
Перехваты ранвье в миелиновом нервном волокне это
Состоят из отростка нервной клетки, покрытого оболочкой, которая формируется олигодендроцитами. Отросток нервной клетки (аксон или дендрит) в составе нервного волокна называется осевым цилиндром.
- безмиелиновое (безмякотное) нервное волокно,
- миелиновое (мякотное) нервное волокно.
Безмиелиновые нервные волокна
Находятся преимущественно в составе вегетативной нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи, в которых на определенном расстоянии друг от друга видны овальные ядра. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько (10—20) осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в смежное. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. При электронной микроскопии безмиелиновых нервных волокон видно, что по мере погружения осевых цилиндров в тяж неиролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне
29. Миелиновые нервные волокна
1) внутренний, более толстый, — миелиновый слой,
2) наружный, тонкий, состоящий из цитоплазмы, ядер нейролеммоцитов и нейролеммы.
Миелиновый слой содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии — насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы видны участки волокна, лишенные миелинового слоя, — узловатые перехваты, или перехваты Ранвье, т.е. границы между соседними леммоцитами.
Отрезок волокна между смежными перехватами называется межузловым сегментом.
В процессе развития аксон погружается в желобок на поверхности нейролеммоцита. Края желобка смыкаются. При этом образуется двойная складка плазмолеммы нейролеммоцита — мезаксон. Мезаксон удлиняется, концентрически наслаивается на осевой цилиндр и образует вокруг него плотную слоистую зону — миелиновый слой. Цитоплазма с ядрами отодвигается на периферию – образуется наружная оболочка или светлая Шванновская оболочка (при окраске осмиевой кислотой).
Осевой цилиндр состоит из нейроплазмы, продольных параллельных нейрофиламентов, митохондрий. С поверхности покрыт мембраной – аксолеммой, обеспечивающей проведение нервного импульса. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Нервный импульс в миелиновом нервном волокне проводится как волна деполяризации цитолеммы осевого цилиндра, "прыгающая" (сальтирующая) от перехвата к следующему перехвату со скоростью до 120 м/сек.
В случае повреждения только отростка нейроцита регенерация возможна и протекает успешно при наличии определенных для этого условий. При этом, дистальнее места повреждения осевой цилиндр нервного волокна подвергается деструкции и рассасывается, но леммоциты при этом остаются жизнеспособными. Свободный конец осевого цилиндра выше места повреждения утолщается - образуется "колба роста", и начинает расти со скоростью 1 мм/день вдоль оставшихся в живых леммоцитов поврежденного нервного волокна, т.е. эти леммоциты играют роль "проводника" для растущего осевого цилиндра. При благоприятных условиях растущий осевой цилиндр достигает бывшего рецепторного или эффекторного концевого аппарата и формирует новый концевой аппарат.
30. шванновские клетки (леммоциты) — вспомогательные клетки нервной ткани, которые формируются вдоль аксонов периферических нервных волокон. Создают, а иногда и разрушают, электроизолирующую миелиновую оболочку нейронов. Выполняют опорную (поддерживают аксон) и трофическую (питают тело нейрона) функции. Описаны немецким физиологом Теодором Шванном в 1838 году и названы в его честь.
Каждое периферическое нервное волокно одето тонким цитоплазматическим слоем — невролеммой или шванновской оболочкой. Волокно является миелинизированным, если между ним и цитоплазмой шванновской клетки имеется значительный слой миелина. Если волокна лишены миелина, то они называются немиелинизированными безмякотными. Шванновские клетки могут осуществлять волнообразные движения, что, вероятно, способствует транспортировке различных веществ по отросткам нервных клеток.
С нарушением работы шванновских клеток связаны такие нервные заболевания, как синдром Гийена — Барре, болезнь Шарко-Мари, шванноматозис и хроническая воспалительная демиелинизирующая полинейропатия. Демиелинизация в основном происходит из-за ослабления двигательных функций шванновских клеток, в результате чего они оказываются не способны образовывать миелиновую оболочку.
Особенности распространения возбуждения по безмиелиновым волокнам:
1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением.
2. Возбуждение распространяется с небольшой скоростью.
3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна).
По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров.
Однако низкая скорость распространения возбуждения и его затухание не всегда выгодно организму. Поэтому природой был выработан еще один дополнительный механизм распространения возбуждения.
32. Механизмы проведения возбуждения в миелиновых волокнах. Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. В миелинизированномволокне токи проводятся только в зонах, не покрытых миелином (перехватах Ранвье). В этих участках генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением.
При действии раздражителя пороговой силы на мембрану миелинового волокна в области перехвата Ранвье изменяется проницаемость для ионов Nа + , которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны, что ведет к возникновению круговых токов. Этот ток идет через межтканевую жидкость к соседнему перехвату, где происходит смена заряда. Таким образом, возбуждение перепрыгивает с одного участка на другой. Обратное движение возбуждения невозможно так как участок, через который оно прошло, находится в фазе абсолютной рефрактерности.
Особенности распространения возбуждения по миелиновым волокнам:
2. Возбуждение распространяется с большой скоростью.
3. Возбуждение распространяется без декримента.
По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции.
Сальтаторное проведение
(лат. saltatorius, от salto — скачу, прыгаю)
Сальтаторное распространение возбуждения в миелинизированном нервном волокне от перехвата к перехвату [стрелками показано направление тока, возникающего между возбуждённым (А) и соседним покоящимся (Б) перехватом].
34. Проведение нервного импульса, передача сигнала в виде волны возбуждения в пределах одного нейрона и от одной клетки к другой. П. н. и. по нервным проводникам происходит с помощью электротонических потенциалов и потенциалов действия, которые распространяются вдоль волокна в обоих направлениях, не переходя на соседние волокна (см. Биоэлектрические потенциалы, Импульс нервный). Передача межклеточных сигналов осуществляется через синапсы чаще всего с помощью медиаторов, вызывающих появление потенциалов постсинаптических. Нервные проводники можно рассматривать как кабели, обладающие относительно низким осевым сопротивлением (сопротивление аксоплазмы — ri) и более высоким сопротивлением оболочки (сопротивление мембраны — rm). Нервный импульс распространяетсявдоль нервного проводника посредством прохождения тока между покоящимися и активными участками нерва (локальные токи). В проводнике по мере увеличения расстояния от места возникновения возбуждения происходит постепенное, а в случае однородной структуры проводника экспоненциальное затухание импульса, который в 2,7 раза уменьшается на расстоянии l = (константа длины). Так как rm и riнаходятся в обратном отношении к диаметру проводника, то затухание нервного импульса в тонких волокнах происходит раньше, чем в толстых. Несовершенство кабельных свойств нервных проводников восполняется тем, что они обладают возбудимостью. Основное условие возбуждения — наличие у нервов потенциала покоя. Если локальный ток через покоящийся участок вызовет деполяризацию мембраны, достигающую критического уровня (порога), это приведёт к возникновению распространяющегося потенциала действия (ПД). Соотношение уровня пороговой деполяризации и амплитуды ПД, обычно составляющее не менее 1: 5, обеспечивает высокую надёжность проведения: участки проводника, обладающие способностью генерировать ПД, могут отстоять друг от друга на таком расстоянии, преодолевая которое нервный импульс снижает свою амплитуду почти в 5 раз. Этот ослабленный сигнал будет снова усилен до стандартного уровня (амплитуда ПД) и сможет продолжить свой путь по нерву.
Основной функцией аксонов является проведение импульсов, возникающих в нейроне. Аксоны могут быть покрыты миелиновой оболочкой (миелиновые волокна) или лишены ее (безмиелиновые волокна). Миелиновые волокна чаще встречаются в двигательных нервах, безмиелиновые преобладают в автономной (вегетативной) нервной системе.
Отдельное миелиновое нервное волокно состоит из осевого цилиндра, покрытого миелиновой оболочкой, образованной шванновскими клетками. Осевой цилиндр имеет мембрану и аксоплазму. Миелиновая оболочка является продуктом деятельности шванновской клетки и состоит на 80% из липидов, обладающих высоким омическим сопротивлением, и на 20% из белка.
Миелиновая оболочка не покрывает сплошным покровом осевой цилиндр, а прерывается, оставляя открытые участки осевого цилиндра, называемые узловыми перехватами (перехваты Ранвье). Длина участков между этими перехватами различна и зависит от толщины нервного волокна: чем оно толще, тем длиннее расстояние между перехватами (рис. 2.17).
Безмиелиновые нервные волокна покрыты только шванновской оболочкой.
Проведение возбуждения в безмиелиновых волокнах отличается от такового в миелиновых волокнах благодаря разному строению оболочек. В безмиелиновых волокнах возбуждение постепенно охватывает соседние участки мембраны осевого цилиндра и так распространяется до конца аксона. Скорость распространения возбуждения по волокну определяется его диаметром.
В нервных безмиелиновых волокнах, где процессы метаболизма не обеспечивают быструю компенсацию расхода энергии на возбуждение, распространение этого возбуждения идет с постепенным ослаблением — с декрементом. Декрементное проведение возбуждения характерно для низкоорганизованной нервной системы.
У высших животных благодаря прежде всего наличию миелиновой оболочки и совершенства метаболизма в нервном волокне возбуждение проходит, не затухая, бездекрементно. Этому способствуют наличие на всем протяжении мембраны волокна равного заряда и быстрое его восстановление после прохождения возбуждения.
В миелиновых волокнах возбуждение охватывает только участки узловых перехватов, т. е. минует зоны, покрытые миелином. Такое проведение возбуждения по волокну называется сальтаторным (скачкообразным). В узловых перехватах количество натриевых каналов достигает 12 000 на 1 мкм , что значительно больше, чем в любом другом участке волокна. В результате узловые перехваты являются наиболее возбудимыми и обеспечивают большую скорость проведения возбуждения. Время проведения возбуждения по миелиновому волокну обратно пропорционально длине между перехватами.
Проведение возбуждения по нервному волокну не нарушается в течение длительного (многочасового) времени. Это свидетельствует о малой утомляемости нервного волокна. Считают, что нервное волокно относительно неутомляемо вследствие того, что процессы ресинтеза энергии в нем идут с достаточно большой скоростью и успевают восстановить траты энергии, происходящие при прохождении возбуждения.
В момент возбуждения энергия нервного волокна тратится на работу натрий-калиевого насоса. Особенно большие траты энергии происходят в перехватах Ранвье вследствие большой плотности здесь натрий-калиевых каналов.
Дж. Эрлангер и X. Гассер (1937) впервые классифицировали нервные волокна пс скорости проведения возбуждения. Различная скорость проведения возбуждения по волокнам смешанного нерва вы является при использовании внеклеточного электрода. Потенциалы волокон, проводящих возбуждение с неодинаковой скоростью, регистрируются раздельно (рис. 2.18).
В зависимости от скорости проведения возбуждения нервные волокна делят на три типа: А, В, С. В свою очередь волокна типа А подразделяют на четыре группы: Аα, Aβ, Aγ, Aδ. Наибольшей скоростью проведения (до 120 м/с) обладают волокна группы Аα, которую составляют волокна диаметром 12—22 мкм. Другие волокна имеют меньший диаметр и соответственно проведение возбуждения по ним происходит с меньшей скоростью (табл. 2.4).
Нервный ствол образован большим числом волокон, однако возбуждение, идущее по каждому из них, не передается на соседние. Эта особенность проведения возбуждения по нерву носит название закона изолированного проведения возбуждения по отдельному нервному волокну. Возможность такого проведения имеет большое физиологическое значение, так как обеспечивает, например, изолированность сокращения каждой нейромоторной единицы.
Таблица- Скорость проведения возбуждения по нервным волокнам
Нервная система человека и позвоночных животных имеет единый план строения и представлена центральной частью – головным и спинным мозгом, а также периферическим отделом – отходящими от центральных органов нервами, представляющими собой отростки нервных клеток – нейронов.
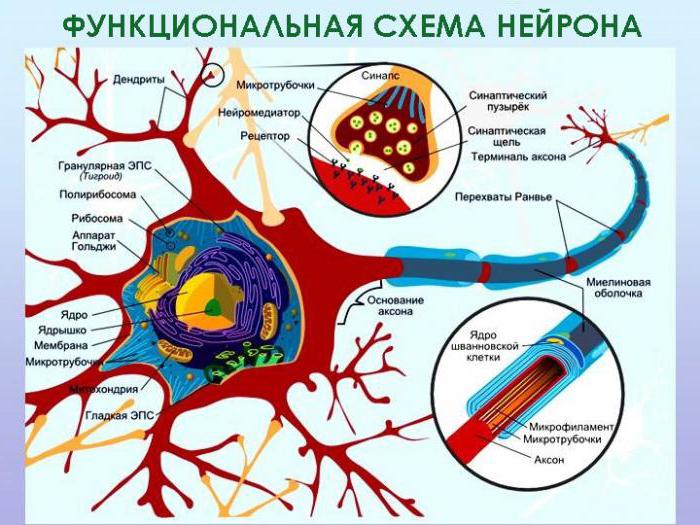
Их совокупность образует нервную ткань, главными функциями которой является возбудимость и проводимость. Эти её свойства объясняются прежде всего особенностями строения оболочек нейронов и их отростков, состоящих из вещества, названного миелином. В данной статье мы рассмотрим строение и функции этого соединения, а также выясним возможные способы его восстановления.
Почему нейроциты и их отростки покрыты миелином
Совсем не случайно дендриты и аксоны имеют защитный слой, состоящий из белково-липидных комплексов. Дело в том, что возбуждение является биофизическим процессом, в основе которого лежат слабые электрические импульсы. Если электрический ток идёт по проводу, то последний должен быть покрыт изоляционным материалом, чтобы уменьшить рассеивание электрических импульсов и не допустить снижение силы тока. Такие же функции в нервном волокне выполняет миелиновая оболочка. Кроме того, она является опорой, а также обеспечивает питание волокна.
Химический состав миелина
Как и большинство клеточных мембран, он имеет липопротеидную природу. Причём содержание жиров здесь очень высокое – до 75%, а белков – до 25%. Миелин в незначительном количестве содержит также гликолипиды и гликопротеиды. Химический состав его различается в спинномозговых и в черепно-мозговых нервах.
В первых наблюдается высокое содержание фосфолипидов – до 45%, а остальная часть приходится на холестерин и цереброзиды. Демиелинизация (то есть замена миелина на другие вещества в нервных отростках) приводит к таким тяжёлым аутоиммунным заболеваниям, как, например, рассеянный склероз.
С химической точки зрения, этот процесс будет выглядеть так: миелиновая оболочка нервных волокон меняет свою структуру, что проявляется прежде всего в уменьшении процентного содержания липидов по отношению к белкам. Далее снижается количество холестерина и возрастает содержание воды. А всё это приводит к постепенной замене миелина, содержащего олигодендроциты или шванновские клетки на макрофаги, астроциты и межклеточную жидкость.
Результатом таких биохимических изменений будет резкое снижение способности аксонов проводить возбуждение вплоть до полной блокировки прохождения нервных импульсов.
Особенности нейроглиальных клеток
Как мы уже говорили, миелиновая оболочка дендритов и аксонов образована специальными структурами, характеризующимися низкой степенью проницаемости для ионов натрия и кальция, а потому имеющих только потенциалы покоя (они не могут проводить нервные импульсы и выполняют электроизоляционные функции).
Данные структуры называются глиальными клетками. К ним относятся:
- олигодендроциты;
- волокнистые астроциты;
- клетки эпендимы;
- плазматические астроциты.
Все они формируются из наружного слоя зародыша – эктодермы и имеют общее название – макроглия. Глия симпатических, парасимпатических и соматических нервов представлена шванновскими клетками (нейролеммоцитами).
Строение и функции олигодендроцитов
Они входят в состав центральной нервной системы и являются клетками макроглии. Так как миелин – это белково-липидная структура, она способствует увеличению скорости проведения возбуждения. Сами клетки образуют электроизолирующий слой нервных окончаний в головном и спинном мозге, формируясь уже в период внутриутробного развития. Их отростки обворачивают в складки своей наружной плазмалеммы нейроны, а также дендриты и аксоны. Получается, что миелин – это основной электроизолирующий материал, разграничивающий нервные отростки смешанных нервов.
Шванновские клетки и их особенности
Миелиновая оболочка нервов периферической системы образована нейролеммоцитами (шванновскими клетками). Их отличительная особенность состоит в том, что они способны образовывать защитную оболочку только одного аксона, и не могут формировать отростки, как это присуще олигодендроцитам.
Между шванновскими клетками на расстоянии 1-2 мм располагаются участки, лишённые миелина, так называемые перехваты Ранвье. По ним скачкообразно происходит проведение электрических импульсов в пределах аксона.
Леммоциты способны к репарации нервных волокон, а также выполняют трофическую функцию. В результате генетических аббераций клетки оболочки леммоцитов начинают неконтролируемое митотическое деление и рост, вследствие чего в различных отделах нервной системы развиваются опухоли – шванномы (невриномы).
Роль микроглии в разрушении миелиновой структуры
Микроглия представляет собой макрофаги, способные к фагоцитозу и умеющие распознавать различные патогенные частицы – антигены. Благодаря мембранным рецепторам эти глиальные клетки вырабатывают ферменты – протеазы, а также цитокины, например, интерлейкин 1. Он является медиатором воспалительного процесса и иммунитета.
Более того, цитокины, активируя рецепторы, провоцируют избыточный транспорт ионов кальция в тело нейрона. Протеазы и фосфолипазы начинают расщеплять органеллы и отростки нервных клеток, что приводит к апоптозу – гибели данной структуры.
Она разрушается, распадаясь на частицы, которые и пожирают макрофаги. Это явление называется эксайтотоксичностью. Оно вызывает дегенерацию нейронов и их окончаний, приводя к таким заболеваниям, как болезнь Альцгеймера и болезнь Паркинсона.
Мякотные нервные волокна
Если отростки нейронов – дендриты и аксоны, покрывает миелиновая оболочка, то они называются мякотными и иннервируют скелетную мускулатуру, входя в соматический отдел периферической нервной системы. Немиелинизированные волокна образуют вегетативную нервную систему и иннервируют внутренние органы.
Мякотные отростки имеют больший диаметр, чем безмякотные, и формируются следующим образом: аксоны прогибают плазматическую мембрану клеток глии и формируют линейные мезаксоны. Затем они удлиняются и шванновские клетки многократно обворачиваются вокруг аксона, образуя концентрические слои. Цитоплазма и ядро леммоцита перемещаются в область наружного слоя, который называется неврилеммой или шванновской оболочкой.
Внутренний слой леммоцита состоит из слоистого мезоксона и называется миелиновой оболочкой. Толщина её в различных участках нерва неодинакова.
Как восстановить миелиновую оболочку
Рассматривая роль микроглии в процессе демиелинизации нервов, мы установили, что под действием макрофагов и нейромедиаторов (например, интерлейкинов) происходит разрушение миелина, что в свою очередь приводит к ухудшению питания нейронов и нарушению передачи нервных импульсов по аксонам.
Данная патология провоцирует возникновение нейродегенеративных явлений: ухудшение когнитивных процессов, прежде всего памяти и мышления, появление нарушения координации движений тела и тонкой моторики.
В итоге возможна полная инвалидизация больного, которая возникает в результате аутоиммунных заболеваний. Поэтому вопрос о том, как восстановить миелин, в настоящее время стоит особенно остро. К таким способам относится прежде всего сбалансированная белково-липидная диета, правильный образ жизни, отсутствие вредных привычек. В тяжелых случаях заболеваний применяют медикаментозное лечение, восстанавливающее количество зрелых глиальных клеток – олигодендроцитов.
Молекулярная специфика в перехватах Ранвье
Organization and function of the nodal environs
Скоординированная дифференцировка аксона и его миелинизирующих клеток требует тесной взаимосвязи между нейронами и глией на самых ранних стадиях развития. Сигналы, передаваемые аксоном, регулируют пролиферацию, выживаемость и дифференцировку олигодендроцитов и Шванновских клеток и участвуют в детерминации толщины миелина. Реципрокные глиальные сигналы влияют на цитоскелет аксонов и аксонный транспорт и необходимы для выживаемости аксонов. В результате таких реципрокных связей миелинизированные волокна приобретают структурные признаки, позволяющие им максимизировать скорость проведения импульса. Одним из таких признаков является разделение мембраны аксона на отдельные молекулярные, структурные и функциональные домены. Такие домены включают перехваты Ранвье, paranodal junction (околоузловое соединение), juxtaparanodes (участок, расположенный за paranodal junction ) и internodal (межперехватный участок (РИС.1b и 2). Авторы сфокусировали внимание на молекулярных механизмах, лежащих в основе образования и поддержания этих уникальных доменов аксона, необходимых для нормального функционирования нервной системы.
The node of Ranvier
Узловые перехваты Ранвье представляют собой небольшие по длине, равномерно расположенные разрывы миелиновой оболочки. Интервал между перехватами Ранвье имеет длину примерно в 100 раз большую диаметра нервного волокна. Имеются некоторые различия в структурных характеристиках перехватов между центральной и периферической нервными системами. В периферических нервах весь миелинизированный участок аксона покрыт базальной оболочкой (базальным слоем) (basal lamina), а внешний слой (outer collar) шванновской клетки имеет микроворсинки, покрывающие узловые перехваты (РИС. 1b). Околоузловое пространство (т.е. пространство между аксолеммой и базальным слоем), содержащее микроворсинки, также заполнено волокнистым матриксом. В ЦНС нет basal lamina и контакт перехватов осуществляется через околоузловые астроциты (perinodal astrocyte=перинодальные астроциты), которые недавно были названы синантоцитами (synantocytes) [Butt, A. M., Kiff, J., Hubbard, P. & Berry, M. Synantocytes: new functions for novel NG2 expressing glia. J. Neurocytol. 31, 551-565. 2002].
The nodal axolemma. Перехваты Ранвье характеризуются высокой плотностью (>1200/ μm2) Na+ каналов, которые важны для образования потенциала действия во время сальтаторной проводимости. Voltage-gated Na+ каналы представляют собой мультимерные комплексы, состоящие из пора-формирующей (pore-forming) α субъединицы и одной или более вспомогательных βсубъединиц (РИС.2а). У млекопитающих αсубъединицы кодируются девятью генами α-(Scn1a–Scn9a, β – четырьмя генами для β -субъединиц (Scn1b–Scn4b). Перехваты Pанвье в ЦНС и периферической нервной системе (ПНС) содержат в основном Nav1.6. Кроме того, во многих перехватах Ранвье ЦНС найдены Nav1.2 и Nav1.8, тогда как Nav1.9 обнаружены лишь в некоторых узловых перехватах ПНС. Во время развития и ЦНС, и ПНС экспрессируются Nav1.2, которые позже замещаются на Nav1.6. Функциональное значение такого переключения пока неясно, но это, вероятно, позволяет нейронам адаптироваться к высоко-частотному firing. Кроме voltage-gated Na+ каналов в узловых перехватах было идентифицировано несколько других трансмембранных и цитоскелетных белков – молекулы клеточной адгезии (cell-adhesion molecules – CAMs) иммуноглобулинового (Ig) суперсемейства Nrcam и neurofascin-186 (Nf186), цитоскелетный адаптер ankyrin G и актин-связывающий белок spectrin βIV (Berghs, S. et al. βIV spectrin, a new spectrin localized at axon initial segments and nodes of ranvier in the central and peripheral nervous system. J. Cell Biol. 151, 985-1002. 2000). Недавно было обнаружено присутствие двух К+ каналов в узловых перехватах Ранвье – Kv3.1 и Kcnq2 (Devaux, J. et al. Kv3.1b is a novel component of CNS nodes.J. Neurosci 23, 4509-4518 (2003). Devaux, J. J., Kleopa, K. A., Cooper, E. C., Bennett, V. & Scherer, S. S. Anatomical and physiological evidence of KCNQ2 subunits at PNS and CNS nodes. Soc. Neurosci Abstr. 28, 368.8. 2003). Kv3.1 обнаружен в основном в перехватах крупных аксонов ЦНС и в небольшом числе перехватов ПНС. Kcnq2 локализован во всей ПНС и в большинстве узловых перехватов ЦНС.
The nodal gap, extracellular matrix and the glial membrane В ПНС узловой пробел (перехват) заполнен микроворсинками Шванновских клеток, происходящими из наружной оболочки клетки (РИС.1b). В проксимальной области микроворсинок мембраны двух соседних Шванновских клеток соединены tight junctions (плотным соединением), но эти соединения не изолируют узелковый пробел. Было обнаружено, что они проницаемы для пероксидазы хрена, прикладываемой снаружи нервного волокна. Три белка – erzin, radixin и moesin, так же как и erzin-связывающий белок EBP50 и Rho-AGTPasa локализованы в микроворсинках. Потенциально эти белки могут связывать актин-обогащенные микроворсинки с интегральными мембранными белками (integral membrane proteins). Кроме того, несколько белков внеклеточного матрикса (extracellular matrix (ECM) proteins) найдено в узловых пробелах под базальной оболочкой, включая hyaluronan-binding proteoglycan versican, tenascin-C и NG2 proteoglycan. Недавно было показано, что dystroglycan в избытке экспрессирующийся на abaxonal поверхности миелинизирующих Шванновских клеток также локализуется в перехватах. Специфическое удаление dystroglycan в Шванновских клетках приводит к дезорганизации микроворсинок, заметной редукции узловых Na+ каналов и к последующему нарушению проводимости нервного волокна.
В отличие от ПНС, в ЦНС отростки перинодальных астроцитов (perinodal astrocytes) (РИС.1b) контактируют с большинством узловых перехватов. Было показано, что узловой пробел в ЦНС включает несколько протеогликанов, в том числе tenascin и phosphacan. Узловые пробелы ЦНС также содержат versican-binding protein Bral1, продуцируемый нейронами. Функции этих белков пока неясны, однако предполагают, что благодаря высокому содержанию в них acidic disaccharides, они могут обеспечивать негативную среду, служащую в качестве внеклеточного резервуара Na+ в перинодальном пространстве. И tenascin-C, и tenascin-R связываются с Na+ каналами и изменяют их электрофизиологические свойства. Генетическое удаление tenascin-R ведет к снижению нервной проводимости, но не оказывает никакого эффекта на распределение Na+ каналов в узловых перехватах, указывая на то, что такое взаимодействие может стабилизировать узловые комплексы или регулировать активность каналов, но не требуется для первичной кластеризации этих каналов. Сообщалось также, что Na+ каналы связывают цитоплазматический конец (tail) и внеклеточный домен Rptpβ – рецепторную тирозин фосфатазу, о локализации которой в узловой аксолемме (nodal axolemma) пока не сообщалось. Более того, значимость этих взаимодействий для нормальной физиологии миелинизированных нервов пока неясна, т.к. распределение узловых Na+ каналов и скорость проводимости миелинизированных аксонов в ЦНС нормальны у Rptpβ-дефицитных мышей. Стоит отметить, что и tenascin-R, и Rptpβ также взаимодействуют с contactin и Nrcam, присутствующими в узловых перехватах ЦНС, указывая на возможное присутствие крупных макромолекулярных комплексов в перинодальном пространстве.
The axoglial paranodal junction
Function of the paranodal junction. Предполагают, что paranodal junction присоединяет миелиновую оболочку к аксону, чтобы отделить электрическую активность в перехватах Ранвье от межперехватного участка (internodal region), находящегося под компактной миелиновой оболочкой, и служит в качестве преграды (fence), которая ограничивает латеральную диффузию белков аксолеммы. Недавние исследования с использованием 4-х паранодальных мутантных мышей – мышей с отсутствием Caspr, contactin, Cgt и Cst, (все с отсутствием характерной перегородки (septa) в аксоглиальном соединении) – позволили прекратить проверку этих оригинальных идей. В ЦНС этих мутантов паранодальные петли (paranodal loops) дезорганизованы, имеется множество перекрывающихся и инвертированных петель, обращенных в сторону от аксона. В ПНС этих мутантов морфологические изменения менее выражены, возможно, благодаря присутствию базального слоя (basal lamina). Параузлы (paranodes) хорошо организованы, однако увеличено пространство между глиальной оболочкой и аксоном. Но даже в отсутствие перегородки (septa) паранональные петли (loops) все еще тесно прикреплены к аксону во многих участках ЦНС и ПНС, указывая на присутствие до сих пор не идентифицированных паранодальных компонентов, которые опосредуют аксо-глиальный контакт в этом участке. Вместе с электронно-микроскопическими данными, показавшими, что поперечные соединения (transverse bands) образуются позже во время миелинизации, эти исследования указывают на возможную роль перегородки (septa) в сохранении паранодальных петель (колец) для аксона в аксоглиальном соединении. В соответствии с этими наблюдениями, постепенное, зависимое от возраста открепление паранодальных колец от аксона наблюдали в ЦНС Caspr-null мышей (Rios, J. C. et al. Paranodal interactions regulate expression of sodium channel subtypes and provide a diffusion barrier for the node of Ranvier. J. Neurosci. 23, 7001-7011. 2003).
Отсутствие паранодальной перегородки (paranodal septa) у четырех паранодальных мутантов ведет к реорганизации мембраны аксона (РИС.3). У этих мутантов shaker-type К+ каналы, в норме присутствующие в juxtaparanodal области неправильно локализованы по отношению к паранодальной мембране аксона. Поэтому, возможно, что паранодальное перегородчатое соединение (paranodal septate junction) функционирует как барьер, который ограничивает передвижение К+ каналов из-под компактного миелина, отделяя их от Na+ каналов в узловых перехватах. В противоположность juxtaparanodal К+ каналам, нарушение паранодальной перегородки минимально влияет на распределение узловых Na+ каналов. Существует небольшое увеличение длины перехватов, сопровождаемое уменьшением мембранных частиц (particles) в узловой аксолемме, определяемое при FREEZE –FRACTURE электронной микроскопии. Это указывает на то, что paranodal septate junction не является необходимым для появления перехватов. Однако глиальное прикрепление в области paranodes в ЦНС необходимо для сохранения Na+ кластеризации в аксолемме.перехвата Ранвье
Juxtaparanode локализуется в короткой области сразу за paranodal junction (РИС.1b). При freeze-fracture электронной микроскопии эта область представляет собой нерегулярно расположенные частицы, которые в большей степени сконцентрированы вблизи paranodes и более диффузны по мере приближения к межперехватам (internodes). Эти частицы соответствуют скорее всего гетеромультимерам delayed rectifier K+ каналов семейства Shaker – Kv1.1, Kv1.2 и Kvβ2. В juxtaparanodal аксолемме эти каналы колокализуются и образуют комплекс с Caspr2, вторым членом Caspr семейства. Кроме того, в этом участке присутствует Kv1.6, преимущественно в небольших аксонах. Два других белка, обнаруженных в juxtaparanodes, представляют собой транзиторный аксональный glycoprotein-1 (Tag1) – GPI-связанный CAM, родственный contactin, и connexin 29 (Cx29), обнаруженый в глиальной мембране. Связь Caspr2 с К+ каналами опосредуется их карбокси-терминальной областью, вероятнее всего через неидентифицируемый PDZ DOMAIN-содержащий белок. И хотя один такой белок Psd95 локализован в juxtaparanodes и ассоциируется с К+ каналами, он, как оказалось, не участвует во взаимодействии этих каналов с Caspr2 или их накоплении в этой области. Два последних исследования показали, что Caspr2 и Tag1 формируют juxtaparanodal complex, состоящий из глиальной Tag1 молекулы и аксонального Caspr2/Tag1 гетеродимера (РИС.2с) [Poliak, S. et al. Juxtaparanodal clustering of Shaker-like K+ channels in myelinated axons depends on Caspr2 and TAG-1. J. Cell Biol. 162, 1149-1160 (2003)]. Этот комплекс необходим для аккумуляции К+ каналов в juxtaparanodes, поскольку прицельное нарушение Caspr2 или Tag1 приводит к значительному снижению juxtaparanodal аккумуляции этих каналов в аксонах как ЦНС, так и ПНС (РИС.3). Эти результаты указывают, что Caspr2 и Tag1 могут формировать скаффолды, способные позиционировать ионные каналы в специфических участках плазменной мембраны, и это напоминает механизм, работающий во время формирования синапсов.
Хотя никакой специфики соединений не наблюдали между глией и аксоном в области межперехватов, при freeze-fracture электронной микроскопии было обнаружено, что межперехватная аксолемма в ПНС содержит продольные нити (longitudinal strands) внутримембранных частиц, напоминающих нити, найденные в paranodes и в juxtaparanodal областях. Как показано на РИС. 1с и 1d, Caspr и contactin локализуются по всей межперехватной области в полосе, фланкированной К+ каналами и Caspr2, которая соединяет внутренний мезаксон (inner mesaxon) миелиновой оболочки и формирует периферическое кольцо ниже внутренней стороны SCHMIDT-LANTERMAN INCISURES. Эта линия, названная juxtamesaxonal и juxta-incisural (Scherer, S. S. & Arroyo, E. J. Recent progress on the molecular organization of myelinated axons. J. Peripher. Nerv. Syst. 7, 1-12. 2002), является прямым продолжением paranodes/juxtaparanodes. Следовательно, Nf155, Cx29 и Tag1 ограничены в дополнительном распространении на adaxonal мембране миелинизирующих Шванновских клеток. Эти находки указывают, что локализация аксонных белков в межперехватах диктуется миелиновой оболочкой, вероятно, посредством механизма, сходного с тем, который работает в paranode/juxtaparanode. Однако недавний анализ Caspr2-null мышей показал, что разные механизмы могут контролировать локализацию К+ каналов в juxtaparanodes и в juxtamesaxon. Молекулярная организация межперехватной области не изучена в миелинизированных нервах ЦНС.
Formation of the nodal environ
Читайте также:
