
При действии анестетика на нерв нарушается закон
Нервное волокно обладает следующими физиологическими свойствами: возбудимостью, проводимостью, лабильностью.
Проведение возбуждения по нервным волокнам осуществляется по определенным законам.
Закон двустороннего проведения возбуждения по нервному волокну. Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от возбужденного участка (места его возникновения), т. е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. Естественным направлением распространения возбуждения является: в афферентных проводниках - от рецептора к клетке, в эфферентных - от клетки к рабочему органу.
Закон анатомической и физиологической целостности нервного волокна. Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву (законы анатомической и физиологической целостности). Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е., к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается.
Закон изолированного проведения возбуждения по нервному волокну. В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервноговолокна на другое, то нормальное функционирование органов было бы невозможно.
Возбуждение (потенциал действия) распространяется по нервному волокну без затухания.
Периферический нерв практически неутомляем.
Механизм проведения возбуждения по нерву.
Особенности распространения возбуждения по безмиелиновым волокнам:
1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением.
2. Возбуждение распространяется с небольшой скоростью.
3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна).
По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров.
Однако низкая скорость распространения возбуждения и его затухание не всегда выгодно организму. Поэтому природой был выробатан еще один дополнительный механизм.
Механизмы проведения возбуждения в миелиновых волокнах. При действии раздражителя пороговой силы на мембрану миелинового волокна в области перехвата Ронвье изменяется проницаемость для ионов Nа + , которые мощным потоком устремляются внутрь волокна. В этом месте также изменяется заряд мембраны, что тоже ведет к возникновению круговых токов. Этот ток идет через межтканевую жидкость к соседнему перехвату, где происходит смена заряда. Таким образом возбуждение перепрыгивает с одного участка на другой. Обратное движение возбуждения невозможно так как участок через который оно прошло находится в фазе абсолютной рефрактерности.
Особенности распространения возбуждения по миелиновым волокнам:
1. Возбуждение распространяется скачкообразно (сальтаторно) и все волокно сразу не охватывается возбуждением.
2. Возбуждение распространяется с большой скоростью.
3. Возбуждение распространяется с без декримента.
По змиелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам т.е. там где требуется высокая скорость ответной реакции.

Анестетики разделяют на:
- Местные
- Общие:
а) ингаляционные - летучие жидкости и газы
б) неингаляционные (внутривенные)
1. Местные анестетики
Местные анестетики обратимо снижают возбудимость чувствительных нервных окончаний и блокируют проведение афферентных импульсов в нервных стволах в зоне непосредственного применения, используются для устранения боли.
Местные анестетики классифицируют на сложные эфиры (анестезин, дикаин, новокаин) и замещенные амиды (лидокаин, тримекаин, бупивакаин). Местные анестетики — сложные эфиры подвергаются гидролизу псевдохолинэстеразой крови и действуют в течение 30 — 60 мин. Их эффект пролонгируют антихолинэстеразные средства (прозерин). Продукт гидролиза — n-амино-бензойная кислота ослабляет бактериостатическое влияние сульфаниламидов. Замещенные амиды кислот инактивируются монооксигеназной системой печени в течение 2 — 3 ч. Бупивакин вызывает местную анестезию продолжительностью 3 — 6 ч, после ее прекращения длительно сохраняется анальгетический эффект.
С точки зрения практического применения анестетики подразделяют на следующие группы:
- Средства, применяемые для поверхностной (терминальной) анестезии: Кокаин, Дикаин, Анестезин, Пиромекаин
- Средства, применяемые преимущественно для инфильтрационной и проводниковой анестезии: Новокаин, Бупивакаин
- Средства, применяемые для всех видов анестезии: Лидокаин, Тримекаин
Механизм действия
Местные анестетики представляют собой третичные азотистые основания. Они состоят из гидрофильной и липофильной частей, соединенных эфирной или амидной связями. Механизм действия определяет липофильная часть, имеющая ароматическую структуру. Для нанесения на слизистые оболочки и кожу и парентерального введения применяют водные растворы хлористоводородных солей местных анестетиков. В слабощелочной среде тканей (рН=7,4) соли гидролизуются с освобождением оснований. Основания местных анестетиков растворяются в липидах мембран нервных окончаний и стволов, проникают к внутренней поверхности мембраны, где превращаются в ионизированную катионную форму.
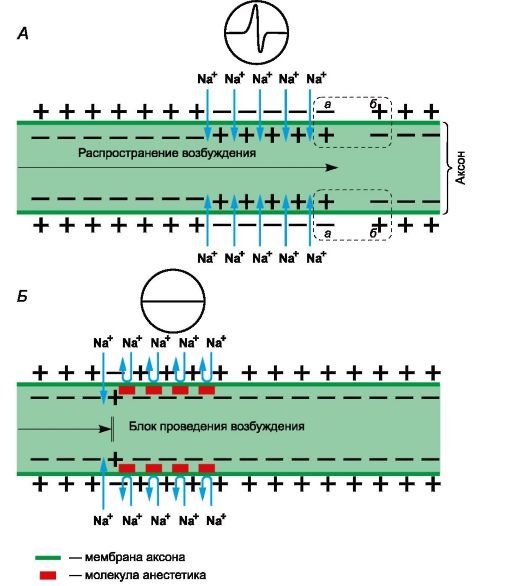
Рисунок 1 | Механизм действия местных анестетиков
Рецепторы для местных анестетиков локализованы в S6-сегменте IV домена внутриклеточной части натриевых каналов. Связываясь с рецепторами, катионы местных анестетиков пролонгируют инактивированное состояние натриевых каналов, что задерживает развитие следующего потенциала действия. Местные анестетики не взаимодействуют с закрытыми каналами в период потенциала покоя. Таким образом, в зоне нанесения местных анестетиков не развиваются потенциалы действия, что сопровождается блоком проведения нервных импульсов. Избирательное влияние местных анестетиков на чувствительные афферентные нервы обусловлено генерацией в них длительных (более 5 мс) потенциалов действия с высокой частотой.
В первую очередь местные анестетики блокируют безмиелиновые С и миелиновые Аδ и Аβ волокна (афферентные пути, проводящие болевые и температурные раздражения; вегетативные нервы). На волокна, окруженные миелиновой оболочкой, местные анестетики действуют в области перехватов Ранвье. Толстые миелиновые волокна (афферентные пути, проводящие тактильные раздражения; двигательные нервы) слабее реагируют на местные анестетики. Кроме того, устойчивость двигательных нервов к анестезии обусловлена низкочастотными короткими (менее 5 мс) потенциалами действия. В очаге воспаления в условиях ацидоза нарушаются диссоциация хлористоводородных солей местных анестетиков и образование их свободных липидорастворимых оснований, поэтому обезболивающее влияние утрачивается. Например, местная анестезия может оказаться неэффективной при удалении зуба в случае тяжелого периодонтита.
2. Общие анестетики
2.1. Ингаляционные анестетики
2.1.1 Летучие жидкости
Теории механизма действия общих анестетиков
Эффекты ингаляционных анестетиков не могут быть объяснены одним молекулярным механизмом. Скорее всего многокомпонентное действие каждого анестетика реализуется через множество мишеней. Тем не менее, эти эффекты сходятся на ограниченном числе изменений, лежащих в основе физиологических эффектов. На данный момент существуют липидная и белковая теории анестезии, но ни одна из них пока не описывает последовательность событий, происходящих от взаимодействия молекулы анестетика и ее мишеней до физиологических эффектов.
Рисунок 2 | Ингаляционные анестетики
Активность ингаляционных анестетиков оценивают по минимальной альвеолярной концентрации (МАК). Доза, создающая 1 МАК, предотвращает у половины пациентов движения в ответ на хирургическое вмешательство. Сила общих анестетиков коррелирует с их растворимостью в жирах, что говорит о важности взаимодействия с гидрофильными мишенями. В частности, обнаружение связи между силой анестетика и его липофильностью (правило Мейер-Овертона) дало начало липидной теории механизма действия анестетиков. Липидная теория анестезии утверждает, что анестетики растворяются в двойном липидном слое биологических мембран и вызывают анестезию, достигая критической концентрации в мембране. Наиболее усложненные версии липидной теории требуют, чтобы молекулы анестетиков вызывали пертурбацию (изменение свойств) мембраны.
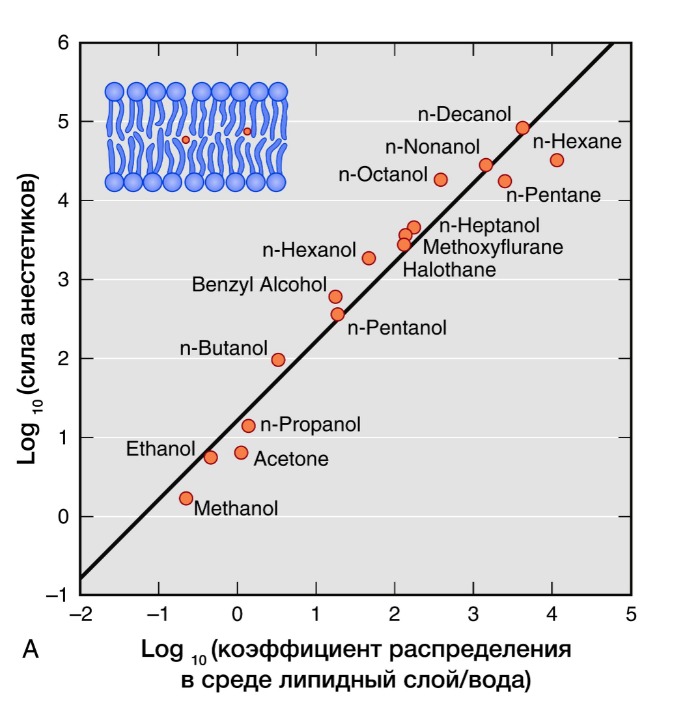
Рисунок 3 | Правило Мейер-Овертона
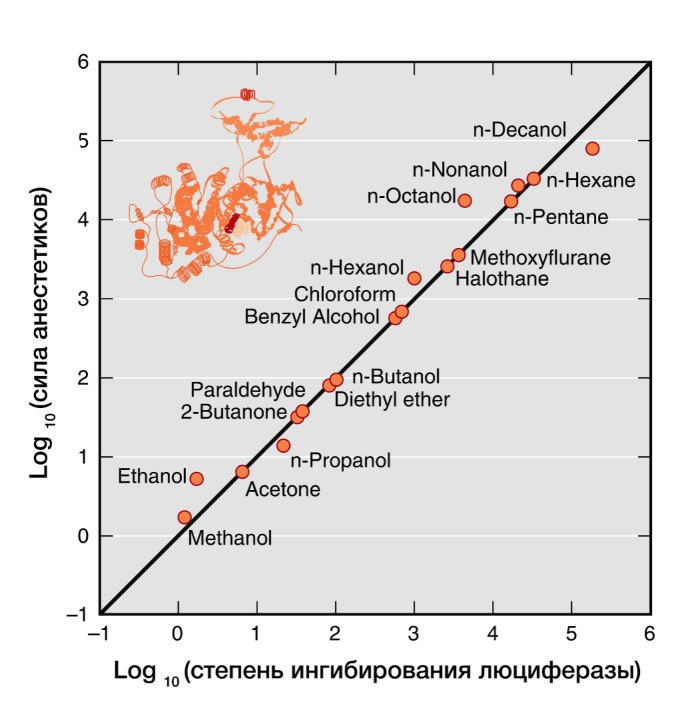
Рисунок 4 | В 20 веке было показано, что сила общих анестетиков коррелируют с их способностью ингибировать активность растворимого фермента люциферазы, который физиологически не является мишенью анестетиков, но служит в качестве безлипидной модели белковой молекулы для связывания анестетика.
Механизм действия
Таким образом, общие анестетики изменяют физико-химические свойства липидов мембран нейронов и нарушают взаимодействие липидов с белками ионных каналов. При этом уменьшается транспорт в нейроны ионов натрия, сохраняется выход менее гидратированных ионов калия, в 1,5 раза возрастает проницаемость хлорных каналов, управляемых ГАМК-А рецепторами. Итогом этих эффектов становится гиперполяризация с усилением процессов торможения. Общие анестетики подавляют вход в нейроны ионов кальция, блокируя Н-холинорецепторы и NMDA-рецепторы глутаминовой кислоты; снижают подвижность Са 2+ в мембране, поэтому препятствуют кальций-зависимому выделению возбуждающих нейромедиаторов. Наиболее чувствительны к действию общих анестетиков полисинаптические системы ЦНС — кора больших полушарий (10 13 – 10 14 синапсов), таламус, ретикулярная формация, спинной мозг. К наркозу устойчивы дыхательный и сосудодвигательный центры продолговатого мозга.
Летучие ингаляционные анестетики:
- постсинаптически усиливают тормозящую передачу путем потенциирования лиганд-управляемых ионных каналов, активируемых ГАМК и глицином;
- экстрасинаптиески путем усиления ГАМК-рецепторов и ионных токов утечки;
- пресинаптически за счет повышения базального высвобождения ГАМК.
Ингаляционные анестетики подавляют возбуждающую синаптическую передачу путем снижения высвобождения глутамата и постсинаптически путем ингибирования ионотропных глутаматных рецепторов. Парализующий эффект местных анестетиков включает действие на спинной мозг, в то время как седация/наркоз и амнезия включают супраспинальные механизмы памяти, сна и сознания.
2.1.2. Газовый наркоз
В анестезиологии широко применяют ингаляционный газовый анестетик азота закись (N2O). В конце 1980-х гг. в зарубежную анестезиологическую практику вошел инертный газ ксенон.
Азота закись представляет собой бесцветный газ характерного запаха, хранится в металлических баллонах под давлением 50 атм в жидком состоянии, не горит, но поддерживает горение. Ее смеси с анестетиками группы летучих жидкостей в определенных концентрациях взрывоопасны. В субнаркотических концентрациях (20 — 30 %) азота закись вызывает эйфорию (веселящий газ) и сильную анальгезию. В концентрации 20 % обеспечивает обезболивание в такой же степени, как 15 мг морфина. Закись азота на ГАМК-А рецепторы не влияет. Используется только в комбинациях, так как МАК составляет 104 %.
Инертный газ ксенон считают лучшей альтернативой азота закиси, так как он обладает более выраженным наркозным действием, индифферентностью и экологической безопасностью. Способность ксенона вызвать наркоз была открыта в связи с практикой глубоководных погружений и развитием гипербарической физиологии. Ксенон бесцветен, не горит и не обладает запахом, при соприкосновении со слизистой оболочкой рта создает на языке ощущение горьковатого металлического вкуса. Отличается низкой вязкостью и высокой растворимостью в липидах, выводится легкими в неизмененном виде. Разработана технология ксенонсберегающей анестезии с включением минимального потока и системы рециклинга для повторного многократного использования газа. Такая технология успешно решает важную в практическом отношении проблему дефицита и дороговизны ксенона. В механизме наркозного эффекта ксенона имеют значение блокада циторецепторов возбуждающих нейромедиаторов — Н-холинорецепторов, NMDA-рецепторов глутаминовой кислоты, а также активация рецепторов тормозящего нейромедиатора глицина. При взаимодействии с циторецепторами ксенон выступает как протонсвязывающий кластер и образует комплексы с катионами НСО + , NH2 + , HNCH + . Ксенон проявляет свойства антиоксиданта и иммуностимулятора, снижает выделение гидрокортизона и адреналина из надпочечников.
2.2. Неингаляционные (внутривенные) наркозные средства подразделяют на три группы:
Препараты короткого действия (3 — 5 мин)
- пропанидин (эпонтол, сомбревин)
- пропофол (диприван, рекофол)
Препараты средней продолжительности действия (20 — 30 мин)
- кетамин (калипсол, кеталар, кетанест)
- мидазолам (дормикум, флормидал)
- гексенал (гексобарбитал-натрий)
- тиопентал-натрий (пентотал)
Препараты длительного действия (0,5 — 2 ч)
- натрия оксибутират
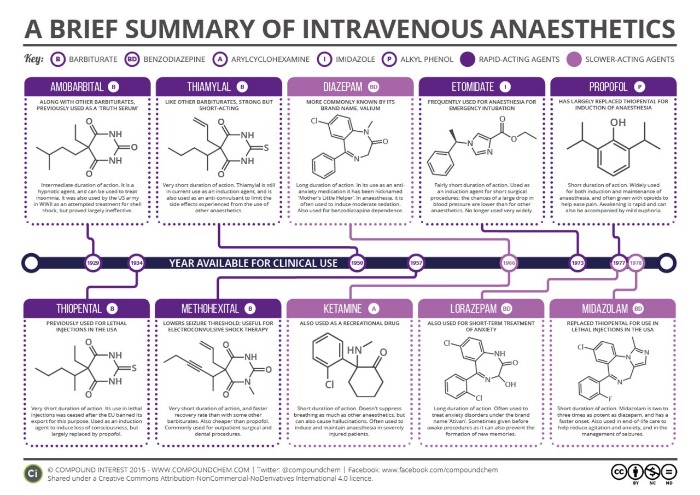
Рисунок 5 | Общие анестетики для внутривенного введения
Механизм действия
Наиболее широко используемым внутривенный анестетиком является пропофол. Его механизм действия связан с увеличение хлорной проводимости ГАМК рецепторов.
Метогекситал по скорости наступления и выхода из наркоза близок к пропофолу.
Барбитураты использовались для анестезии до введения в практику пропофола. Тиопентал вызывает быстрое наступление и быстрый выход из наркоза при разовом введении, но он быстро накапливается при повторном или пролонгированном введении и таким образом замедляет выход из анестезии. Барбитураты являются лигандами барбитуратных рецепторов. В малых дозах они аллостерически усиливают действие ГАМК на ГАМК А-рецепторы . При этом удлиняется открытое состояние хлорных каналов, возрастает вход в нейроны анионов хлора, развиваются гиперполяризация и торможение. В больших дозах барбитураты прямо повышают хлорную проницаемость мембран нейронов. Кроме того, они тормозят высвобождение возбуждающих медиаторов ЦНС — ацетилхолина и глутаминовой кислоты, блокируют АМРА-рецепторы (квисквалатные рецепторы ) глутаминовой кислоты. Барбитураты обладают церебропротективными свойствами и могут быть использованы с такой целью.
Бензодиазепины используются в основном как анксиолитики и для седации с сохранением сознания.Все бензодиазепиновые рецепторы аллостерически усиливают кооперацию ГАМК с ГАМК А-рецепторами, что сопровождается повышением хлорной проводимости нейронов, развитием гиперполяризации и торможения. Реакция с бензодиазепиновыми рецепторами происходит только в присутствии ГАМК. Ремимазолам - самый молодой бензодиазепин, имеет очень короткое время действия из-за быстрой нейтрализации эстеразами плазмы.
Кетамин химически является производным фенциклидина. Синаптические механизмы действия кетамина многообразны. Он является неконкурентным антагонистом возбуждающих медиаторов головного мозга глутаминовой и аспарагиновой кислот в отношении NMDA-рецепторов. Эти рецепторы активируют натриевые, калиевые и кальциевые каналы мембран нейронов. При блокаде рецепторов нарушается деполяризация. Кроме того, кетамин стимулирует освобождение энкефалинов и β-эндорфина; тормозит нейрональный захват серотонина и норадреналина. Последний эффект проявляется тахикардией, ростом АД и внутричерепного давления. Кетамин расширяет бронхи. При выходе из кетаминового наркоза возможны бред, галлюцинации, двигательное возбуждение (эти нежелательные явления предупреждают введением дроперидола или транквилизаторов).Важным терапевтическим эффектом кетамина является нейропротективный. Как известно, в первые минуты гипоксии мозга происходит выброс возбуждающих медиаторов — глутаминовой и аспарагиновой кислот. Последующая активация NMDA-рецепторов, увеличивая во внутриклеточной среде концентрацию ионов натрия и кальция и осмотическое давление, вызывает набухание и гибель нейронов. Кетамин как антагонист NMDA-рецепторов устраняет перегрузку нейронов ионами и связанный с этим неврологический дефицит.
Самым новым внутривенным анестетиком является дексмедетомидин. Это высокоселективный агонист а2-адренорецепторов с седативными, симпатолитическими, снотворными и анальгетическими эффектами. Его основное действие - в качестве агониста на а2 рецепторы в голубом пятне.
Законы проведения возбуждения в нервах
При изучении проведения возбуждения в нервных волокнах уста лено несколько правил, или законов , протекания этого процесса.
Закон физиологической непрерывности нерва . Обязательным условием проведения возбуждения по нервному волокну является анатомия и функциональная целость возбудимой мембраны осевого цилиндра.
Поэтому не только перерезка нерва, но и любое воздействие, нарушающее целость мембраны осевого цилиндра, например перевязка нерва, чрезмерное натяжение нервных волокон, создают непроводимость. Непроводимость наступает также при воздействиях, нарушающих генерацию нервного импульса, чрезмерное охлаждение или согревание, прекращение кровоснабжения, различные химические агенты, в частности местные обезболивающие — новокаин, кокаин, дикаин, прекращают проведение по нерву.
На основании подобных фактов сформулирован закон, гласящий, что проведение возможно только лишь при условии физиологической непрерывностн нервных волокон.
Закон двустороннего проведения . При нанесении раздражения на нервное волокно возбуждение распространяется по нему двусторонне, т.е. и в центробежном и в центростремительном направлениях. Это доказывают следующим опытом.
К нервному волокну, безразлично к двигательному или чувствительному, прикладывают две пары электродов, связанных с двумя электроизмерительными приборами А и Б ( рис. 154 ). Раздражение наносят между этими электродами. В результате двустороннего проведения возбуждения приборы зарегистрируют прохождение импульса как под электродами А, так и под электродами Б.
Рис. 154. Доказательство двустороннего проведения импульса в нерве (объяснение в тексте).
Доказательство двустороннего проведения возбуждения может быть получено и без помощи специальной электрофизиологической аппаратуры. Одна из бедренных мышц лягушки — m. gracilis — иннервирована таким образом, что к двум ее половинам подходят разветвления аксонов одних и тех же двигательных нервных волокон. На этой мышце В. Кюне проделал следующий опыт: он разрезал мышцу на две части, соединенные только посредством разветвлений нервных волокон, а затем одну часть мышцы он раздражал электрическим током. Это приводило к сокращению обеих половин мышцы, так как возбуждение распространялось по веточкам одного и того же аксона сначала центростремительно, а затем центробежно.
Закон двустороннего проведения нерва не противоречит тому факту, что в рефлекторных дугах центральной нервной системы и в нервных окончаниях на периферии возбуждение распространяется односторонне. Эта односторонность есть свойство не нервного волокна, а синапса, т. е. места перехода возбуждения с одного нейрона на другой или с нейрона на иннервированную им ткань.
Закон изолированного проведения по нерву . Всякий периферический нервный ствол состоит из большого числа нервных волокон — двигательных, чувствительных и вегетативных. Волокна нервных стволов снабжают большое число иногда далеко отстоящих друг от друга периферических образований. Так, блуждающий нерв иннервирует все органы грудной и значительную часть органов брюшной полости, седалищный нерв иннервирует всю мускулатуру, костный аппарат, сосуды и кожу нижней конечности.
Понятно, что нормальное функционирование всех периферических органов и тканей возможно лишь в том случае, если импульсы распространяются по каждому волокну нерва изолированно, не переходя с одного волокна на другое и оказывая действие только на те клетки, на которых нервные волокна оканчиваются.
Изолированное проведение в отдельных волокнах смешанного нерва может быть доказано опытом на скелетной мышце, иннервированной смешанным нервом, в образовании которого участвует несколько спинномозговых корешков. Если раздражать один из этих корешков, то сокращается не вся мышца, как это было бы, если бы возбуждение переходило с одних нервных волокон на другие, а только те группы мышечных волокон, которые иннервированы раздражаемым корешком. Еще более строгое доказательство изолированного проведения возбуждения может быть получено ври отведении потенциалов действия от различных нервных волокон нервного ствола.
Проведение возбуждения в нерве
195. Один конец нерва раздражают электрическим током. На другом его конце размещены две пары электродов. При помощи одной из них можно регистрировать на осциллографе ПД, при помощи другой – измерять посредством микроамперметра силу тока, проходящего через участок нерва. Затем нерв раздавливают в средней части. Что покажут приборы, если теперь снова нанести раздражение?
196. В результате патологического процесса поражен участок нерва, содержащий несколько перехватов Ранвье. Проведение возбуждения прекратилось. Как можно восстановить его?
197. Седалищный нерв лягушки содержит аксоны мотонейронов, аксоны чувствительных клеток и аксоны нейронов симпатических ганглиев. Представим, что у нас имеется препарат, избирательно блокирующий проведение возбуждения в двигательных и чувствительных волокнах. Как доказать в эксперименте, что препарат подействовал и теперь в нерве работают только симпатические волокна?
Решение. Понятно, что следует применить правило САС. Нужно сравнить особенности каких-то элементов в каждой из трех рассматриваемых систем. Проще всего использовать порог раздражения и скорость проведения возбуждения. Порог раздражения у симпатических волокон в сто раз выше, чем, например, для волокон А – альфа, а скорость проведения в 10 – 100 раз меньше, чем в двигательных и чувствительных волокнах.
Следовательно, для решения задачи достаточно сравнить величины одного из этих показателей (или их обоих) до и после воздействия препарата.
198. Скорость проведения возбуждения в мякотных волокнах пропорциональна диаметру волокна, а в безмякотных – корню квадратному из диаметра. Чем объясняется наличие такой зависимости и ее различие для указанных двух групп волокон, т. е., меньшая выраженность зависимости для безмякотных волокон?
Решение. Задача состоит из двух частей. Рассмотрим их последовательно. Начнем с мякотных волокон. Сначала применим правило АСФ. Требуется ответить всего на один вопрос – как распространяется возбуждение в мякотных волокнах? Сальтаторно, скачками от одного перехвата Ранвье к другому. Теперь на минуту нужно вспомнить правило АРР-ВС. Какие отличия между тонкими и толстыми волокнами связаны с перехватами Ранвье? Если Вы этого не помните – не страшно. Важно, что мы установили, какую информацию следует получить. Оказывается, что в толстых волокнах расстояние между перехватами Ранвье больше. Поэтому скачок нервного импульса длиннее и возбуждение распространяется с большей скоростью.
Внимание!Объем очередной нашей задачи по сравнению с предыдущими увеличен. Это означает, что Вам придется иметь дело с большим количеством элементов, требующих сопоставления. Последовательность мышления в данной задаче особенно необходима.
199.Выполнен эксперимент на двух нервах. На каждом из них установлены раздражающие электроды и две пары отводящих электродов. Первая пара размещена рядом с раздражающими электродами, вторая на некотором расстоянии от них. Каждый нерв однократно раздражают ударом тока и регистрируют ПД.
Получены следующие кривые (рис. 7.9). Затем каждый из нервов начинают раздражать ударами тока нарастающей силы. Как при этом будет изменяться амплитуда ПД у первого и второго нерва?
Решение. Хотя ответ на первую часть задачи Вам может быть уже известен из учебника, попробуем найти его по правилам. Наверно, Вы довольно быстро определили, что задача на обратное правило АРР-ВС. Действительно, на двух нервах получены разные результаты, а почему –
Первая часть задачи решена. Перейдем ко второй. Теперь различия между нервами установлены и нужно определить ожидаемые результаты. Следовательно, применим прямое правило АРР-ВС. Если в первом нерве все волокна одинаковы (это допущение в известной степени условно, но оно необходимо для логики наших рассуждений), то и возбудимость их одинакова или по крайней мере близка. Поэтому при увеличении силы раздражения амплитуда суммарного ПД будет изменяться мало или вообще не изменится. Во втором же нерве волокна разные, следовательно, и возбудимость их разная. В таком случае на более слабые раздражения ответят только самые возбудимые волокна. При увеличении силы раздражения начнут возбуждаться менее возбудимые и т. д. Суммарная амплитуда ПД будет возрастать до тех пор, пока не возбудятся все волокна.
200. Если рассматривать организм в эволюционном ряду, то выясняется, что развитие рефлекторной деятельности совпадает с миелинизацией нервных волокон. Чем можно объяснить это совпадение?
Решение. Вспомним принцип целесообразности. Рефлекторные реакции обеспечивают приспособление организмов к воздействию факторов внешней и внутренней среды. Эффективность приспособления зависит, в частности, от того, насколько быстро протекают соответствующие реакции. А миелинизация нервных волокон как раз и обеспечивает более быстрое проведение возбуждения.
201. Как изменится количество молекул медиатора в окончаниях аксона, если в аксон ввести вещество, угнетающее метаболические процессы?
Решение. Правило АСФ. Медиатор синтезируется в теле нервной клетки и по аксону транспортируется в окончания. Этот процесс (аксонный транспорт) идет с затратой энергии. При ее нехватке транспорт нарушается. Значит, количество молекул медиатора в нервных окончаниях уменьшится.
202. Почему возбуждение, переходя в участок, соседний с возбужденным, не возвращается в уже пройденную точку? (рис. 7.11). В участке 1 возник ПД. Это приводит к появлению местного тока, который, проходя через мембрану в участке 2, деполяризует ее и в этом участке возникает новый ПД. Затем тот же процесс повторяется по отношению к участку 3. Но почему при этом не возникает ПД и в участке 1, тоже соседствующим с участком 2, но с другой стороны?
Решение. Применим обратное правило АРР-ВС. Узлы пересечения очевидны – ПД в участке 2 воздействует на участки 1 и 3. Результаты воздействия различны. Значит, нужно найти различия между участками 1и 3.
В участке 3 возбудимость нормальная, а вот в участке 1 еще не закончился АРП, который и не позволяет возникнуть возбуждению. Когда же возбудимость в участке 1 восстановится, то возбуждение уже перейдет в участок 3 и теперь по той же причине не сможет возбудиться участок 2 и т.д.
Решение. Применим правило АСФ. Возбуждение в двух соседних участках погашается, потому что в каждом из них АРП. Убрать состояние невозбудимости невозможно – это коренное свойство возбудимых тканей. В таком случае остается убрать один из возбужденных участков, чтобы он не мог погашать встречное возбуждение. Для этого нужно сделать так, чтобы после нанесения раздражения возбуждение могло двигаться только в одну сторону. С этой целью участок нервного кольца по одну сторону от раздражающих электродов на очень короткое время блокировали. Возбуждение после раздражения могло поэтому двигаться только в одном направлении. Но за время пробега волны возбуждения по всему кольцу возбудимость блокированного участка восстановилась и теперь возбуждение беспрепятственно двигалось по кругу, как уже было сказано, в течение многих часов.
Опыт, действительно, очень красив. Помимо прочего он лишний раз показывает, что в организме можно получать самые необычные реакции, если искусственным путем вмешиваться в работу механизмов, сложившихся за миллионы лет эволюции.
204. В каком нерве при возбуждении выделяется больше тепла – мякотном или безмякотном? Почему?
Решение. Применим правило АСФ. Почему в живых тканях выделяется тепло при их функционировании? Это связано с протеканием экзотермических реакций. При возбуждении нерва они обеспечивают выработку энергии, которая расходуется на работу натриево-калиевого насоса. Чем больше ионов натрия и калия проходит через мембрану, тем интенсивней должен работать насос. В мякотном нерве ионы проходят только в перехватах Ранвье, в безмякотном – на всем его протяжении. (Вот главная информация, без которой нельзя решить задачу). Поэтому в безмякотном нерве расходуется больше ионов. Следовательно, при его возбуждении затрачивается больше энергии и выделяется больше тепла.
205. В свое время русский физиолог Н. Е. Введенский в эксперименте на НМП доказал, что нерв даже при длительном раздражении лишь в очень малой степени подвержен утомлению.
Трудность заключалась в том, что в то время не существовало приборов для регистрации ПД, а мышца, которую использовали в качестве индикатора утомления нерва, утомляется гораздо быстрее, чем нерв. Как бы Вы поставили опыт?
Примечание.При выключении сильного постоянного тока возбудимость нерва восстанавливается не сразу. Это искажает получаемый результат. Поэтому Введенский применил метод минимальной поляризации. Блок вызывали сильным током, а поддерживать возникшую после этого непроводимость нерва можно было уже относительно слабым током. При его выключении проводимость восстанавливалась сразу же. В этих условиях нерв не обнаруживал утомления даже после многочасового раздражения.
Задачи для самоконтроля
206. При удалении зуба для обезболивания используют раствор анестетика. Почему его вводят не в десну возле удаляемого зуба, а в область прохождения чувствительного нерва?
207. Для проверки закона изолированного проведения возбуждения можно использовать седалищный нерв лягушки, который образуется тремя раздельно выходящими из спинного мозга корешками. Раздражение отдельного корешка вызывает сокращение различных мышц. Однако, если корешки расположить близко друг от друга, то теперь раздражение надпороговым током каждый раз вызывает сокращение всей лапки. В чем причина?
209. При перерезке двигательного нерва мышца, которую он иннервировал, атрофируется. Чем можно объяснить это?
210. Нежная мышца лягушки иннервируется двумя веточками одного и того же нервного волокна. Как нужно поставить на этом объекте опыт, чтобы доказать закон двустороннего проведения возбуждения?
211. Это более сложная задача. Прежде чем приступить к решению, загляните в задачу 199. На смешанный нерв нанесли два раздражения разной силы. При этом установили, что в обоих случаях величины возникающего ПД были одинаковы, а составной ПД, который регистрировали на некотором расстоянии от места раздражения, носил обычный характер. Затем нерв обработали некоторым веществом и после этого опыт повторили. Обнаружилось, что применяемые раздражители теперь вызывали ПД не одной и той же величины, а разной. Изменился ли при этом характер кривой составного ПД?
212. Известно, что прохождение тока сопровождается падением напряжения по длине проводника. Многие аксоны имеют большую длину и обладают весьма большим сопротивлением. Однако амплитуды ПД в начале и в конце аксона одинаковы. Чем это объясняется?
213. Вы знакомы с законом двустороннего проведения возбуждения в нервном волокне. Однако в реальных условиях возбуждение движется по нервным волокнам в одном направлении – или центробежно, или центростремительно. Нет ли здесь противоречия?
214. Миелинизированные аксоны лягушки проводят импульсы возбуждения со скоростью 30 м/с, а аксоны кошки такого же диаметра – в три раза быстрей. Почему?
215. Почему при сальтаторном проведении возбуждения в миелинизированных волокнах ПД может возбудить не только соседний перехват Ранвье, но и один-два следующих?
Решения задач для самоконтроля
206. Правило АСС. При введении анестетика в участок, где проходит ствол чувствительного нерва, блокируется проведение болевых импульсов из всех областей, иннервируемых этим нервом. Если же сделать такую инъекцию в десну возле удаляемого зуба, то анестезия возникнет только в этом ограниченном участке. В первом случае блокада более надежна.
209. Иннервация мышцы обеспечивает не только ее сократительную деятельность, но и трофические влияния. В частности, нервные импульсы стимулируют активность ряда ферментов, например, АТФазы, синтез белков и т. д. При денервации нарушается синтез белков в мышце, уменьшается ее масса – происходит атрофия.
доказать. Для чистоты опыта дополнительно перережем общий ствол волокна, чтобы исключить возможное влияние нервной клетки.
211. Это более сложная задача, поэтому нужно, не торопясь, строго последовательно проанализировать условие. Предварительно загляните еще раз в задачу № 199. Правило АСФ. Сначала раздражитель разной силы вызывал ПД одинаковой амплитуды. Значит, в каждом случае возбуждались все волокна данного нерва. Составной характер ПД говорит о том, что в нерве имеются волокна, обладающие разной скоростью проведения возбуждения. После воздействия препарата раздражители разной силы стали вызывать ПД неодинаковой величины. Из этого следует, что более слабый раздражитель возбуждал меньшее количество волокон, а более сильный – большее. В таком случае введенный препарат снизил возбудимость определенной группы волокон и теперь для них более слабый раздражитель стал подпороговым., Если же данная группа волокон перестала возбуждаться, то на кривой составного ПД выпадает соответствующий ей зубец.
212. Правило АРР-ВС. В обычном проводнике разность потенциалов прилагается к концам проводника. В нерве же разность потенциалов возникает в ходе проведения возбуждения не между его началом и концом, а между двумя соседними участками, между которыми и проходит местный ток. Значит, элемент, определяющий различия между узлами пересечения, – это расстояние между точками, к которым прилагается разность потенциалов. В нерве это расстояние очень мало и поэтому падения напряжения на столь малом участке практически не происходит. Возникший же местный ток вызывает деполяризацию соседнего участка мембраны, после чего картина повторяется. Таким образом возбуждение и сопутствующий ему местный ток каждый раз возникают заново. Это и обеспечивает бездекрементное, без снижения амплитуды ПД проведение возбуждения. Аналогия в технике – ретрансляция слабых сигналов.
215. В данной задаче, как и в ряде других, можно использовать и прямое, и обратное правило АРР-ВС. Прямое правило. Идем от различий к результату. Амплитуда ПД в 4-5 раз превышает величину, необходимую для возбуждения соседнего перехвата (фактор надежности). Поэтому действие местного тока может распространиться и на соседние перехваты. Обратное правило.
Идем от различных результатов к элементам, их обусловившим. Если действие местного тока может распространиться на несколько перехватов, а не только на ближайший, значит, амплитуда ПД превышает уровень, позволяющий возбудить только соседний перехват.
| | | следующая лекция ==> | |
| Законы раздражения Тренировочные задачи | | | Мионевральная передача Тренировочные задачи |
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Читайте также:
