительного количества таких центров, объединенных с помощью синаптических контактов и отличаются сложностью внутренних и внешних связей.
Нейрогуморальная регуляция функций обеспечивает адаптационные реакции от субклеточных до поведенческих. Можно выделить два вида механизмов, которые заложены в ее основе: жесткие (фиксированные) и гибкие (нефиксированные) (Бехтерева Η. П.).
В основе работы жестких механизмов регуляции лежат генетические программы, которые зафиксированы в процессе эволюции и осуществляют регуляцию достижения постоянно присутствующей цели. Например, регуляция функционирования дыхательного центра; чередование вдоха и выдоха обусловливается генотипическими механизмами; фенотипически же могут меняться продолжительность каждой фазы и амплитуда этих процессов в зависимости от потребности организма.
Гибкие механизмы обеспечивают выполнение организмом сегодняшних задач благодаря организации определенной функциональной системы и тут же прекращают функционирование. Осуществляются они срочно создаваемыми нейронными ансамблями. При этом главным принципом объединения нейронов является доминанта, которая обеспечивает синхронизацию работы нервных образований, входящих в ансамблей.
Программа нейрогуморальной регуляции функций реализуется осуществлением эфферентных влияний на исполнительные органы, работа которых обеспечивает адекватные изменения параметров, которые регулируются. Выделяют три вида таких воздействий: пусковые - вызывают или причиняют деятельность структуры, которую регулируют; адаптационные - влияют на величину реакции и соотношения отдельных ее компонентов; влияние готовности - формирует состояние готовности к ответам на пусковые и адаптационные воздействия.
Пространственная суммация. Это явление, в результате которого происходит увеличение силы сигнала, передается путем вовлечения в его передачу все большего числа сенсорных волокон. Совокупность таких рецепторов, сформированная одним болевым волокном, часто охватывает область кожи размером до 5 см в диаметре. Эту область называют рецепторным полем данного волокна. Количество окончаний в центре поля велико, но уменьшается к периферии.Разветвления разных болевых волокон частично перекрываются в пространстве, поэтому булавочный укол кожи обычно одновременно стимулирует окончания многих волокон. Поскольку число свободных окончаний отдельного болевого волокна в середине его рецепторного поля намного больше, чем на периферии, булавочный укол в центре поля стимулирует это волокно в гораздо большей степени, чем такой же укол на периферии.
Временная суммация. Вторым способом передачи сигналов возрастающей силы является увеличение частоты нервных импульсов в каждом волокне, которое называют временной суммацией.
Большое значение в координации процессов в ЦНС имеет временное и пространственное облегчение (или суммация). Временное облегчение проявляется в повышении возбудимости нейронов в ходе последовательных ВПСП в результате ритмических стимуляций нейрона. Это вызвано тем, что длительности ВПСП продолжаются дольше, чем рефрактерный период аксона. Пространственное облегчение в нервном центре наблюдается при одновременной стимуляции, например, двух аксонов. При раздельной стимуляции каждого из аксонов возникают подпороговые ВПСП в определенной группе нейронов в нервном центре. Совместное раздражение этих аксонов дает большее количество нейронов с надпороговым возбуждением (возникает ПД).
Кратко:Временное и пространственное облегчение – это усиление рефлекторной реакции при действии ряда последовательных раздражителей или одновременном их воздействии на несколько рецептивных полей. Объясняется явлением суммации в нервных центрах.
Противоположное облегчению в ЦНС существует явление окклюзии. Окклюзия – это взаимодействие двух потоков возбуждения между собой.Впервые окклюзия была описана Ч. Шеррингтоном. Сущность окклюзии заключается во взаимном угнетении рефлекторных реакций, при котором суммарный эффект оказывается значительно меньше, чем сумма взаимодействующих реакций. Согласно Ч. Шеррингтону, явление окклюзии объясняется перекрытием синаптических полей, образуемых афферентными звеньями взаимодействующих реакций. В связи с этим при поступлении двух афферентных посылок ВПСП вызывается каждым из них отчасти в одних и тех же мотонейронах спинного мозга. Окклюзия используется в электрофизиологических экспериментах для определения общего звена для двух путей распространения импульсов.
Кратко:Окклюзия явление противоположное облегчению. Когда рефлекторная реакция на два или более сверхпороговых раздражителя меньше, чем ответы на их раздельное воздействие. Оно связано с конвергенцией нескольких возбуждающих импульсов на одном нейроне.
Еще раз:Центральное облегчение - объясняется особенностями строения нервного центра. Каждое афферентное волокно, входя в нервный центр, иннервирует определенное количество нервных клеток. Эти нейроны -нейронный пул. В каждом нервном центре много пулов. В каждом нейронном пуле - 2 зоны: центральная (здесь афферентное волокно над каждым нейроном образует достаточное для возбуждения количество синапсов), периферическая или краевая кайма (здесь количество синапсов недостаточно для возбуждения). При раздражении возбуждаются нейроны центральной зоны. Центральное облегчение: при одновременном раздражении 2-х афферентных нейронов ответная реакция может быть больше арифметической суммы раздражения каждого из них, т. к. импульсы от них отходят к одним и тем же нейронам периферической зоны.
Окклюзия - при одновременном раздражении 2-х афферентных нейронов ответная реакция может быть меньше арифметической суммы раздражения каждого из них. Механизм: импульсы сходятся к одним и тем же нейронам центральной зоны. Возникновение окклюзии или центрального облегчения зависит от силы и частоты раздражения. При действии оптимального раздражителя, (максимального раздражителя (по силе и частоте) вызывающего максимальную ответную реакцию) - появляется центральное облегчение. При действии пессимального раздражителя (с силой и частотой вызывающих снижение ответной реакции) - возникает явление окклюзии.
Свойство облегчения проведения и окклюзии нервного импульса — результат конвергенции (схождения) нервных импульсов от разных аксонов к одной нервной клетке.Для генерации нервного импульса может быть недостаточно возбуждения, поступающего к нервной клетке по отростку одного аксона. В этом случае возбуждение от другого аксона, поступающее к той же нервной клетке, облегчает генерацию нервного импульса.
К нервной клетке могут подходить несколько аксонов и каждый из них в отдельности может вызвать возбуждение. При одновременном их возбуждении некоторые сигналы, направляющиеся к нервной клетке, оказываются закупоренными, не находящими выхода на эффекторный аппарат. Разумеется, это лишь общая схема возможного взаимодействия возбуждении, поступающих от разных нервных проводников к одной нервной клетке. В действительности же, когда к одной клетке сходятся возбуждающие или тормозные влияния сотен и тысяч нервных отростков, суммация и окклюзия становятся чисто статистическими явлениями.
Лекция 2. Физиология центральной нервной системы (ЦНС).
ЦНС – система, осуществляющая регуляцию практически всех функций в организме. ЦНС осуществляет связь в единое целое всех клеток и органов нашего организма. С ее помощью происходят наиболее адекватные изменения работы различных органов, направленные на обеспечение той или иной его деятельности. Кроме того, ЦНС осуществляет связь организма с внешней средой, путем анализа и синтеза поступающей к ней информации от рецепторов и формирует ответную реакцию, направленную на поддержание гомеостаза.
Строение ЦНС.
Структурной и функциональной единицей нервной системы является нервная клетка (нейрон). Нейрон -специализированная клетка, способная принимать, кодировать, передавать и хранить информацию, организовывать ответные реакции организма на раздражения, устанавливать контакты с другими нейронами.
Нейрон состоит из тела (сомы) и отростков - многочисленных дендритов и одного аксона (рис1).

Рис.1. Строение нейрона.
Дендриты обычно сильно ветвятся и образуют множество синапсов с другими нервными клетками, что определяет их ведущую роль в восприятии нейроном информации. Аксон начинается от тела клетки аксонным холмиком, функцией которого является генерация нервного импульса, который по аксону проводится к другим клеткам. Аксон сильно ветвится, образуя множество коллатералей (паралелльных путей) и терминалей. Терминаль – окончание аксона, с помощью которого образуется синапс с другой клеткой. В ЦНС терминали формируют нейро-нейрональные синапсы, на периферии (за пределами ЦНС) аксоны образуют либо нейро-мышечные, либо нейросекреторные синапсы. Окончание аксона чаще называют не терминалью, а синаптической бляжкой (или синаптической пуговкой). Синаптическая бляшка – это концевое (терминальное) утолщение аксона, служащее для депонирования медиатора (смотрите лекции по синапсу). Мембрана окончаний содержит большое число потенциалозависимых кальциевых каналов, через которые ионы кальция поступают внутрь окончания при его возбуждении.
В большинстве центральных нейронов (т.е. нейронов ЦНС) ПД первично возникает в области мембраны аксонного холмика, и отсюда возбуждение распространяется по аксону к синаптической бляшке. Таким образом, уникальными особенностями нейрона являются способность генерировать электрические разряды и передавать информацию с помощью специализированных окончаний – синапсов.
Обработка импульсации (обработка информации, трансформация импульсации) - это наиболее значимая функция нейрона, которая осуществляется на аксонном холмике.
Помимо нейронов в ЦНС имеются глиальные клетки, занимающие половину объема мозга. Периферические аксоны (периферические – значит находящиеся за пределами ЦНС) также окружены оболочкой из глиальных клеток. Функции клеток глии многообразны:
1)они являются для нейронов опорным, защитным и трофическим аппаратом;
2)поддерживают определенную концентрацию ионов кальция и калия в межклеточном пространстве;
3)активно поглощают нейромедиаторы, ограничивая, таким образом, время их действия.
Рефлекторный принцип деятельности НС.
Основным механизмом деятельности ЦНС является рефлекс. Рефлекс- это ответная реакция организма на действия раздражителя, осуществляемая с участием ЦНС. Например, отдергивание руки при уколе, смыкание век при раздражении роговицы – это тоже рефлекс. Отделение желудочного сока при попадании пищи в желудок, дефекация при наполнении прямой кишки, покраснения кожи при тепловом воздействии, коленный, локтевой, Бабинского, Розенталя – это все примеры рефлексов Количество рефлексов безгранично. Общим для них всех является обязательное участие в их реализации ЦНС.
Другим определением рефлекса, также подчеркивающим роль ЦНС, является следующее: рефлекс- это центробежный ответ на центростремительное раздражение. (В приведенных примерах самостоятельно определите, что является центробежным ответом, а что раздражением. Раздражение всегда центростремительное, т.е. действующий на рецепторы раздражитель вызывает импульсацию, которая поступает в ЦНС).
Структурной основой рефлекса, его материальным субстратом является рефлекторная дуга (рис.2)

Рис. 2.Рефлекторная дуга
Рефлекторная дуга состоит из 5 звеньев:
2)афферентного (чувствительного, центростремительного) звена;
3)вставочного звена (центрального);
4)эфферентного (двигательного, центробежного) звена;
5)эффектора (рабочего органа);
Участок тела, содержащий рецепторы, при раздражении которых возникает определенный рефлекс, называется рецептивным полем рефлекса.
Рефлекс может осуществляться только тогда, когда сохранена целостность всех звеньев рефлекторной дуги.
Нервный центр.
Нервный центр (центр ЦНС или ядро) – это совокупность нейронов, принимающих участие в осуществлении конкретного рефлекса. Т.е. каждый рефлекс имеет свой центр: существует центр коленного рефлекса, свой центр у локтевого рефлекса, свой - у мигательного, есть сердечно-сосудистый, дыхательный, пищевой центры, центры сна и бодрствования, голода и жажды и т.д. В целом организме при формировании сложных адаптивных процессов происходит функциональное объединение нейронов, расположенных на различных уровнях ЦНС, т.е. сложное объединение большого количества центров.
Объединение нервных центров (ядер) между собой осуществляется проводящими путями ЦНС с помощью нейро-нейрональных (межнейронных) синапсов. Существует 3 типа соединения нейронов: последовательное, дивергентное и конвергентное.
Нервные центры обладают рядом характерных функциональных свойств, которые во многом обусловлены этими тремя типами нейронных сетей, а также свойствами межнейронных синапсов.
Основные свойства нервных центров:
1. Конвергенция (схождение) (рис.3). В ЦНС к одному нейрону могут сходиться возбуждения от различных источников. Эта способность возбуждений сходиться к одним и тем же промежуточным и конечным нейронам получила название конвергенции возбуждений

Рис.3. Конвергенция возбуждения.
2. Дивергенция (расхождение) - расхождение импульсаций от одного нейрона сразу на многие нейроны. На основе дивергенции происходит иррадиация возбуждения и становится возможным быстрое вовлечение в ответную реакцию многих центров, расположенных на разных уровнях ЦНС.

Рис.4. Дивергенция возбуждения.
3 Возбуждение в нервных центрах распространяется односторонне - от рецептора к эффектору, что обусловливается свойством химических синапсов односторонне проводить возбуждение от пресинаптической мембраны к постсинаптической.
4. Возбуждение в нервных центрах проводится медленнее, чем по нервному волокну. Это обусловлено замедленным проведением возбуждения через синапсы (синаптическая задержка), которых в ядре много.
5. В нервных центрах осуществляется суммация возбуждений. Суммация – сложение допороговых импульсов. Различают два вида суммации.
Временная или последовательная, если импульсы возбуждения приходят к нейрону по одному и тему же пути через один синапс с интервалом меньше, чем время полной реполяризации постсинаптической мембраны. В этих условиях локальные токи на постсинаптической мембране воспринимающего нейрона суммируются и доводят ее деполяризацию до уровня Ек, достаточного для генерации нейроном потенциала действия. Временной данная суммация называется, потому что на нейрон в течение некоторого промежутка времени приходит серия импульсов (раздражений). Последовательной она называется, потому что реализуется в последовательном соединении нейронов.
Пространственная или одновременная - наблюдается в том случае, когда импульсы возбуждения поступают к нейрону одновременно через разные синапсы. Пространственной данная суммация называется, потому что раздражитель действует на некоторое пространство рецептивного поля, т.е. несколько (минимум 2) рецепторов разных участков рецептивного поля. (Тогда как временная суммация может реализоваться при действии серии раздражителей на один и тот же рецептор). Одновременной она называется, потому что информация к нейрону приходят одновременно по нескольким (минимум 2) каналам связи, т.е. одновременная суммация, реализуется конвергентным соединением нейронов.
6.Трансформация ритма возбуждения - изменение количества импульсов возбуждения, выходящих из нервного центра, по сравнению с числом импульсов, приходящих к нему. Различают два вида трансформации:
1) понижающая трансформация, в основе которой, лежит явление суммации возбуждений, когда в ответ на несколько пришедших допороговых возбуждений к нервной клетке, в нейроне возникает только одно пороговое возбуждение;
2) повышающая трансформация, в ее основе лежат механизмы умножения (мультипликации), способные резко увеличить количество импульсов возбуждения на выходе.
7. Рефлекторное последействие - заключается в том, что рефлекторная реакция заканчивается позже прекращения действия раздражителя. Это явление обусловлено двумя причинами:
1) длительной следовой деполяризацией мембраны нейрона, на фоне прихода мощной афферентации (сильной чувствительной импульсации), вызывающей выделение большого количества (квантов) медиатора, что обеспечивает возникновение нескольких потенциалов действия на постсинаптической мембране и, соответственно, кратковременное рефлекторное последействие;
2) пролонгированием выхода возбуждения к эффектору в результате циркуляции (реверберации) возбуждения в нейронной сети типа "нейронной ловушки" ( рис. ) Возбуждение, попадая в такую сеть, может длительное время циркулировать в ней, обеспечивая длительное рефлекторное последействие. Возбуждение в такой цепочке может циркулировать до тех пор, пока какое-либо внешнее воздействие затормозит этот процесс или в ней наступит утомление. Примером последействия может служить хорошо всем известная жизненная ситуация, когда даже после прекращения действия сильного эмоционального раздражителя (после прекращения ссоры) еще какое-то более или менее продолжительное время продолжается общее возбуждение, артериальное давление остается повышенным, сохраняется гиперемия лица, тремор кистей.
8. Нервные центры обладают высокой чувствительностью к недостатку кислорода.Нервные клетки отличаются интенсивным потреблением О2. Мозг человека поглощает около 40-70 мл О2 в минуту, что составляет 1/4-1/8 часть всего количества О2, потребляемого организмом. Потребляя большое количество О2, нервные клетки высокочувствительны к его недостатку. Частичное прекращение кровообращения центра ведет к тяжелым расстройствам деятельности его нейронов, а полное прекращение - к гибели в течение 5-20мин.
9. Нервные центры, как и синапсы, обладают высокой чувствительностью к действию различных химических веществ, особенно ядов. На одном нейроне могут располагаться синапсы, обладающие различной чувствительностью к различным химическим веществам. Поэтому можно подобрать такие химические вещества, которые избирательно будут блокировать одни синапсы, оставляя другие в рабочем состоянии. Это делает возможным корректировать состояния и реакции как здорового, так и больного организма.
10. Нервные центры, как и синапсы, обладают быстрой утомляемостью в отличие от нервных волокон, которые считаются практически неутомляемыми. Это обусловлено резким уменьшением запасов медиатора, уменьшением чувствительности к медиатору постсинаптической мембраны, уменьшением ее энергетических запасов, что наблюдается при длительной работе и является основной причиной развития утомления.
11 .Нервные центры, как и синапсы, обладают низкой лабильностью,основной причиной которой является синаптическая задержка. Суммарная синаптическая задержка, наблюдающаяся во всех нейро-нейрональных синапсах при проведении импульсации по ЦНС, или в нервном центре называется центральной задержкой.
12. Нервные центры обладают тонусом, который выражается в том, что даже при отсутствии специальных раздражений, они постоянно посылают импульсы к рабочим органам.
13. Нервные центры обладают пластичностью- способностью изменять собственное функциональное назначение и расширять свои функциональные возможности. Так же пластичность можно определить, как способность одних нейронов брать на себя функцию пораженных нейронов того же центра. Именно, с явлением пластичности связана способность восстанавливать двигательную активность конечностей, например, ног, утраченную в результате травм спинного мозга. Однако это возможно только при поражении части нейронов данного центра или при сохранении целостными части проводящих путей ЦНС. При полном разрыве спинного мозга восстановление двигательной активности оказывается невозможным. Кроме того, нейроны одного центра, например, сгибателей не могут брать на себя функцию нейронов другого центра - разгибателей. Т.е. явление пластичности центров ЦНС ограничено.
14. Окклюзия (запирание) (рис.5) - это сложение пороговой импульсации. Окклюзия осуществляется (так же как и пространственная суммация) в конвергирующей системе соединения нейронов. Одновременной активации нескольких (минимум двух) рецепторов сильным или сверхсильным раздражителями к одному нейрону будут конвергировать несколько пороговых или сверхпороговых импульса. На этом нейроне будет происходить окклюзия, т. е эти два раздражителя он ответит с той же максимальной силой, что и на каждый из них отдельности. Феномен окклюзии состоит в том, что количество возбужденных нейронов при одновременном раздражении афферентных входов обоих нервных центров оказывается меньше, чем арифметическая сумма возбужденных нейронов при отдельном раздражении каждого афферентного входа в отдельности.

Рис.6. Явление окклюзии в ЦНС.
Явление окклюзии приводит к снижению силы ответной реакции. Окклюзия имеет охранительное значение, предотвращая перенапряжение нейронов при действии сверхсильных раздражителей.
Конвергенция и окклюзия
Импульсы, приходящие в центральную нервную систему по различным афферентным волокнам, могут сходиться (конвергировать) к одним и тем же промежуточным и эффекторным нейронам. Этот факт лег в основу принципа конвергенции установленного Шеррингтоном.
Конвергенция нервных импульсов объясняется тем, что на теле и дендритах, каждого нейрона центральной нервной системе оканчиваются аксоны множества других нервных клеток. В спинном и продолговатом мозгу конвергенция имеет сравнительно ограниченный характер: на вставочных и моторных нейронах конвергируют афферентные импульсы, возникающие в различных участках рецептивного поля только одного и того же рефлекса. В отличие от этого в высших оттделах центральной нервной системы - подкорковых ядрах и в коре больших полушарий - наблюдается конверсия импульсов, исходящих из разных рецептивных зон. Поэтому один и тот же нейрон может возбуждаться импульсами, возникающими при раздражении и слуховых, и зрительных, и кожных рецепторов.
Конвергенция объясняет пространственную суммацию возбуждений и явлении окклюзии. Сущность окклюзии (закупорки) состоит в том, что при одновременном раздражении двух групп афферентных волокон, каждая из которых вызывает сильное рефлекторное сокращение мышцы возникает эффект, который по своей величине оказывается меньше арифметической суммы величин этих рефлексов, взятых порознь.
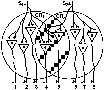
Так, максимальное раздражение у собаки одного из nn. plantares, содержащего афферентные волокна, сопровождается рефлекторным сокращением m. tibialis anticus, напряжение которого равно 1,57 кг, а раздражение второго n. Plantaris этой же конечности вызывает напряжение той же мышцы, равное 1.58 кг. При одновременном же раздражении нервов величина напряжения мышцы составляет всего лишь 1.81 кг, что намного меньше арифметической суммы напряжений при этих рефлексах. Зависимость такого эфффекта от конвергенции становится понятной при рассмотрении рис. 182 .
На этом рисунке представлены моторные нейроны, получающие импульсы от двух групп афферентных волокон А и В. При максимальном раздражении каждой из этих групп и в отдельности возбуждение охватывает по 4 нейрона. При совместном же раздражении А и В возбуждение возникает не в 8, а только в 6 нейронах, так как 2 нервные клетки вследствие конвергенции иннервируются обоими группами нервных кон. Поэтому рефлекторный ответ совместного раздражения волокон А и В меньше арифметической суммы этих рефлексов, вызнанных порознь.
В нормальной деятельности нервных центров пространственная суммация и окклюзия постоянно перемежаются друг с другом, становясь то более, то менее выраженными, в зависимости от силы взаимодействующих афферентных раздражении: при слабой их силе на первый план выступает суммация, а при сильных раздражениях — окклюзия .
Рис. 182. Схема, иллюстрирующая явлении окклюзии (по Ч. Шеррингтону). Внутри круга изображено большое число моторных клеток. Шесть клеток показаны с отходящими от них аксонами. А и В — нервные полокна, приносящие возбуждения с периферии к моторным нейронам. При максимальном раздражении волокна А или волокна В разряды импульсов возникают в 4 нейронах: два из них возбуждаются обоими волокнами.
Окклюзия — взаимодействие двух импульсных потоков между собой. Впервые явление окклюзии было описано Ч.Шеррингтоном. Сущность его заключается во взаимном угнетении рефлекторных реакций, при котором суммарный результат оказывается значительно меньше, чем сумма взаимодействующих реакций
Окклюзия (occlusio; лат. occludo, occlusum закрывать, замыкать)
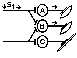
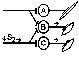
При последовательном раздражении
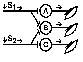
При одновременном раздражении
S1 + S2 > S’1 + S’2
Согласно Ч.Шеррингтону, явление окклюзии объясняется перекрытием синаптических полей, образуемых афферентными звеньями взаимодействующих рефлексов. В связи с этим при одновременном поступлении двух афферентных посылок возбуждающий постсинаптический потенциал вызывается каждым из них отчасти в одних и тех же мотонейронах спинного мозга. Окклюзию используют в электрофизиологических экспериментах для определения общего звена для двух путей распространения импульсов. Если имеется общее звено, то одновременное раздражение двух путей с максимальной интенсивностью вызывает ответ меньшей величины, чем сумма ответов, получаемых при раздельном раздражении этих путей стимулами той же интенсивности.
Приведем два неточных объяснения явления окклюзии.

При объяснении явления окклюзии нужно говорить о перекрытии синаптическихполей нервных центров, а нерецептивныхполей.


Почему это трудно признать правильным объяснением? Нейроны не конвергируют на мышце.
В основе окклюзии лежит явление конвергенции.
Облегчение
В ряде случаев вместо такого ослабления реакции при совместном раздражении рецептивных полей двух рефлексов можно наблюдать феномен облегчения (т.е. суммарная реакция выше суммы реакции при изолированном раздражении этих рецептивных полей). Это результат того, что часть общих для обоих рефлексов нейронов при изолированном раздражении оказывает подпороговый эффект для вызывания рефлекторных реакций. При совместном раздражении они суммируются и достигают пороговой силы, в результате конечная реакция оказывается больше суммы изолированных реакций.
Синоним — суммация. Да да суммация. Какая? Пространственная.
Читайте также:
