
Почему в физиологическом растворе клетки нервной
Наряду с нервными клетками выделяют глиальные клетки: 1)олигодендроциты.2) астроциты.3) шванновские клетки. 4) микроглиоциты
Глиальные клетки окружают нейроны и в некоторых местах соприкасаются с ними. Число глиальных клеток на порядок выше числа нейронов. Основную роль глиальные клетки играют в формировании миелиновых оболочек аксонов. Участки без миелиновых оболочек – перехваты Ренвье, которые играют большую роль в передаче возбуждения. При возбуждении нейроны захватыват ионы натрия и кальция и отдают ионы калия. В поддержании концентрации ионов К межклеточных щелей в допустимых пределах большую роль играют астроциты. Не имея каналов, они тем не менее высоко проницаемы для К, вбирают в себя его избыток, а по необходимости выделяют его в среду. Шванновские клетки работают аналогично. Астроциты связаны м/собой контактами и образуют синцитий, эти оболочки тоже проницаемы для ионов калия. Основная роль глиальных клеток: 1)поддержание ионного гомеостаза. 2)изоляция нервных клеток. 3)участие в обмене медиатов
11. Функциональные свойства рецепторов. Особенности возникновения возбуждения в первичночувствующих и вторичночувствующих рецепторах.
Рецепторы – специализированные чувствительные образования, воспринимающие и преобразующие раздражения из внешней и внутренней среды организма в специфическую активность нервной системы. Они образованы терминалями дендритов чувствительных нейронов.
Особенности:1) Энергия раздражитель служит для них стимулом к запуску процессов, совершаемых за счет энергии, которая накоплена в следствии обменных реакций в самой клетке. 2) Рецепторная клетка обладает на выходе электр энергией, обязательно передаваемой другим клеткам, которые сами не способны воспринимать энергию данного внешнего воздействия.
Главными свойствами рецепторов являются:1)Специфичность - это способность определенных рецепторов воспринимать только определенный вид энергии. 2) Низкий порог чувствительности - это способность рецептора приходить в состояние возбуждения при самом незначительном воздействии. 3)Адаптация - способность рецепторов "привыкать" к постоянно действующему стимулу.
По месту приложения раздражителя рецепторы являются первичночувствующими (такитильные, обонятельные, интерорецепторы) – мономодальны (слух, зрение); и вторичночувствующими (зрительные, вкусовые, слуховые, вестибулярные) – бывают и бимодальными (тактильное чувство+боль). Перв чувств. рецепторы трансформируют энергию стимула в нервн активность непосредственно в сенсорном нейроне, и по его аксону без промежуточных преобразований нерв активность передается сенсорному ядру (1-ый сенсорный уровень). Втор чувств рецепторы представляют собой высокоспециализированные эпителиальные клетки, к которым подходят нервные волокна (сенсорные волокна), образуя с клетками синаптические контакты. Нервн активность в нейроне возникает лишь после синаптического преобразования потенциала высокоспециализированных клеток, а не в самой нервн клетке.
Механизм преобразования сигналов в рецепторе. Каскадный процесс.1) первичные чувств рецепторы
1 – специфическое взаимодействие раздражителя с мембраной рецептора на молекулярном уровне. В итоге: изменение проницаемости мембраны в зоне взаимодействия раздражителя с мембраной. 2 – возникновение рецепторного потенциала в месте действия раздражителя. В результате увеличивается проницаемость для ионов. Натрий входит внутрь и образуется локальный ответ. 3 – электротоническое распространение рецепторного потенциала в направлении аксона нейрона. 4 – перекодировка рецепторного потенциала в ПД. Нерв импульс возникает когда деполяриз эффект рецепторного потенциала или суммация нескольких рецепторных потенциалов достигает КУД в зоне наибольшего приспособления для генерации ПД = спайк. 5 – проведение ПД по аксону нейрона в центростремит направлении 2) вторичный чувств рецептор
Осуществляется двумя видами клеток: рецепторные клетки (восприятие сигнала), нервн клетки. В мембране клеток воспринимателей нет электрогенных участков, в которых мог бы возникать ПД, таким образом место генерации ПД и возникновения специфич трансформирующих изменений разделены. Взаимодействие м/у клетками обеспечивается синапсами.
12. Адренергические и холинергические рецепторы клеток разных органов, физиологические эффекты возбуждения этих рецепторов.
Адренорецепторы — рецепторы к адренэргическим веществам. Реагируют на адреналин и норадреналин. Различают 4 группы рецепторов, которые несколько различаются по опосредуемым эффектам, локализации, а также аффинитету к различным веществам: альфа-1, альфа-2, бета-1 и бета-2 адренорецепторы. Альфа-1 и бета-1 рецепторы локализуются в основном на постсинаптических мембранах и реагируют на действие норадреналина, выделяющегося из нервных окончаний постганглионарных нейронов симпатического отдела. Альфа-2 и бета-2 рецепторы являются внесинаптическими, а также имеются на пресинаптической мембране тех же нейронов. На альфа-2 рецепторы действуют как адреналин, так и норадреналин. Бета-2 рецепторы чувствительны в основном к адреналину. На альфа-2 рецепторы пресинаптической мембраны норадреналин действует по принципу отрицательной обратной связи - ингибирует собственное выделение. При действии адреналина на бета-2 адренорецепторы пресинаптической мембраны выделение норадреналина усиливается. Поскольку адреналин выделяется из мозгового слоя надпочечников под действием норадреналина, возникает петля положительной обратной связи.
13. Рецепторный отдел сенсорных систем. Зависимость между силой раздражения и интенсивностью ощущения в рецепторах. ( Закон Вебера и Вебера –Фехнера).
1/30, а для зрения минимальная воспринимаемая разница в интенсивности света также не зависит от величины интенсивности и составляет
Нервные ткани в составе органов относятся к стабильным тканям, так как нейроны митозом не делятся. Физиологическая и репаративная регенерация происходит путем частичной полиплоидизации ядер, восстановления синапсов после их повреждения, роста поврежденных отростков, а главное — путем обновления химических и метаболических компонентов нейронов при внутриклеточном обмене веществ. На месте дефекта в нервной ткани разрастается нейроглия. Она является менее дифференцированной тканью, клетки которой способны делиться митозом. Описаны глиальные клетки, обладающие высокими потенциями к размножению и развитию. Эти клетки принимают активное участие в восстановительных процессах нервной ткани.
При повреждениях, приводящих к нарушению целостности нервных волокон (огнестрельные раны, разрывы), их периферические части распадаются на фрагменты осевых цилиндров и миелиновых оболочек, погибают и фагоцитируются макрофагами (уоллеровская дегенерация осевых цилиндров). В сохранившейся части нервного волокна начинается пролиферация нейролеммоцитов, формирующих цепочку (бюнгнеровская лента), вдоль которой происходит постепенный рост осевых цилиндров. Таким образом, нейролеммоциты являются источником факторов, стимулирующих рост осевого цилиндра. При отсутствии препятствий в виде очагов воспаления и соединительнотканных рубцов возможно восстановление иннервации тканей.
Регенерация нервных отростков идет со скоростью 2-4 мм в сутки. В условиях лучевого воздействия происходит замедление процессов репаративного гистогенеза, что обусловлено в основном повреждением нейролеммоцитов и клеток соединительной ткани в составе нерва. Способность нервных волокон к регенерации после повреждения при сохранении целостности тела нейрона используется в микрохирургической практике при сшивании дистального и проксимального отростков поврежденного нерва. Если это невозможно, то используют протезы (например, участок подкожной вены), куда вставляют концы поврежденных нервов (футлериз). Регенерацию нервных волокон ускоряет фактор роста нервной ткани — вещество белковой природы, выделенное из тканей слюнных желез и из змеиного яда.
Важнейшее свойство живого — отвечать комплексом изменений метаболизма, двигательной активности, размножения или гибели и др. на внешние и внутренние воздействия. Восприятие, трансформация и передача последних осуществляется рецепторно-трансдукторной системой клетки, элементы которой связаны с внутренней средой клетки. На клеточном уровне организации живого первично формируется каскад реакций, распространяющихся на тканевый, органный и организменный уровни.
Под реактивностью тканей с гистогенетических позиций следует понимать изменения основных закономерных сторон развития ткани — пролиферации, дифференцировки, интеграции клеток, межклеточных взаимодействий и других закономерных процессов гистогенеза, происходящих под действием внешних для ткани факторов. При самых разнообразных воздействиях (травмы, ожоги, стрессовые ситуации и т. п.) ткани, для которых в норме характерно клеточное обновление путем митоза, реагируют прежде всего понижением или повышением степени пролиферативной активности клеток. Угнетение митозов или, наоборот, "вспышки" митотической активности — непременные показатели реактивности таких тканей. В некоторых тканях результатом реактивного изменения пролиферации клеток являются эндомитоз и амитоз, образование двуядерных и многоядерных клеток.
Наряду с повышением уровня дифференцировки клеток, реактивность тканей может характеризоваться и явлениями дедифференцировки клеток. Дедифференцировка клеток — это упрощение их внутренней структуры, связанное с временным снижением уровня дифференцировки (специализации) клеток. Дедифференцированные клетки приобретают способность к пролиферации. В них активизируется синтез ДНК и общих неспецифических белков. Дедифференцировка как реактивно-приспособительное изменение клеток сопровождается увеличением относительных объемов ядер и ядрышек, увеличением числа свободных рибосом, исчезновением специальных органелл и включений в цитоплазме, редукцией мембран комплекса Гольджи, уменьшением числа митохондрий и другими признаками. Под дедифференцировкой следует понимать структурно-адаптационную реакцию клетки, а не обратное ее развитие.
При изменении функциональной нагрузки на ткани реактивно-приспособительные изменения клеток проявляются в метаболических сдвигах — трофических нарушениях типа гипертрофии, дистрофии, атрофии, гипо- или гиперсекреции и т. д.
При гипертрофии клеток наблюдается гиперплазия внутриклеточных структур (митохондрий, эндоплазматической сети, органелл) как морфологический эквивалент повышенной функциональной активности.
Реактивность тканей характеризуется также изменениями межклеточных взаимодействий. При действии повреждающих факторов в тканях может возникнуть состояние дезинтеграции клеток (например, нарушение межнейронных связей при интоксикациях, дискомплексации эпителиев при нарушении контактов между эпителиоцитами и т. д.). Как проявление реактивности тканей следует рассматривать программированную гибель клеток за счет активации внутренней программы самоуничтожения или ее запуска внешними стимулами, например факторами, вырабатываемыми клетками микроокружения.
Реактивные изменения тканей зависят, конечно, во многом от силы раздражителя — повреждающего фактора. Существенное значение в реактивности тканей имеет наследственная природа клеток самой ткани, так как каждая ткань отличается генетически детерминированным диапазоном изменчивости или нормы адаптивной реакции, обеспечивающей нормальное функционирование ткани. После воздействия повреждающих факторов реактивные изменения клеток могут выходить за пределы диапазона изменчивости, характерного для нормального функционирования ткани. Однако обычно резко выраженные вначале реактивные изменения клеток постепенно сглаживаются, и структурно-функциональные свойства их приближаются к норме, обеспечивая адаптацию ткани к функционированию в новых условиях.
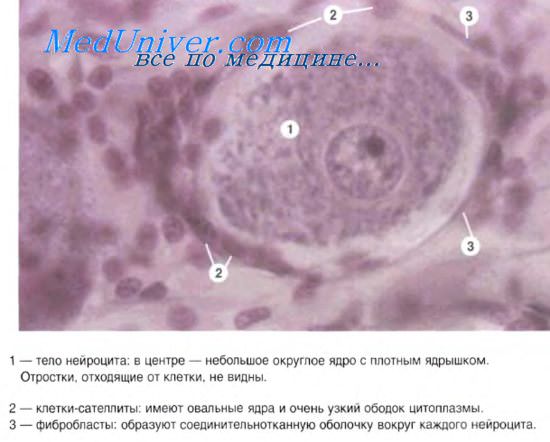
Структурно-функциональной единицей нервной ткани является нервная клетка – нейрон. Нейрон – специализированная клетка, которая способна принимать, кодировать, передавать и хранить информацию, устанавливать контакты с другими нейронами, организовывать ответную реакцию организма на раздражение.
Сколько нервных клеток в нашем мозге? Распространено мнение, что мозг человека состоит из ста миллиардов нейронов: это приблизительно столько же, сколько звезд в нашей Галактике. Число на слуху, правда, никто не знает, откуда оно взялось. В соответствии с имеющимися на сегодня лабораторными данными, в человеческом мозге содержится по разным данным от 15 до 86 миллиардов нейронов.
Формы нейронов чрезвычайно многообразны, не найдется и двух нейронов, одинаковых по виду. Несмотря на это, большинству нейронов присущи определенные структурные особенности, позволяющие выделить три области клетки: клеточное тело, дендриты и аксон. Тело содержит ядро и биохимический аппарат синтеза ферментов и других молекул, необходимых для жизнедеятельности клетки. Обычно тело нейрона имеет приблизительно сферическую или пирамидальную форму. Дендриты представляют собой тонкие трубчатые выросты, которые многократно делятся и образуют ветвистое дерево вокруг тела клетки. Они создают ту основную физическую поверхность, на которую поступают идущие к данному нейрону сигналы. Аксон тянется далеко от тела клетки и служит той линией связи, по которой сигналы, генерируемые в теле данной клетки, могут передаваться на большие расстояния в другие нейроны и исполнительные органы. Аксон отличается от дендритов как по строению, так и по свойствам своей наружной мембраны. Большинство аксонов длиннее и тоньше дендритов и имеет отличный от них характер ветвления: если отростки дендритов в основном группируются вокруг клеточного тела, то отростки аксонов располагаются на конце волокна, в том месте, где аксон взаимодействует с другими нейронами. Нейрон проводит возбуждение только от дендрита к аксону.
Таким образом, основные функции нейрона распределены следующим образом:
1) генерация и передача нервного импульса - передающая часть нейрона (аксональный холмик с аксоном);
2) получение информации - воспринимающая часть (дендриты и мембрана сомы нейрона);
3) способность суммировать возбуждающие и тормозящие сигналы - интегративная часть (тело нейрона).
Основными свойствами нейрона являются раздражимость и возбудимость.
Раздражимость, т.е. способность клетки организма реагировать, то есть изменять свои физиологические свойства и проявлять функциональные отправления в ответ на действие раздражителей. Раздражители – это факторы внешней или внутренней среды, вызывающие раздражение возбудимых структур.
Различают две группы раздражителей:
1) естественные (нервные импульсы, возникающие в нервных клетках и различных рецепторах);
2) искусственные: физические (механические – удар, укол; температурные – тепло, холод; электрический ток – переменный или постоянный), химические (кислоты, основания, эфиры и т.п.), физико-химические (осмотические – кристаллик хлорида натрия).
По биологическому принципу раздражителей делятся на:
1) адекватные, которые при минимальных энергетических затратах вызывают возбуждение ткани в естественных условиях существования организма;
2) неадекватные, которые могут вызвать возбуждение при достаточной силе и продолжительном воздействии.
Второе свойство нейронов, которое обеспечивает нормальное функционирование нервной системы – возбудимость. Возбудимость – свойство клеток отвечать на действие раздражителя специфическими изменениями ионной проницаемости и мембранного потенциала.
В двойной липидный слой мембраны встроены белки, которые выполняют несколько функций:
1) белки-насосы – пронизывают мембрану насквозь (интегральные белки), перемещают в клетке ионы и молекулы против градиента концентрации;
2) белки-каналы - пронизывают мембрану насквозь (интегральные белки), они образуют места избирательного прохождения для ряда ионов (натрия, калия, кальция, хлора) по градиенту концентрации;
3) рецепторные белки прикреплены на внешней поверхности мембраны и состоят не только из молекул белка, но и из полисахаридов, осуществляют распознавание нужных молекул (например: медиаторов, гормонов и др.) и их фиксацию на мембране;
4) ферменты облегчают протекание химической реакции на внутренней и внешней поверхности нейрона (полуинтегральные белки).
В некоторых случаях один и тот же белок может выполнять функции как рецептора, так и насоса.
Рис. Схема строения мембраны нейрона:
1. молекула липида;
2. интегральные белки;
3. липидный бислой;
4. полуинтегральные белки;
5. периферические белки;
7. субмембранный слой;
8. актиновые микрофиламенты;
10. промежуточные филаменты;
11. углеводные части молекул гликопротеинов и гликолипидов
Основные функции мембраны следующие:
1. Мембрана – граница между внутренней и внешней средой клетки, которые существенно отличаются по своим физико-химическим показателям.
2. Участие в процессах обмена веществ, благодаря наличию ферментов. Большинство биохимических процессов в клетке происходит на мембранах.
3. Регуляция физиологических процессов в клетке. На мембране расположены рецепторные белки, воспринимающие сигналы, поступающие извне и преобразующие их в команды, регулирующие внутриклеточные процессы.
Как же работает нейрон? В любой живой клетке существует разность потенциалов на мембране. Разница разрядов между наружной и внутренней поверхностью покоящейся мембраны называется потенциалом покоя (ПП). ПП нейрона всегда отрицателен, это значит, что внутренняя среда клетки заряжена более отрицательно, чем внеклеточная среда. То есть внутри клетки существует избыточный отрицательный заряд.
Потенциал покоя возникает в результате двух причин:
1) неодинакового распределения ионов по обе стороны мембраны. Внутри клетки находится больше всего ионов калия, снаружи его мало. Ионов натрия и ионов хлора больше снаружи, чем внутри. Такое распределение ионов называется ионной асимметрией;
2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана хорошо проницаема для ионов калия, малопроницаема для ионов натрия и хлора и совсем непроницаема для органических веществ.
За счет чего формируется разность содержания ионов по обе стороны мембраны? Дело в том, что в любой живой клетке в мембране существует постоянно работающий и потребляющий энергию Na+/K+ ионный насос, который откачивает из клетки ионы натрия и закачивает внутрь ионы калия. Существует асимметричность обмена Na+ на K+ в соотношении 3:2. Работа насоса приводит к тому, что внутри клетки концентрация ионов калия примерно в 30 раз превышает их концентрацию вне клетки, тогда как концентрация ионов натрия в клетке примерно в 50 раз меньше, чем снаружи. В результате работыNa+/K+ насоса на мембране появляется слабый электрический потенциала (-10 мВ).
Далее натрий так и остается снаружи (так как мембрана для него малопроницаема), а калий из-за хорошей мембранной проницаемости начинает потихоньку выходить из клетки во внеклеточное пространство. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузии по градиенту. Выходя, калий выносит свой положительный заряд, что приводит к увеличению положительного заряда наружной стороны мембраны (до -70 мВ). Оставшиеся внутри отрицательно заряженные ионы хлора и органические соединения прижимаются к внутренней стороне мембраны (мембрана для них непроницаема). В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны не может быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. В нейроне внутренняя поверхность клетки заряжена отрицательно на величину около 70 мВ относительно внешней.
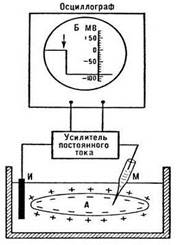
Рис. Регистрация потенциала покоя
Теперь рассмотрим вопрос: Как возбуждается нейрон? Под возбуждением нейрона понимают генерацию нейроном потенциала действия (ПД).
Потенциалом действияназывают сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны. Дело в том, что при действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов натрия она повышается в 400–500 раз. Напомним, что благодаря постоянной работе насосных каналов концентрация натриевых ионов вне клетки примерно в 50 раз больше, чем в клетке, поэтому при открытии натриевых каналов ионы натрия устремляются в клетку, а ионы калия через открытые калиевые каналы начинают выходить из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны приобретает отрицательный заряд, внутренняя – положительный.

Рис. Потенциал действия: a - локальный ответ; b,c – спайк (b - фаза деполяризации, с - фаза реполяризации); d,e - следовые колебания ( d - отрицательный следовой потенциал; e - положительный следовой потенциал).
Выделяют следующие компоненты потенциала действия:
1) локальный ответ;
2) высоковольтный пиковый потенциал (спайк);
3) следовые колебания:
а) отрицательный следовой потенциал;
б) положительный следовой потенциал.
Локальный ответ связан с небольшим током натрия внутрь клетки. Пока раздражитель не достиг 50–75 % от величины порога, проницаемость клеточной мембраны остается неизменой, и электрический сдвиг мембранного потенциала объясняется раздражающим агентом. Достигнув уровня 50–75 %, открываются потенциалзависимые ворота натриевых каналов, и возникает локальный ответ. Ионы натрия путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это та разность потенциалов, которая должна быть достигнута, чтобы возник лавинообразный ход ионов натрия в клетку.
Высоковольтный пиковый потенциал (спайк).
Пик потенциала действия состоит из двух фаз:
1) восходящей части – фазы деполяризации;
2) нисходящей части – фазы реполяризации.
Лавинообразное поступление ионов натрия в клетку приводит к падению потенциала на клеточной мембране (деполяризации). Чем больше ионов натрия войдет в клетку, тем в большей степени деполяризуется мембрана, тем больше откроется потенциалзависимых натриевых ворот. Постепенно заряд с мембраны снимается, а потом возникает с противоположным знаком. Возникновение заряда с противоположным знаком называется инверсией потенциала мембраны или овершутом. Движение ионов натрия внутрь клетки продолжается до момента электрохимического равновесия по иону натрия. Амплитуда потенциала действия не зависит от силы раздражителя, она зависит от концентрации ионов натрия и от степени проницаемости мембраны к ионам натрия.
Нисходящая фаза (фаза реполяризации) возвращает заряд мембраны к исходному знаку. Онахарактеризуется:
1. снижением проницаемости клеточной мембраны для натрия (инактивация натриевых ворот);
2. возрастает проницаемость мембраны для калия, в результате повышается выход калия из клетки с увеличением положительного заряда на мембране;
3. увеличение активности Nа + -К + насоса, который восстанавливает заряд клеточной мембраны
Однако сразу полного восстановления мембранного потенциала не происходит.
В процессе восстановительных реакций на клеточной мембране регистрируются следовые потенциалы – положительный и отрицательный. Следовые потенциалы являются непостоянными компонентами потенциала действия. Отрицательный следовой потенциал результат проникновения внутрь клетки какого-то количества натрия. Таким образом, отрицательный следовой потенциал - это следовая деполяризация.
Положительный следовой потенциал возникает при гиперполяризации клеточной мембраны в процессе восстановления клеточного заряда за счет выхода ионов калия и работы натрий-калиевого насоса. Как только калиевая проницаемость возвращается к исходному уровню - регистрируется мембранный ПП.
Далее это возбуждение (ПД) должно передаться другому нейрону или какой-то другой клетке, например мышечной, железистой и др. Как же осуществляется проведение возбуждения?
Возбуждение в виде потенциала действия покидает тело нейрона по его отростку, который называется аксоном. Аксоны отдельных нейронов обычно объединяются в пучки - нервы, а сами аксоны в этих пучках называются нервными волокнами. Природа позаботилась, чтобы волокна максимально хорошо справлялись с функцией проведения возбуждения в виде потенциалов действия. Для этой цели отдельные нервные волокна (аксоны отдельных нейронов) имеют специальные чехлы – миелиновые оболочки, которые образуется из вещества миелина, примерно на 2 /3 состоящего из жира и являющегося хорошим электрическим изолятором. Через каждые 0,5-1,5 мм оболочка прерывается, формируются перехваты Ранвье. Каждый участок образован только одной клеткой, поэтому перехваты - это стыки между двумя клетками, образующими соседние участки миелиновой оболочки. В периферических нервах миелин образуется клетками, которые получили название шванновских, а в головном мозге это происходит за счет клеток олигодендроглии.
Известно, что у новорожденного ребенка миелинизировано примерно 2 /3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция внимания, он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций. В то же время известны заболевания нервной системы человека, которые связаны с демиелинизацией нервных волокон, что сопровождается тяжелыми страданиями. К самым известным относится рассеянный склероз. Это заболевание развивается незаметно и очень медленно, последствием является паралич движения.
Почему же так важна миелинизация нервных волокон? Оказывается, миелинизированные волокна в сотни раз быстрее проводят возбуждение, чем немиелинизированные, т. е. нейронные сети нашего мозга могут работать с большей скоростью, а значит, более эффективно. Поэтому не миелинизируются в нашем организме только самые тонкие волокна (менее 1 мкм в диаметре), которые проводят возбуждение к медленно работающим органам - кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре.
Как происходит распространение возбуждения по нервному волокну? Вначале разберем случай немиелинизированного нервного волокна.
Рис. Схема распространения потенциала действия по немиелинизированному волокну.
Практически все нервные волокна в центральной нервной системе человека имеют миелиновые чехлы. На рисунке показано, что если вдоль волокна, покрытого миелином, регистрировать прохождение возбуждения, то потенциал действия возникает только в перехватах Ранвье. Оказывается, миелин, являясь хорошим электрическим изолятором, не пропускает выхода линий тока от предшествующего возбужденного участка. Выход тока в этом случае возможен только через те участки мембраны, которые не покрыта изолятором. Благодаря такому устройству мембрана волокна возбуждается только в местах перехватов Ранвье. Вследствие этого потенциал действия (возбуждение) как бы перескакивает через участки изолированной мембраны. Другими словами, возбуждение движется скачками от перехвата к перехвату – сальтаторно. В узловых перехватах количество натриевых каналов достигает 12 000 на 1 мкм2, что значительно больше, чем в любом другом участке волокна. В результате узловые перехваты являются наиболее возбудимыми. Сальтаторный способ распространения возбуждения экономичен, скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с). Время проведения возбуждения по миелиновому волокну обратно пропорционально длине между перехватами.
Рис. Распространение потенциала действия по миелинизированным волокнам
Законы проведения раздражения по нервному волокну.
1.Закон анатомо-физиологической целостности.Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
2. Закон изолированного проведения возбуждения. В нервах по нервным волокнам возбуждение передается изолированно.
3. Закон двустороннего проведения возбуждения. Нервное волокно при раздражении проводит нервные импульсы в двух направлениях – центростремительно и центробежно. В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
4. Распространение возбуждения в обе стороны идет с одинаковой скоростью.
5. В миелинизированных волокнах возбуждение проходит, не затухая, без декремента.

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.
Читайте также:
