
Передача импульса в ганглиях
Ганглии нервной системы — скопление нейронов и глии, находящиеся за пределами головного и спинного мозга.
Подобные образования в центральной нервной системе называют ядрами. Они выступают связующими звеньями структур нервной системы, осуществляют первичную обработку импульсов, отвечают за функции висцеральных органов.
Человеческий организм осуществляет два типа функций – соматические и вегетативные. Соматические подразумевают восприятие внешних раздражителей и реакцию на них при помощи скелетной мускулатуры. Эти реакции могут контролироваться сознанием человека, а за их выполнение отвечает центральная нервная система.
Вегетативные функции – пищеварение, метаболизм, кроветворение, кровообращение, дыхание, потоотделение и другое, контролирует вегетативная система, которая не зависит от человеческого сознания. Кроме регуляции работы висцеральных органов, вегетативная система обеспечивает трофику мускулатуры и ЦНС.
Ганглии, отвечающие за соматические функции, представляют спинномозговые узлы и узлы черепных нервов. Вегетативные, в зависимости от расположения центров в ЦНС, делят на: парасимпатические и симпатические.
Первые находятся в стенках органа, а симпатические дистанционно – в структуре, называемой пограничным стволом.
Строение ганглия
В зависимости от морфологических особенностей, размер ганглиев колеблется от нескольких микрометров до нескольких сантиметров. По сути это скопление нервных и глиальных клеток, покрытое соединительной оболочкой.
Соединительнотканный элемент пронизан лимфатическими и кровеносными сосудами. Каждый нейроцит (или группа нейроцитов) окружен капсульной оболочкой, выстланной изнутри эндотелием, а снаружи волокнами соединительной ткани. Внутри капсулы располагается нервная клетка и глиальные структуры, которые обеспечивают жизнедеятельность нейрона.
От нейрона отходит один аксон, покрытый миелиновой оболочкой, который разветвляется на две части. Одна из них входит в состав периферического нерва и образует рецептор, а вторая направляется в ЦНС.
Вегетативные центры находятся в стволе головного мозга и спинном мозге. Парасимпатические центры локализуются в краниальном и сакральном отделах, а симпатические – в тораколюмбальном.
Ганглии вегетативной нервной системы
Симпатическая система включает два типа узлов: вертебральные и превертебральные.
Вертебральные располагаются с двух сторон от позвоночного столба, образуя пограничные стволы. Они связаны со спинным мозгом с помощью нервных волокон, которые дают начало белым и серым соединительным веткам. Выходящие из узла нервные волокна направляются к висцеральным органам.
Превертебральные расположены на большем расстоянии от позвоночника, при этом также дистанционно они находятся от органов, за которые они ответственны. Примером превертебральных узлов служат шейные, брыжеечные скопления нейронов, солнечное сплетение.
Парасимпаческий отдел образован ганглиями, расположенными на органах, или в непосредственной к ним близости.
Внутриорганные нервные сплетения размещены на органе или в его стенке. Крупные внутриорганные сплетения расположены в сердечной мышце, в мышечном слое стенки кишечника, в паренхиме железистых органов.
Ганглии вегетативной и центральной нервной системы имеют свойства:
- проведение сигнала в одну сторону;
- входящие в узел волокна перекрывают зоны влияния друг друга;
- пространственная суммация (сумма импульсов способна породить потенциал в нейроците);
- окклюзия (стимуляция нервов вызывает малый ответ, чем стимуляция каждого в отдельности).
Синоптическая задержка в вегетативных ганглиях больше, чем в аналогичных структурах ЦНС, а постсинаптический потенциал продолжительный. Волну возбуждения в ганглионарных нейроцитах сменяет депрессия. Эти факторы приводят к относительно низкому ритму импульсов, сравнительно с ЦНС.
Какие функции выполняют ганглии
Основным назначением вегетативных узлов является распределение и передача нервных импульсов, а также генерация локальных рефлексов. Каждый ганглий, зависимости от расположения и особенностей трофики, отвечает за функции определенного участка тела.
Ганглиям присуща автономность от ЦНС, что позволяет им регулировать деятельность органов без участия головного и спинного мозга.
В структуре внутристеночных узлов содержатся клетки – водители ритма, способные задавать периодичность сокращений гладкой мускулатуры кишечника.
Особенность связана с прерыванием, направляющимся к внутренним органам, волокон ЦНС на периферических узлах вегетативной системы, где они образуют синапсы. При этом непосредственное влияние на внутренний орган оказывают аксоны, выходящие из ганглия.
Каждое нервное волокно, приходящее в симпатический ганглий, осуществляет иннервацию до тридцати постганглионарных нейроцитов. Это дает возможность умножать сигнал и распространять импульс возбуждения, выходящий из нервного узла.
В парасимпатические узлах одно волокно осуществляет иннервацию не более четырех нейроцитов, а передача импульса осуществляется локально.
Ганглии – рефлекторные центры
Ганглии нервной системы принимают участие в рефлекторной дуге, что позволяет корректировать деятельность органов и тканей без участия мозга. В конце девятнадцатого века русский гистолог Догель в результате опытов по изучению нервных сплетений в желудочно-кишечном тракте, выявил три типа нейронов – моторные, вставочные и рецепторные, а также синапсы между ними.
Наличие рецепторных нервных клеток подтверждает и возможность трансплантации сердечной мышцы от донора реципиенту. Если бы регуляция сердечного ритма осуществлялась через ЦНС, после пересадки сердца нервные клетки подверглись дегенерации. Нейроны и синапсы в пересаженном органе продолжают функционировать, что свидетельствует об их автономности.
В конце двадцатого века экспериментальным путем установлены механизмы периферических рефлексов, которые делают превертебральные и интрамуральные вегетативные узлы. Возможность создавать рефлекторную дугу свойственна некоторым узлам.
Местные рефлексы позволяют разгрузить центральную нервную систему, делают регуляцию важных функций более надежность, способны продолжать автономную работу внутренних органов в случае прерывания связи с ЦНС.
Вегетативные узлы получают и обрабатывают информацию о работе органов, после чего направляют ее в мозг. Это вызывает рефлекторную дугу как в вегетативной, так и в соматической системе, что запускает не только рефлексы, но и сознательные поведенческие реакции.
а) Ганглионарная передача нервных импульсов. Преганглионарные нейроны симпатической и парасимпатической систем — холинергические: при образовании аксодендритических синапсов с ганглионарными клетками из этих нейронов высвобождается ацетилхолин (АХ). Рецепторы на поверхности ганглионарных клеток называют никотиновыми, так как их возбуждение может происходить при местном действии никотина.
б) Передача нервных импульсов в нейроэффекторном синапсе. Постганглионарные нервные волокна симпатической и парасимпатической систем образуют нейроэффекторные соединения (синапсы)с эффекторными тканями (тканями-мишенями). За высвобождение нейромедиаторов отвечают многочисленные пресинаптические утолщения, расположенные по ходу нервных волокон.
Главный нейромедиатор в симпатических нейроэффекторных соединениях — норадреналин (норэпинефрин), который высвобождается из гранулярных везикул. Постганглионарные симпатические волокна — преимущественно адренергические; исключение составляют холинергические волокна, отвечающие за симпатическую иннервацию расположенных по всему телу эккриновых потовых желез. Главный нейромедиатор в парасимпатических нейроэффекторных соединениях — ацетилхолин (АХ). Постганглионарные парасимпатические волокна преимущественно холинергические.
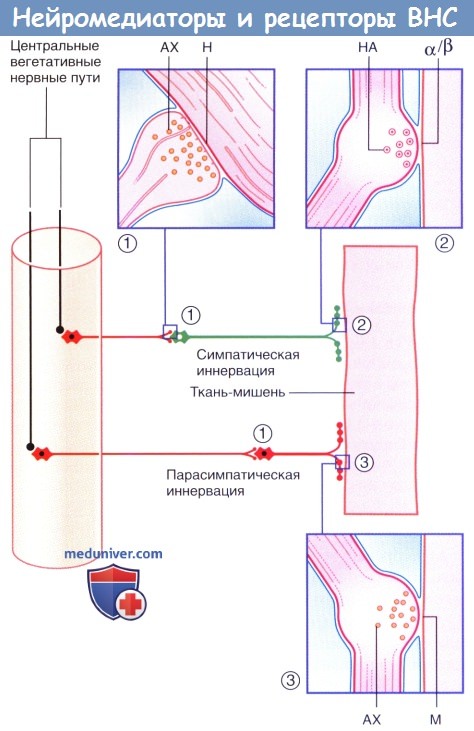
Нейромедиаторы и рецепторы вегетативной нервной системы.
(1) Аксодендритические синапсы с никотиновыми рецепторами.
(2) Нейроэффекторные синапсы с адренергическими рецепторами.
(3) Нейроэффекторные синапсы с мускариновыми рецепторами. Ганглионарные нейроны и постганглионарные волокна выделены красным цветом.
АХ — ацетилхолин; М — мускариновые рецепторы; Н — никотиновые рецепторы; НА — норадреналин.
в) Рецепторы в нейрозффекторном синапсе. Физиологические эффекты вегетативной нервной системы (ВНС) зависят от вида рецепторов на постсинаптической мембране (плазматической мембране эффекторных клеток). На высвобождение нейромедиаторов также влияют рецепторы пресинаптической мембраны (аксолеммы).
- Синаптические рецепторы симпатической системы (адренорецепторы). Для норадреналина существуют два вида а-адренорецепторов и два вида β-адренорецепторов.
1. Постсинаптические α1-адренорецеторы, активация которых вызывает сокращение гладких мышц мелких периферических артерий и крупных артериол, дилататора зрачка, а также мышц семявыносящего протока, сфинктеров ЖКТ и шейки мочевого пузыря.
2. Пресинаптические α2-адренорецеторы локализуются как на симпатических, так и на парасимпатических нервных окончаниях и ингибируют в них высвобождение нейромедиаторов. Пресинаптические α2-адренорецеторы симпатической системы называют ауторецепторами.
3. Постсинаптические адренорецепторы, возбуждение которых приводит к увеличению пейсмекерной активности клеток сердца, а также к повышению силы сердечных (желудочковых) сокращений. При резком падении артериального давления за счет активации симпатических β1-адренорецепторов юкстагломерулярных клеток почек происходит секреция ренина, который, в свою очередь, вызывает секрецию мощного вазоконстриктора ангиотензина II.
4. β2-Адренорецепторы, которые реагируют как на норадреналин, так и на адреналин (эпинефрин).
Активация постсинаптических β2-адренорецепторов вызывает расслабление гладких мышц, наиболее ярко выраженное в трахеобронхиальном дереве и мышцах глаза, участвующих в аккомодации. Часть постсинаптических β2-адренорецепторов располагается на поверхности гепатоцитов. При возрастании энергетических затрат организма их активация вызывает распад гликогена, необходимый для поддержания уровня глюкозы в крови.
Пресинаптические β2-адренорецепторы адренергических нервных окончаний отвечают за высвобождение норадреналина.
В симпатических нервных окончаниях большая часть высвобождаемого норадреналина подвергается обратному захвату моноаминным ферментным насосом. После обратного захвата часть норадреналина разрушается митохондриальным ферментом — моноаминоксидазой (МАО). Влияние лекарственных средств на симпатическую систему подробно рассмотрено в блоке клинической информации ниже.
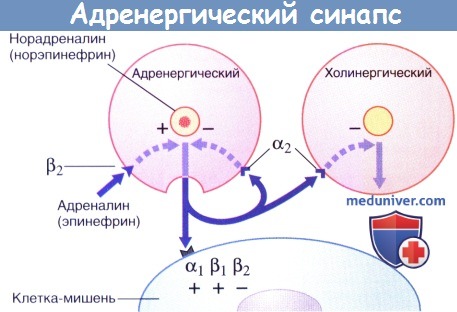
Передача возбуждения в адренергическом нейроэффекторном синапсе.
Секреция норадреналина стимулируется адреналином и ингибируется путем активации пресинаптических α2-рецепторов (данные рецепторы активируются и на пресинаптической мембране близлежащих парасимпатических синапсов, где также ингибируют секрецию).
- Синаптические рецепторы парасимпатической системы. Парасимпатические синаптические рецепторы называют мускариновыми, так как их активацию, помимо АХ, может вызывать другое химическое вещество — мускарин. Стимуляция парасимпатической системы приводит к следующим М-холинергическим эффектам.
• Замедление (в ответ на стимуляцию блуждающего нерва) частоты сердечных сокращений, а также уменьшение силы желудочковых сокращений.
• Сокращение гладких мышц, которое обусловливает такие процессы, как перистальтика кишечника, опустошение мочевого пузыря, а также рефлекс аккомодации при взгляде на близкое расстояние.
• Секреция желез.
Помимо представленных выше эффектов, пресинаптические мускариновые рецепторы симпатических нервных окончаний также отвечают за ингибирование высвобождения норадреналина.
Влияние лекарственных средств на парасимпатическую систему подробно рассмотрено в Блоке клинической информации. Лекарственные средства, обладающие мускариноподобным действием, называют холинергическими (холиномиметическими). Лекарственные средства, блокирующие доступ АХ к постсинаптической мембране, называют антихолинергическими (холинолитическими).
Главный фактор, который необходимо учитывать при назначении препаратов, активирующих или подавляющих деятельность симпатической и парасимпатической систем,— наличие рецепторов к этим лекарственным веществам α-, β- и мускариновых) в ЦНС. Так, в частности, применение данных лекарственных средств в психиатрии обусловлено их действием на центральные, а не на периферические рецепторы.
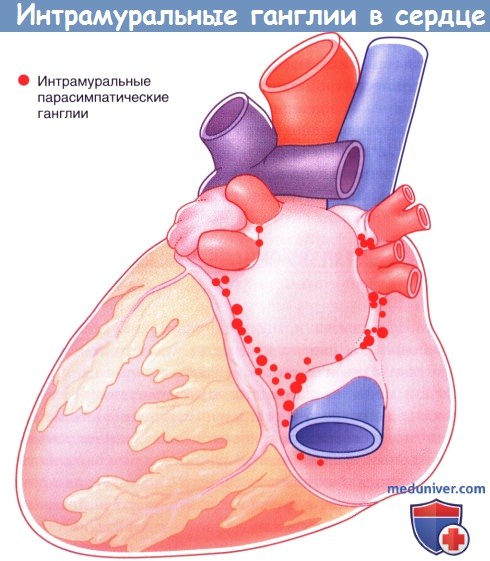
Расположение парасимпатических интрамуральных ганглиев в сердце. 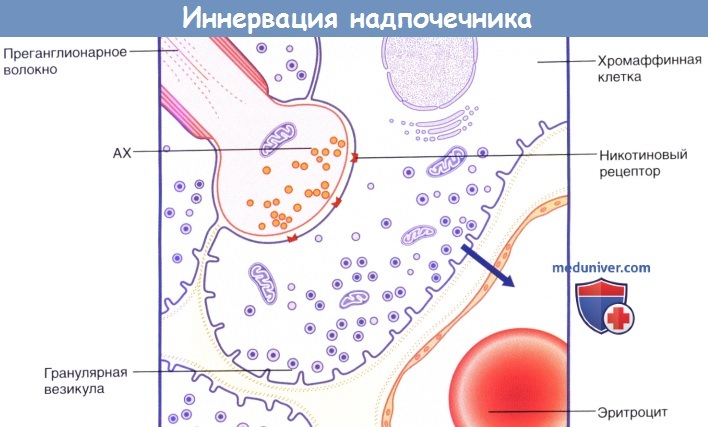
Синаптическое соединение преганглионарного волокна грудного внутренностного нерва и хромаффинной клетки мозгового вещества надпочечников.
Активация никотиновых рецепторов ацетилхолином (АХ).
8% клеток содержат крупные гранулярные везикулы (представлены на рисунке), высвобождающие адреналин; его секрекция в капиллярное русло обозначена стрелкой.
20% клеток содержат маленькие гранулярные везикулы, высвобождающие норадреналин. 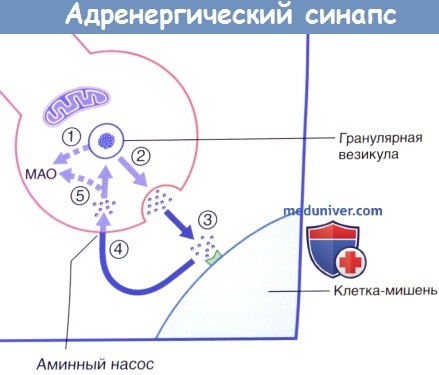
Высвобождение и обратный захват нейромедиатора в адренергическом нервном окончании.
МАО — моноаминоксидаза. 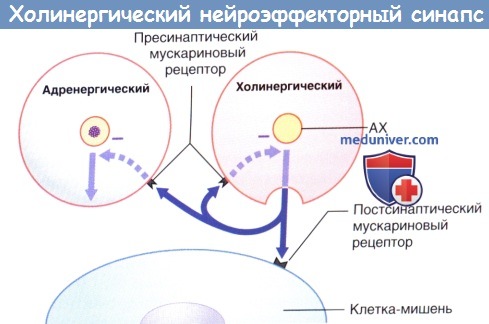
Передача возбуждения в холинергическом нейроэффекторном синапсе.
Избыточная секреция ацетилхолина (АХ) ингибируется путем активации пресинаптических мускариновых рецепторов (данные рецепторы активируются и на пресинаптической мембране близлежащих симпатических синапсов, где также ингибируют секрецию).
г) Другие типы нейронов. И в симпатическом, и в парасимпатическом отделах ВНС часть нейронов представлена неадренергическими, нехолинергическими нейронами (NANC-нейронами). Находящиеся в симпатических ганглиях небольшие вставочные нейроны отвечают за высвобождение дофамина — предшественника норадреналина. Часть дофамина выделяется в капиллярное русло, другая часть связывается с дофаминовыми рецепторами на основных (адренергических) нейронах, оказывая на них слабое тормозное действие.
Самое большое количество NANC-нейронов находится в области ганглионарных клеток стенки ЖКТ и в тазовых ганглиях. Эти нейроны содержат более 50 разнообразных белковых субстанций, находящихся в несвязанном состоянии или в различных комбинациях. Функция большей части этих веществ — регуляторная: действуя на пресинаптическую или постсинаптическую мембрану, они влияют на продолжительность работы основных нейромедиаторов. Другие белковые субстанции (вещества, выделяющиеся совместно с ацетилхолином (АХ)) представляют собой комедиаторы (сопутствующие медиаторы).
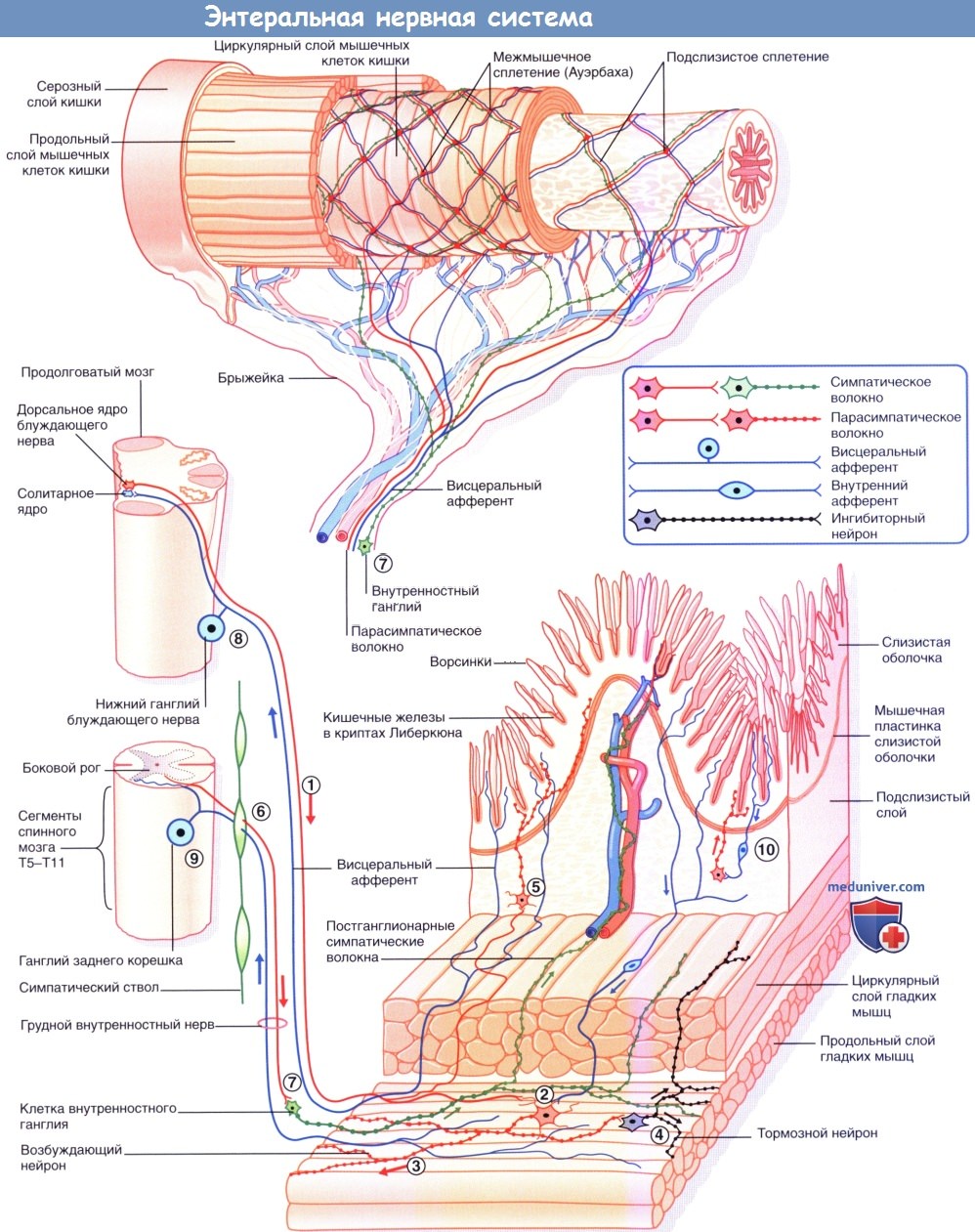
Редактор: Искандер Милевски. Дата публикации: 14.11.2018
Передача возбуждения в вегетативных ганглиях имеет следующие особенности:
— выраженный феномен мультипликации (дивергенции);
— большая длительность синаптической задержки (от 1,5 до 30 мс);
— низкая лабильность ганглионарных нейронов с частотой генерации потенциалов действия в среднем от 2 до 15 Гц.
Передача нервных импульсов с преганглионарных волокон на нейроны всех вегетативных ганглиев осуществляется Н-холинергическими синапсами. Постганглионарные холинергические волокна образуют на клетках исполнительных органов (желез, ГМК органов пищеварения, сосудов и т.д.) М-холинергические синапсы. Их постсинаптическая мембрана содержит мускаринчувствительные рецепторы (блокатор-атропин). И в тех и других синапсах передача возбуждения осуществляется ацетилхолином. М-холинергические синапсы оказывают возбуждающее влияние на гладкие мышцы пищеварительного канала, мочевыводящей системы (кроме сфинктеров), железы ЖКТ. Однако они уменьшают возбудимость, проводимость и сократимость сердечной мышцы и вызывают расслабление некоторых сосудов головы и таза. Постганглионарные симпатические волокна образуют 2 типа адренергических синапсов на эффекторах- α-адренергические и β-адренергические. Постсинаптическая мембрана первых содержит α1-и α2 -адренорецепторы. При воздействии НА на α1-адренорецепторы происходит сужение артерий и артериол внутренних органов и кожи, сокращение мышц матки, сфинктеров ЖКТ, но одновременно расслабление других гладких мышц пищеварительного канала. Постсинаптические β-адренорецепторы также делятся на β1- и b2- типы. b1-адренорецепторы расположены в клетках сердечной мышцы. При действии на них НА повышается возбудимость, проводимость и сократимость кардиомиоцитов. Активация β2-адренорецепторов приводит к расширению сосудов легких, сердца и скелетных мышц, расслаблению гладких мышц бронхов, мочевого пузыря, торможению моторики органов пищеварения.
14. Медиаторы вегетативной нервной системы. Основные виды рецепторов( адренергические, холинергические и др.)
Ацетилхолин-основной медиатор преганглионарных нейронов(симпат и парасимпат). Медиатором постганглионарных парасимпатических нейронов также служит ацетилхолин, симпатических-норадреналин. Т.е. парасимпатические нервы действуют на органы с помощью АХ, а симпатические-норадреналина.
Рецепторы АХ(холинорецепторы) делятся на два основых типа:
-N-холинорецепторы. (чувствительны к никотину)Располагаются на постганглионарных парасимпатических и симпатических нейронах.
-М-холинорецепторы.(чувствительные к яду мухомора мускарину) Располагаются на внутренних органах.
Рецепторы адреналина(адрунорецепторы) делятся на
Ацетилхолин высвобождается в окончаниях холинергических парасимпатических и симпатических волокон. Процесс освобождения медиатора является кальцийзависимым. Инактивация медиатора происходит с помощью фермента ацетилхолинэстеразы.
Ацетилхолин оказывает свое воздействие на органы и ткани посредством специфических холинорецепторов. Действие ацетилхолина на постсинаптическую мембрану постганглионарных нейронов может быть воспроизведено никотином, а действие ацетилхолина на исполнительные органы - мускарином (токсин гриба мухомора). На этом основании холинорецепторы разделили на Н-холинорецепторы (никотиновые) и М-холинорецепторы (мускариновые).
Н-холинорецепторы в периферических отделах вегетативной нервной системы расположены в ганглионарных синапсах симпатического и парасимпатического отделов, в каротидных клубочках и клетках мозгового слоя надпочечников. Возбуждение этих холинорецепторов сопровождается соответственно облегчением проведения возбуждения через ганглии, что ведет к повышению тонуса симпатического и парасимпатического отделов вегетативной нервной системы; повышением рефлекторного возбуждения дыхательного центра, в результате чего углубляется дыхание; повышением секреции адреналина.
Н-холинорецепторы, локализованные на клетках скелетных мышц (в нервно-мышечном синапсе). Н-холинорецепторы подразделили на Н-холинорецепторы ганглионарного типа (Нн-холинорецепторы) и мышечного тина (Нм-холинорецепторы).
М-холипорецепторы находятся на обкладочных клетках желудочных желез и их возбуждение приводит к усилению секреции соляной кислоты. М2-холинорецепторы располагаются в проводящей системе сердца. Возбуждение этих рецепторов приводит к понижению концентрации цАМФ, открытию калиевых каналов и увеличению тока К+, что приводит к гиперполяризации и тормозным эффектам: брадикардии, замедлению атрио-вентрикулярной проводимости, ослаблению сокращений сердца, понижению потребности сердечной мышцы в кислороде.
М3-хо-линорецепторы локализованы в основном в гладких мышцах некоторых внутренних органов и экзокринных железах. Взаимодействие ацетилхолина с этими рецепторами приводит к активации натриевых каналов, деполяризации, формированию ВПСП, вследствие чего клетки возбуждаются и происходит сокращение гладких мышц и выделение соответствующих секретов.
НА обеспечивает химическую передачу нервного импульса в норадренергических синапсах вегетативной нервной системы.
Действие норадреналина на клетку опосредуется адренорецепторами. Адренорецепторы находятся в различных тканях организма и воспринимают действие норадреналина и адреналина. Адренорецепторы делят на α-адренорецепторы и β-адренорецепторы, а в пределах этих классов выделяют α1-, α2-, β1-, β2- и β3-адренорецепторы. На одной и той же клетке могут располагаться различные адренорецепторы.
Конечный эффект возбуждения симпатических волокон зависит от того, какие адренорецепторы преобладают в органе.
Возбуждение α1-адренорецепторов приводит к:
сужению радиальной мышцы глаза и расширению зрачка (мидриаз), сужению соответствующих сосудов и повышению АД, сокращению капсулы селезенки и выбросу депонированной крови, сокращению сфинктеров пищеварительного тракта и мочевого пузыря, расслаблению гладких мышц кишечника и снижению его перистальтики и т.д.
Среди α2-адренорецепторов выделяют пре-, пост- и внесинаптические. Возбуждение пресинаптических α2-адренорецепторов по механизму отрицательной обратной связи уменьшает выделение норадреналина при его избытке в синаптической щели. Постсинаптические α2-адренорецепторы находятся в бета-клетках поджелудочной железы. Их возбуждение вызывает угнетение выброса инсулина в кровь. Внесинаптические α2-адренорецепторы обнаружены преимущественно на мембране тромбоцитов, эндотелии некоторых сосудов, в жировых клетках. Возбуждение этих рецепторов вызывает сужение сосудов, агрегацию тромбоцитов, угнетение липолиза.
Β1-адренорецепторы (постсинаптические) выявлены в основном в проводящей системе сердца и гладкой мышце кишечника. Их возбуждение приводит к:
увеличению частоты сердечных сокращений,
повышению проводимости и сократимости сердечной мышцы,
увеличению потребности сердца в кислороде,
понижению тонуса и моторной активности кишечника.
Стимуляция пресинаптических β2-адренорецепторов по механизму положительной обратной связи вызывает выделение норадреналина при его недостатке в синаптической щели. Постсинаптические β2-адренорецепторы расположены в основном в эндотелии сосудов скелетных мышц, головного мозга, легких, коронаров, а также в гладкой мускулатуре бронхов, матки и на гепатоцитах.
ВОЗБУЖДЕНИЕ Β2-АДРЕНОРЕЦЕПТОРОВ вызывает :
расширение соответствующих сосудов и понижение АД,
расслабление бронхов и матки,
усиление в печени гликогенолиза за счет активации цАМФ-зависимой фосфорилазы
повышение в крови сахара.
Β3-Адренорецепторы находятся в жировых клетках. Их стимуляция приводит к активации липолиза.
Дофамин. Осуществляет химическую передачу нервных импульсов не только в дофаминергических синапсах ЦНС, но и во вставочных нейронах симпатических ганглиев и во внутриорганном отделе вегетативной нервной системы.
Д-рецепторы выявлены на гладкомышечных клетках кишечника, сосудов почек, аорты, паращиторидных железах, канальцах почек. Возбуждение этих рецепторов приводит к расслаблению гладких мышц, понижению тонуса кишечника, расширению соответствующих сосудов,
Гистамин. Наибольшее количество его находится в постганглионарных симпатических волокнах. -рецепторы локализуются в гладкой мускулатуре бронхов, желудочно-кишечного тракта, сосудов, в сердце (атриовентрикулярный узел).
Возбуждение -рецепторов сопровождается: дз
· повышением тонуса и перистальтики кишечника
· сужением крупных сосудов, но расширением артериол, венул и развитием, в общем итоге, гипотензии
· повышением сосудистой проницаемости
· уменьшением времени проведения по атриовентрикулярному узлу
· увеличением образования простагландинов
Возбуждение -рецепторов приводит к:
повышению секреции кислоты в желудке и секреции бронхиальных желез
уменьшению высвобождения гистамина базофилами
Эфферентная (центробежная) иннервация включает:
1) вегетативные нервы всех внутренних органов, кровеносных сосудов, желез внешней секреции;
2) двигательные (соматические) нервные волокна скелетной мускулатуры.
Центральные отделы вегетативной нервной системы представлены в спинном, продолговатом и головном мозге. Эфферентный путь вегетативной нервной системы прерывается в нервных узлах - ганглиях. Они делят вегетативные волокна на преганглионарные, осуществляющие нервную передачу из центров вегетативной нервной системы к ганглиям, и постганглионарные, проводящие нервные импульсы с ганглиев на исполнительные органы.
Вегетативная иннервация подразделяется на парасимпатическую часть и симпатическую.
Парасимпатические нервы начинаются от клеток (первый нейрон) стволовой части головного мозга и крестцовой части спинного мозга (преганглионарные волокна) и заканчиваются в интрамуральных (расположенных внутри органов) ганглиях (второй нейрон), откуда короткие аксоны (постганглионарные волокна) достигают исполнительных клеток.
Симпатические нервы исходят из клеток боковых рогов грудного и поясничного отделов спинного мозга (первый нейрон) и прерываются в симпатических ганглиях (второй нейрон), аксоны (постганглионарные волокна) которых заканчиваются на клетках исполнительных органов и тканей.
Синапсы (от греческого synopsis -соединение, "смыкать") - место контакта аксона (нервного окончания) и иннервируемого им ганглия, органа или мышечного волокна. Контакт реализуется химическим посредником - медиатором (ацетилхолином или норадреналином).
Преганглионарные волокна парасимпатической и симпатической нервной системы выделяют ацетилхолин, и синаптическая передача во всех ганглиях осуществляется с помощью ацетилхолина. Постганглионарные нервные окончания парасимпатической нервной системы выделяют ацетилхолин, следовательно в синапсах, образованных этими окончаниями и клетками органов (постганглионарные синапсы парасимпатической нервной системы), медиатором является ацетилхолин.
В постганглионарных синапсах симпатической нервной системы передача осуществляется за счет медиатора норадреналина. Таким
образом, вегетативные нервы включают два нейрона (один расположен преганглионарно, другой - в составе ганглия), преганглионарные и постганглионарные волокна и два синапса - ганглионарный и постганглионарный.
Некоторые структуры организма (мозговой слой надпочечников, синокаротидная зона) эмбрионально являются симпатическими ганглиями и получают только преганглионарную импульсацию (от боковых рогов спинного мозга) при участии медиатора ацетилхолина. Последний стимулирует, например, хромаффинные клетки надпочечников, выделяющие адреналин.
Двигательные соматические нервы являются аксонами нервных клеток, расположенных в передних рогах спинного мозга; заканчиваются они в скелетной мускулатуре. Медиатор нервно-мышечного синапса - ацетилхолин.
При исследовании различных синапсов выяснена однотипность их строения. В синапсе различают пресинаптическую мембрану (поверхность нервного окончания, обращенная к иннервируемому органу или мышце) и постсинаптическую мембрану. Участок клеток органа или мышцы постсинаптической мембраны, воспринимающий импульс нейрона, называется рецептором. Пресинаптическую и постсинаптическую мембраны разделяет синаптическая щель, куда выделяется медиатор. Медиатор синтезируется в цитоплазме нервного окончания и депонируется в синаптических пузырьках (везикулах). При поступлении импульса медиатор выбрасывается в синаптическую щель, взаимодействует с рецепторами постсинаптической мембраны, а затем инактивируется (разрушается).
Эфферентные нервы и синапсы делятся на холинергические и адренергические, в зависимости от синтезирующегося в окончаниях и выделяющегося в синаптическую щель медиатора.
Холинергическими являются все двигательные, все преганглионарные (парасимпатические и симпатические), постганглионарные парасимпатические волокна, а также постганглионарные симпатические нервные волокна, иннервирующие потовые железы.
К адренергическим относятся только постганглионарные симпатические нервные волокна (исключая иннервацию потовых желез). Средства, влияющие на передачу импульса в холинергических синапсах, называют холинергическими, в адренергических - адренергическими.
Ганглии симпатического отдела вегетативной нервной системы в зависимости от их локализации разделяют на вертебральные (иначе их называют паравертебральными) и превертебральные. Ганглии парасимпатического отдела вегетативной нервной системы расположены внутри органов или вблизи них.
Вегетативные ганглии имеют большую продолжительность ВПСП. Длительный период следовой гиперполяризации, поэтому легко возникает торможение после возбуждения. Очень малая скорость проведения возбуждения - в 5-10 раз больше, чем в центральной нервной системе. Нейроны вегетативных ганглиев характеризуются низкой лабильностью, пропуская из центральной нервной системы на периферию малое количество импульсов. При частоте 100 импульсов в секунду в вегетативных ганглиях возникает полный блок. Таким образом, вегетативные ганглии - автономное образование, регулирующее проведение импульсов к рабочим органам.
Феномен мультипликации (умножения) в вегетативном ганглии - в вегетативном ганглии одновременно выражены феномены конвергенции и дивергенции импульсов: на теле одного постганглионарного нейрона конвергируют импульсы от нескольких преганглионарных нейронов и любой преганглионарный нейрон иннервирует много постганглионарных нейронов. Это обеспечивает надёжность передачи возбуждения. Вегетативные ганглии играют важную роль в распределении и распространении проходящих через них нервных влияний. Число нервных клеток в ганглиях в несколько раз больше числа приходящих к ганглию преганглионарных волокон. Каждое из этих волокон сильно ветвится и образует синапсы на многих клетках ганглия. Поэтому нервные импульсы, поступающие по преганглионарному волокну в ганглий, могут оказывать влияние на большое число ганглионарных нейронов и, следовательно, на еще большее число мышечных и железистых клеток иннервируемого органа. Таким образом, достигается расширение зоны влияния преганглионарных волокон.
Адренергические синапсы. В адренергических синапсах передача возбуждения осуществляется посредством норадреналина. В пределах периферической иннервации норадреналин принимает участие в передаче импульсов с адренергических волокон на эффекторные клетки. Адренергические аксоны, подходя к эффектору, разветвляются на тонкую сеть волокон с варикозными утолщениями, выполняющими функцию нервных окончаний. Последние участвуют в образовании синаптических контактов с эффекторными клетками. В варикозных утолщениях находятся везикулы (пузырьки), содержащие медиатор норадреналин. Биосинтез норадреналина осуществляется в адренергических нейронах из тирозина с участием ряда энзимов. Образование ДОФА и дофамина происходит в цитоплазме нейронов, а норадреналина — в везикулах.
В ответ на нервные импульсы происходит высвобождение норадреналина в синаптическую щель и последующее его взаимодействие с адренорецепторами постсинаптической мембраны. Существующие в организме адренорецепторы обладают неодинаковой чувствительностью к химическим соединениям.
Основываясь на этом, выделяют:
- Альфа-1и бета-1рецепторы локализуются в основном на постсинаптических мембранах и реагируют на действие норадреналина, выделяющегося из нервных окончаний постганглионарных нейронов симпатического отдела.
- Альфа-2и бета-2 рецепторы являются внесинаптическими, а также имеются на пресинаптической мембране тех же нейронов. На альфа-2 рецепторы действуют как адреналин, так и норадреналин. Бета-2 рецепторы чувствительны в основном к адреналину. На альфа-2 рецепторы пресинаптической мембраны норадреналин действует по принципу отрицательной обратной связи - ингибирует собственное выделение. При действии адреналина на бета-2 адренорецепторы пресинаптической мембраны выделение норадреналина усиливается. Поскольку адреналин выделяется из мозгового слоя надпочечников под действием норадреналина, возникает петля положительной обратной связи.
Холинергические синапсы. Медиатор холинергического синапса - ацетилхолин - синтезируется в нервном окончании из ацетилкоэнзима-А и холина и накапливается в пузырьках у пресинаптической мембраны. Под действием нервного импульса пузырьки лопаются, и ацетилхолин высвобождается в синаптическую щель. Далее он посредством диффузии достигает постсинаптической мембраны и возбуждает холинорецепторы, находящиеся на ней, что и обеспечивает контакт. В конечном итоге все выделенные в синаптическую щель молекулы ацетилхолина расщепляются до холина и уксусной кислоты при помощи специфического фермента ацетилхолинэстеразы, что прекращает активирующее действие медиатора на холинорецепторы. Активность ацетилхолинэстеразы настолько велика, что период полужизни ацетилхолина в синаптической щели измеряется в миллисекундах.
Рецепторы ацетилхолина на постсинаптической мембране (холинорецепторы) неоднородны, они разделяются на два больших класса в зависимости от чувствительности к двум природным алкалоидам - мускарину и никотину. Выделяют М-холинорецепторы, которые специфически активируются мускарином и блокируются атропином, и Н-холинорецепторы, которые специфически активируются малыми концентрациями никотина и блокируются большими концентрациями никотина. Для понимания эффектов препаратов, влияющих на холинергические процессы, важно знать локализацию М- и Н-холинорецепторов в организме. Препараты, влияющие на холинергические процессы, могут быть разделены на два больших класса:
1) препараты, активирующие холинорецепторы, т.е. влияющие подобно самому ацетилхолину, и поэтому они называются холиномиметиками.
2) препараты, блокирующие холинорецепторы, т.е. препятствующие действию ацетилхолина, и они называются холиноблокаторами.
Читайте также:
