
При действии на нервную клетку
В физиологии применяются различные раздражители: электрические, химические, механические, температурные и др. Недостатки механического и температурного раздражителей заключается в том, что они трудно дозируются, и главное оказывают повреждающее действие на ткань. Химическое раздражение тоже трудно дозируется. Поэтому действие его сохраняется, несмотря на промывание препарата (наблюдается длительное сокращение мышц). Наиболее удобно электрическое раздражение. Его преимущество состоит в том, что сила и длительность раздражения легко и точно дозируются, количественный учет интенсивности раздражителя прост, а , главное, повторное применение электрического раздражения не оказывает действия.
При изучении действия эл тока на возбудимое образование можно использовать стимулятор любой системы. В качестве источника постоянного тока можно использовать гальванический пинцет, ножки которого состоят из разных металлов: одна из цинка, другая из меди. При прикосновении ножек пинцета к нерву образуется замкнутая цепь двух металлов и нерва, играющего роль проводника второго рода. Возникший ток и служит источником раздражения мышцы. Обычно гальванический пинцет используется для проверки сохранности нервно-мышечного препарата.
Источником раздражения препарата может быть его высыхание. При высыхании нерва начинает сокращаться мышца, что создает препятствие для работы с препаратом. Поэтому препарат необходимо постоянно смачивать раствором Рингера.
Для работы необходимы: стимулятор, электроды, лягушка, набор инструментов для препарирования, раствор Рингера, кристаллы поваренной соли, спиртовка, сосуд с горячей водой.
Методика выполнения работы: приготовить нервно-мышечный препарат, в течение всего опыта смачивать нерв раствором Рингера, Раздражение наносить на нерв как можно дальше от мышцы. Показателем возбудимости и проводимости нерва служит сокращение мышцы.
№1. Действие электрического раздражения
a)Раздражение ритмическим током: включите симулятор в сеть. Трансформатор подключите в положение 1-10, электроды от трансформатора подведите к объекту. Препарат разместите на электродах, воспользовавшись установкой. Рис. 7.А
Поставьте нужные параметры раздражения: частота 20имп в сек, длительность 1 мс, амплитуда 10-15. Пошлите ток к объекту и наблюдайте ответную реакцию – сокращение мышцы, которое прекращается сразу после выключения стимула.
Вывод:мышца сокращается множественно в соответствии с ритмом, амплитудой и длительностью наносимых раздражений. При одиночных импульсах, которые идут с определенным интервалом, мышца отвечает одиночным сокращением. При предъявлении импульсов с большой частотой мышца титанически сокращается, она в состоянии напряжения и не успевает расслабиться.
b) Раздражение постоянным током. Прикоснитесь гальв. Пинцетом к нерву препарата и наблюдайте сокращение мышцы. Внимание на быстроту возникновения и прекращения реакции при действии эл раздражения.
Вывод:одно прикосновение пинцетом вызывает одиночное сокращение мышцы.
№2. Механическое раздражение.
На участок нерва как можно ближе к кусочку позвоночника нанесите мех раздражение ребром закрытых ножниц(короткий удар), ущипните пинцетом.
Вывод:при одиночном коротком ударе наблюдается одиночное сокаращение. Реакция наблюдается во время действия раздражителя.
№ 3. Тепловое раздражение
Нагрейте препаровальную иглу, прикоснитесь иглой нагретой ( не острием). Проверьте, сокращается ли мышца при таком прикосновении к нерву не подогретой иглой.
Вывод: также наблюдается одно или несколько сокращений
№4. Химическое раздражение.
Положите на нерв несколько кристалликов поваренной соли. Отметьте момент наступления мышечных сокращений ( через сколько после соли), обратите внимание на их характер(сравните с эл). Смойте соль раствором Рингера. Заметьте, сразу ли прекращаются сокращения мышцы после снятия раздражения.
Вывод:характеризуется длительным латентным периодом с момента нанесения раздражителя до появления ответной реакции. Множественные сокращения мышц. Мышцы включаются в сокращение постепенно, по мере проникновения раздражителя в ткань.
№5.Раздражение вследствие высыхания
Расположите нерв так, чтобы он свободно свисал с электродов. Смачивая мышцу раствором Рингера, оставляйте нерв сухим. Дождитесь появления сокращения мышцы. Смочите нерв раствором Рингера. После этого сокращения мышцы обычно прекращаются, так как снимается раздражающее действие высыхания нерва.
Вывод:несколько раз сокращается
№6.Влияние нарушения проводимости
Наблюдайте ответную реакцию нервно-мышечного препарата на электрическое раздражение, на пощипывание пинцетом ( механическое раздражение). Затем нарушьте проводимость нерва путем наложения лигатуры между электродами и мышцей. Отметьте отсутствие эффекта при действии тех же раздражителей на нерв выше места наложения лигатуры.
Сделайте вывод об условиях сохранения нервно-мышечного препарата и об особенностях действия различных раздражителей.
1. Строение и функции элементов нервной клетки……………. ……..… 4
2. Обмен веществ в нейроне……………………. ………………………… 6
4. Основные функции нервной клетки……………………………….……. 8
4.1. Воспринимающая функция нейрона …………………………………. 8
4.2. Интегративная функция нейрона …………………………………….. 11
4.3. Эффекторная функция нейрона ………………………………. ……. 13
Основным структурным элементом нервной системы являются нервные клетки, или нейроны. Через нейроны осуществляется передача информации от одного участка нервной системы к другому, обмен информацией между нервной системой и различными участками тела. В нейронах происходят сложнейшие процессы обработки информации. С их помощью формируются ответные реакции организма (рефлексы) на внешние и внутренние раздражения. Нейроны разделяются на три основных типа: афферентные, эфферентные, промежуточные нейроны.
Афферентные нейроны (чувствительные, или центростремительные) передают информацию от рецепторов в центральную нервную систему. Тела этих нейронов расположены вне центральной нервной системы - в спинномозговых ганглиях и в ганглиях черепно-мозговых нервов.
Эфферентные нейроны (центробежные) связаны с передачей нисходящих влияний от вышележащих этажей нервной системы к нижележащим или из центральной нервной системы к рабочим органам. Для эфферентных нейронов характерна разветвленная сеть дендритов и один длинный отросток - аксон.
Промежуточные нейроны (интернейроны, или вставочные) - это, как правило, более мелкие клетки, осуществляющие связь между различными (в частности, афферентными и эфферентными) нейронами. Благодаря многочисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов.
1. Строение и функции элементов нервной клетки
Различные структурные элементы нейрона имеют свои функциональные особенности и разное физиологическое значение. Нервная клетка состоит из тела, или сомы, и различных отростков. Многочисленные древовидно разветвленные отростки дендриты (от греч. dendron - дерево) служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток аксон (от греч. axis - ось), который передает нервные импульсы дальше - другой нервной клетке или рабочему органу (мышце, железе).
Форма нервной клетки, длина и расположение отростков чрезвычайно разнообразны и зависят от функционального назначения нейрона.
Среди нейронов встречаются самые крупные клеточные элементы организма. Размеры их поперечника колеблются от 6-7 мк (мелкие зернистые клетки мозжечка) до 70 мк (моторные нейроны головного и спинного мозга). Плотность их расположения в некоторых отделах центральной нервной системы очень велика. Например, в коре больших полушарий человека на 1мм 3 приходится почти 40 тыс. нейронов. Тела и дендриты нейронов коры занимают в целом примерно половину объема коры.
Внутренняя часть клетки заполнена цитоплазмой, в которой расположены ядро и различные органоиды. Цитоплазма очень богата ферментными системами (в частности, обеспечивающими гликолиз) и белком. Ее пронизывает сеть трубочек и пузырьков - эндоплазматический ретикулум. В цитоплазме имеются также отдельные зернышки - рибосомы и скопления этих зернышек - тельца Ниссля, представляющие собой белковые образования, содержащие до 50% РНК. Это белковые депо нейронов, где также происходит синтез белков и РНК.
В специальных аппаратах нервных клеток - митохондриях совершаются окислительные процессы с образованием богатых энергией соединений (макроэргических связей АТФ). Это энергетические станции нейрона. В них происходит трансформация энергии химических связей в такую форму, которая может быть использована нервной клеткой. Митохондрии концентрируются в наиболее активных частях клетки.
2. Обмен веществ в нейроне
Основной особенностью обмена веществ в нейроне является высокая скорость обмена и преобладание аэробных процессов. Потребность мозга в кислороде очень велика. Хотя вес мозга по отношению к весу тела составляет всего 2%, потребление кислорода мозгом достигает в состоянии покоя у взрослых людей 25% от общего его потребления организмом, а у маленьких детей - 50%. Даже кратковременное нарушение доставки кислорода кровью может вызвать необратимые изменения в деятельности нервных клеток: в спинном мозгу - через 20 - 30 мин., в стволе головного мозга - через 15 - 20 мин., а в коре больших полушарий - уже через 5 - 6 мин.
Энергетические траты мозга составляют 1 /6 - 1 /8 суточных затрат организма человека. Основным источником энергии для мозговой ткани является глюкоза. Мозг человека требует для обмена около 115г. глюкозы в сутки. Содержание ее в клетках мозга очень мало, и она постоянно черпается из крови.
Деятельное состояние нейронов сопровождается трофическими процессами - усилением в них синтеза белков. При различных воздействиях, вызывающих возбуждение нервных клеток, в том числе при мышечной тренировке, в их ткани значительно возрастает количество белка и РНК, при тормозных же состояниях и утомлении нейронов содержание этих веществ уменьшается. В процессе восстановления оно возвращается к исходному уровню или превышает его.
3. Глиальные клетки
В процессах питания нервных клеток и их обмене веществ участвуют также окружающие нейрон клетки глии (глиальные клетки, или нейроглия). Эти клетки заполняют в мозгу все пространство между нейронами. В коре больших полушарий их примерно в 5 раз больше, чем нервных клеток. Тела нейронов, как и их аксоны, окружены глиальными клетками. Глиальные клетки выполняют несколько функций: опорную, защитную, изолирующую, обменную (снабжение нейронов питательными веществами). Показано, что при длительном возбуждении в нейроне высокое содержание белка и нуклеиновых кислот поддерживается за счет клеток глии, в которых их количество соответственно уменьшается. В процессе восстановления после работы запасы белка и нуклеиновых кислот сначала нарастают в клетках глии, а затем и в цитоплазме нейрона.
Глиальные клетки обладают способностью перемещаться в пространстве по направлению к наиболее активным нейронам. Это наблюдается при различных афферентных раздражениях и при мышечной нагрузке. Например, уже через 20 мин. плавания у крыс было обнаружено увеличение числа глиальных клеток вокруг мотонейронов переднего рога спинного мозга.
Возможно, клетки глии участвуют в условно-рефлекторной деятельности мозга и в процессах памяти.
4. Основные функции нервной клетки
Основными функциями нервной клетки являются восприятие внешних раздражений (рецепторная функция), их переработка (интегративная функция) и передача нервных влияний на другие нейроны или различные рабочие органы (эффекторная функция).
Особенности осуществления этих функций позволяют разделить все нейроны центральной нервной системы на 2 большие группы:
1) клетки, передающие информацию на большие расстояния (из одного отдела центральной нервной системы в другой, от периферии к центру, от центров к исполнительному органу). Это крупные, афферентные и эфферентные нейроны, имеющие на своем теле и отростках большое количество синапсов, как возбуждающих, так и тормозящих, и способные к сложным процессам переработки поступающих через них влиянии;
2) клетки, обеспечивающие межнейроальные связи в пределах ограниченных нервных структур (промежуточные нейроны спинного мозга, коры больших полушарий и др.). Это мелкие клетки, воспринимающие нервные влияния только через возбуждающие синапсы. Эти клетки не способны к сложным процессам интеграции локальных синаптических влияний потенциалов, они служат передатчиками возбуждающих или тормозящих влияний на другие нервные клетки.
4.1. Воспринимающая функция нейрона
Все раздражения, поступающие в нервную систему, передаются на нейрон через определенные участки его мембраны, находящиеся в области синаптических контактов. В большинстве нервных клеток эта передача осуществляется химическим путем с помощью медиаторов. Ответом нейронов на внешнее раздражение является изменение величины, мембранного потенциала.
Нервный импульс
Строго говоря, движущееся по нервам возбуждение представляет собой нервные импульсы, а не потенциалы действия.
Можно сказать короче:
Но в физиологической литературе в качестве синонима для нервного импульса принято использовать также и термин "потенциал действия". Хотя потенциал действия - это только электрический компонент нервного импульса.
Потенциал действия – это резкое скачкообразное изменение мембранного потенциала с отрицательного на положительный и обратно.
Потенциал действия - это электрическая характеристика (электрическая составляющая) нервного импульса.
Нервный импульс - это сложный структурно-электро-химический процесс, распространяющийся по мембране нейрона в виде бегущей волны изменений.
Потенциал действия - это только электрический компонент нервного импульса, характеризующий изменения электрического заряда (потенциала) на локальном участке мембраны во время прохождения через него нервного импульса (от -70 до +30 мВ и обратно). (Кликните на изображение слева, чтобы увидеть анимацию.)
Сравните два приведённых выше рисунка (покликайте по ним) и, как говорится, почувствуйте разницу!
Где рождаются нервные импульсы?
Как ни странно, не все студенты, изучившие физиологию возбуждения, могут ответить на этот вопрос. ((
Хотя ответ не сложен. Нервные импульсы рождаются на нейронах всего в нескольких местах:
1) аксонный холмик (это переход тела нейрона в аксон),
2) рецепторное окончание дендрита,
3) первый перехват Ранвье на дендрите (триггерная зона дендрита),
4) постсинаптическая мембрана возбуждающего синапса.
Места возникновения нервных импульсов:
1. Аксонный холмик - главный породитель нервных импульсов.
Аксонный холмик - это самое начало аксона, там где он начинается на теле нейрона. Именно аксонный холмик является главным породителем (генератором) нервных импульсов на нейроне. Во всех остальных местах вероятность рождения нервного импульса намного меньше. Дело в том, что у мембраны аксонного холмика повышена чувствительность к возбуждению и понижен критический уровень деполяризации (КУД) по сравнению с остальными участками мембраны. Поэтому, когда на мембране нейрона начинают суммироваться многочисленные возбуждающие постсинаптические потенциалы (ВПСП), которые возникают в самых разных местах на постсинаптических мембранах всех его синаптических контактов, то раньше всего КУД достигается именно на аксонном холмике. Там-то эта сверхпороговая для холмика деполяризация и открывает потенциал-чувствительные натриевые каналы, в которые входит поток ионов натрия, порождающий потенциал действия и нервный импульс.
Итак, аксонный холмик является интегративной зоной на мембране, он интегрирует все возникающие на нейроне локальные потенциалы (возбуждающие и тормозные) - и первый срабатывает на достижение КУД, порождая нервный импульс.
Важно также учесть следующий факт. От аксонного холмика нервный импульс разбегается по всей мембране своего нейрона: как по аксону к пресинаптическоим окончаниям, так и по дендритам к постсинаптическим "начинаниям". Все локальные потенциалы при этом снимаются с мембраны нейрона и со всех его синапсов, т.к. они "перебиваются" потенциалом действия от пробегающего по всей мембране нервного импульса.
2. Рецепторное окончание чувствительного (афферентного) нейрона.
Если нейрон имеет рецепторное окончание, то на него может воздействовать адекватный раздражитель и порождать на этом окончании сначала генераторный потенциал, а затем и нервный импульс. Когда генераторный потенциал достигает КУД, то на этом окончании открываются потенциал-зависимые натриевые ионные каналы и рождается потенциал действия и нервный импульс. Нервный импульс бежит по дендриту к телу нейрона, а затем по его аксону к пресинаптическим окончаниям для передачи возбуждения на следующий нейрон. Так работают, к примеру, болевые рецепторы (ноцицепторы), являющиеся дендритными окончаниями болевых нейронов. Нервные импульсы в болевых нейронах вознимают именно на рецепторных окончаниях дендритов.
3. Первый перехват Ранвье на дендрите (триггерная зона дендрита).
Локальные возбуждающие постсинаптические потенциалы (ВПСП) на окончаниях дендрита, которые формируются в ответ на возбуждения, приходящие к дендриту через синапсы, суммируются на первом перехвате Ранвье этого дендрита, если он, конечно, миелинизирован. Там находится участок мембраны с повышенной чувствительностью к возбуждению (пониженным порогом), поэтому именно в этом участке легче всего преодолевается критический уровень деполяризации (КУД), после чего открываются потенциал-управляемые ионные каналы для натрия - и возникает потенциал действия (нервный импульс).
4. Постсинаптическая мембрана возбуждающего синапса.
В редких случаях ВПСП на возбуждающем синапсе может быть настолько силён, что прямо там же достигает КУД и порождает нервный импульс. Но чаще это бывает возможно только в результате суммации нескольких ВПСП: или с нескольких соседних синапсов, сработавших одновременно (пространственная суммация), или за счёт того, что на данный синапс пришло несколько импульсов подряд (временная суммация).
Видео: Проведение нервного импульса по нервному волокну
Потенциал действия как нервный импульс
Ниже размещён материал, взятый из учебно-методического пособия автора данного сайта, на который вполне можно ссылаться в своём списке литературы:
Сазонов В.Ф. Понятие и виды торможения в физиологии центральной нервной системы: Учебно-методическое пособие. Ч. 1. Рязань: РГПУ, 2004. 80 с.
Все процессы мембранных изменений, происходящих в ходе распространяющегося возбуждения, достаточно хорошо изучены и описаны в научной и учебной литературе. Но не всегда это описание легко понять, поскольку в данном процессе задействовано слишком много компонентов (с точки зрения обычного студента, а не вундеркинда, конечно).
Для облегчения понимания мы предлагаем рассматривать единый электрохимический процесс распространяющегося динамичного возбуждения с трех сторон, на трех уровнях:
Электрические явления – развитие потенциала действия.
Химические явления – движение ионных потоков.
Структурные явления – поведение ионных каналов.
Три стороны процесса распространяющегося возбуждения
1. Потенциал действия (ПД)
Потенциал действия – это скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно.
Обычно мембранный потенциал в нейронах ЦНС изменяется от –70 мВ до +30 мВ, а затем вновь возвращается к исходному состоянию, т.е. к –70 мВ. Как видим, понятие потенциала действия характеризуется через электрические явления на мембране.
На электрическом уровне изменения начинаются как смена поляризованного состояния мембраны на деполяризацию. Сначала деполяризация идет в виде локального возбуждающего потенциала. Вплоть до критического уровня деполяризации (примерно –50 мВ) это относительно простое линейное уменьшение электроотрицательности, пропорциональное силе воздействующего раздражителя. А вот потом начинается более крутая самоусиливающаяся деполяризация, она развивается не с постоянной скоростью, а с ускорением . Говоря образно, деполяризация так разгоняется, что перескакивает через нулевую отметку, не заметив этого, и даже переходит в положительную поляризацию. После достижения пика (обычно +30 мВ) начинается обратный процесс – реполяризация , т.е. восстановление отрицательной поляризации мембраны.
Кратко опишем электрические явления во время течения потенциала действия:
Восходящая ветвь графика:
потенциал покоя – исходное обычное поляризованное электроотрицательное состояние мембраны (–70 мВ);
нарастающий локальный потенциал – пропорциональная раздражителю деполяризация;
критический уровень деполяризации (–50 мВ) – резкое ускорение деполяризации (за счет самораскрытия натриевых каналов), с этой точки начинается спайк – высокоамплитудная часть потенциала действия;
самоусиливающаяся круто нарастающая деполяризация;
переход нулевой отметки (0 мВ) – смена полярности мембраны;
пик (+30 мВ) – вершина процесса изменения полярности мембраны, вершина потенциала действия.
Нисходящая ветвь графика:
реполяризация – восстановление прежней электроотрицательности мембраны;
переход нулевой отметки (0 мВ) – обратная смена полярности мембраны на прежнюю, отрицательную;
переход критического уровня деполяризации (–50 мВ) – прекращение фазы относительной рефрактерности (невозбудимости) и возврат возбудимости;
следовые процессы (следовая деполяризация или следовая гиперполяризация);
восстановление потенциала покоя – норма (–70 мВ).
Итак, сначала – деполяризация, затем – реполяризация. Сначала – утрата электроотрицательности, затем – восстановление электроотрицательности.
2. Ионные потоки
Итак, важно осознать, что все электрические токи, которые идут через мембрану, являются ионными потоками . Привычного нам из физики тока в виде потока электронов в клетках, как в водных системах, просто нет. Ссылки на потоки электронов будут ошибкой.
На химическом уровне мы, описывая распространяющееся возбуждение, должны рассмотреть, как изменяются характеристики ионных потоков, идущих через мембрану. Главное в этом процессе то, что при деполяризации резко усиливается поток ионов натрия внутрь клетки, а затем он внезапно прекращается на спайке потенциала действия. Входящий поток натрия как раз и вызывает деполяризацию, так как ионы натрия приносят с собой положительные заряды в клетку (чем и снижают электроотрицательность). Затем, после спайка, значительно нарастает выходящий наружу поток ионов калия, что вызывает реполяризацию. Ведь калий, как мы неоднократно говорили, выносит с собой из клетки положительные заряды. Отрицательные заряды остаются внутри клетки в большинстве, и за счет этого усиливается электроотрицательность. Это и есть восстановление поляризации за счет выходящего потока ионов калия. Заметим, что выходящий поток ионов калия возникает практически одновременно с появлением натриевого потока, но нарастает медленно и длится в 10 раз дольше. Несмотря на продолжительность калиевого потока самих ионов расходуется немного – всего одна миллионная доля от запаса калия в клетке (0,000001 часть).
Подведем итоги. Восходящая ветвь графика потенциала действия образуется за счет входа в клетку ионов натрия, а нисходящая – за счет выхода из клетки ионов калия.
3. Ионные каналы
Все три стороны процесса возбуждения – электрическая, химическая и структурная – необходимы для понимания его сущности. Но все-таки все начинается с работы ионных каналов. Именно состояние ионных каналов предопределяет поведение ионов, а поведение ионов в свою очередь сопровождается электрическими явлениями. Начинают процесс возбуждения натриевые каналы .
Пожар как метафора распространяющегося возбуждения
Когда мы искали образный пример, аналогию или метафору, которая может наглядно передать смысл распространяющегося возбуждения, то остановились на образе пожара. Действительно, распространяющееся возбуждение похоже на лесной пожар, когда горящие деревья остаются на месте, а фронт огня распространяется и уходит все дальше и дальше во все стороны от очага возгорания.
Как же в этой метафоре будет выглядеть явление торможения?
Ответ очевиден – торможение будет выглядеть как тушение пожара, как уменьшение горения и затухание огня. Но если огонь распространяется сам по себе, то тушение требует усилий. Из потушенного участка процесс тушения сам по себе не пойдет во все стороны.
Существует три варианта борьбы с пожаром: (1) либо надо ждать, когда все сгорит и огонь истощит все горючие запасы, (2) либо надо поливать водой горящие участки, чтобы они погасли, (3) либо надо поливать заранее ближайшие нетронутые огнем участки, чтобы они не загорелись.
АВТОВОЛНЫ В АКТИВНО-ВОЗБУДИМЫХ СРЕДАХ (АВС)
При распространении волны в активно-возбудимых средах не происходит переноса энергии. Энергия не переносится, а освобождается, когда до участка АВС доходит возбуждение. Можно провести аналогию с серией взрывов зарядов, заложенных на некотором расстоянии друг от друга (например, при тушении лесных пожаров, строительстве, мелиоративных работах), когда взрыв одного заряда вызывает взрыв рядом расположенного и так далее. Лесной пожар также является примером распространения волны в активно- возбудимой среде. Пламя распространяется по области с распределенными запасами энергии - деревья, валежник, сухой мох.
Основные свойства волн, распространяющихся в активно-возбудимых средах (АВС)
Волна возбуждения распространяется в АВС без затухания; прохождение волны возбуждения связано с рефрактерностью - невозбудимостью среды в течение некоторого промежутка времени (периода рефрактерности).
Видео: Потенциал действия (Action potential)
Законы раздражения возбудимости тканей
Закон силы
Возникновение распространяющегося возбуждения (ПД) возможно при условии, когда действующий на клетку раздражитель имеет некоторую минимальную (пороговую силу), иначе говоря, когда сила раздражителя соответствует порогу раздражения.
Порог раздражения – это та наименьшая величина раздражителя, которая действуя на клетку какое-то определенное время, способна вызвать максимальное возбуждение.
– наименьшая величина раздражителя, при действии которой потенциал покоя может сместиться до уровня критической деполяризации;
– критическая величина деполяризации клеточной мембраны, при которой активируется перенос ионов натрия внутрь клетки.

Рисунок 5. Возникновение местного потенциала при прохождении электрического тока, через участок нерва. Ток идет от анода к катоду (оба электрода – вне нерва) частично через пленку жидкости на поверхности нерва, а частично через оболочку нерва и в продольном направлении внутри волокна. Кривая внизу показывает вызываемое током изменения мембранного потенциала нервного волокна (по В. Катц)
Зависимость пороговой силы стимула от его длительности
Пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. Эта зависимость, открытая Гоорвегом, Вейсом, Лапиком получила название кривой “сила – длительность” или “сила – время”. Кривая “силы времени” имеет форму, близкую к равносторонней гиперболе и в первом приближении может быть описана эмпирической формулой:
где I – сила тока, Т – длительность его действия, а и b – постоянные, определяемые свойствами ткани.
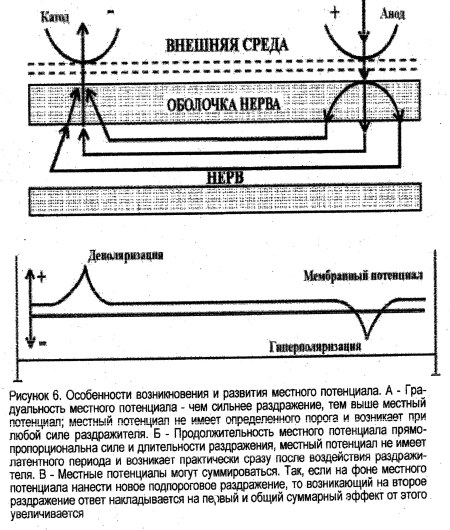
Рисунок 6. Особенности возникновения и развития местного потенциала. А – Градуальность местного потенциала – чем сильнее раздражение, тем выше местный потенциал; местный потенциал не имеет определенного порога и возникает при любой силе раздражителя. Б – Продолжительность местного потенциала прямопропорциональна силе и длительности раздражения, местный потенциал не имеет латентного периода и возникает практически сразу после воздействия раздражителя. В – Местные потенциалы могут суммироваться. Так, если на фоне местного потенциала нанести новое подпороговое раздражение, то возникающий на второе раздражение ответ накладывается на первый и общий суммарный эффект от этого увеличивается
Таким образом, го этой кривой вытекают два следствия:
1. Ток величиной ниже порога не вызывает возбуждения, как бы длительно он не действовал.
2. Какой бы сильный не был раздражитель, но если он действует очень короткое время, то возбуждение не возникает.
Реобаза – минимальная сила тока (или напряжение), способная вызвать возбуждение. Наименьшее время, в течение которого должен действовать стимул в одну реобазу, чтобы вызвать возбуждение – полезное время. Дальнейшее его увеличение не имеет значения для возникновения возбуждения.
Порог (реобаза) – величины непостоянные, зависят от функционального состояния клеток в покое, поэтому Лапик предложил определять более точный показатель – хронаксию.
Хронаксия – наименьшее время, в течение которого ток в две реобазы должен действовать на ткань, чтобы вызывать возбуждение.
Метод определения хронаксии – хронаксиметрия используется в клинике для диагностики повреждения нервных стволов и мышц.
Зависимость порога от крутизны нарастания раздражителя (аккомодация)
Порог раздражения имеет наименьшую величину при толчках электрического тока прямоугольной формы, когда сила нарастает очень быстро.
При уменьшении крутизны нарастания стимула ускоряются процессы инактивации натриевой проницаемости, приводящие к повышению порога и снижению амплитуды потенциалов действия. Чем круче должен нарастать ток, чтобы вызвать возбуждение, тем выше скорость аккомодации. Очень мала скорость аккомодации тех образований, которые склонны к автоматической деятельности (миокард, гладкие мышцы).
“Все” — на пороговые и сверхпороговые стимулы ответ максимальный и развивается потенциал действия; “ничего” – надо – пороговый стимул потенциал действия не развивается. Закон “все или ничего” установлен Боудичем в 1871 г. на мышце сердца: при подпороговой силе раздражения мышца сердца не сокращается, а при пороговой силе раздражения – сокращение максимально. При дальнейшем увеличении силы раздражения амплитуда сокращений не увеличивается.
Со временем была установлена и относительность этого закона. Оказалось, что “все” зависит от функционального состояния ткани (охлаждение, исходное растяжение мышцы и т. д.). С появлением микроэлектродной техники было установлено еще одно несоответствие: подпороговое раздражение вызывает местное, не распространяющееся возбуждение, следовательно, нельзя говорить, что допороговое раздражение не дает ничего. Процесс развития возбуждения подчиняется этому закону с уровня критической деполяризации, когда запускается лавинообразное поступление ионов калия в клетку.
Изменение возбудимости при возбуждении
Мера возбудимости – это порог раздражения. При местном, локальном, возбуждений возбудимость увеличивается. Потенциал действия сопровождается многофазными изменениями возбудимости.
1. Период повышенной возбудимости соответствует локальному ответу, когда мембранный потенциал достигает УКД, возбудимость повышена.
2. Период абсолютной рефрактерности соответствует фазе деполяризации потенциала действия, пику и началу фазы реполяризации, возбудимость снижена вплоть до полного отсутствия во время пика.
3. Период относительной рефрактерности соответствует оставшейся части фазы реполяризации, возбудимость постепенно восстанавливается к исходному уровню.
4. Супернормальный период соответствует фазе следовой деполяризации потенциала действия (отрицательный следовый потенциал), возбудимость повышена.
5. Субнормальный период соответствует фазе следовой гиперполяризации потенциала действия (положительный следовый потенциал), возбудимость снижена.

Рисунок 7. Изменения возбудимости нервного волокна в различные фазы потенциала действия и следовых изменений потенциала действия (по Б.И. Ходорову). Для наглядности длительность первых двух фаз на каждой кривой несколько увеличена. Пунктирной линией на рисунке А обозначает потенциал покоя, а на рисунке Б исходный уровень возбудимости
Закон лабильности или функциональной подвижности
Лабильность — скорость протекания физиологических процессов в возбудимой ткани. Например, можно творить о максимальной частоте раздражения, которую возбудимая ткань способна воспроизводить без трансформации ритма.
Мерой лабильности могут служить:
– длительность отдельного потенциала
– величина абсолютной рефрактерной фазы
– скорость восходящей и нисходящей фаз ПД.
Уровень лабильности характеризует скорость возникновения и компенсации возбуждения в любых клетках и уровень их функционального состояния. Можно измерять лабильность мембран, клеток, органов, причем, в системе из нескольких элементов (тканей, органов, образований) лабильность определяется по участку с наименьшей лабильностью:
Полярный закон раздражения (закон Пфлюгера)
Закон изменения мембранного потенциала при действии на возбудимые ткани постоянного электрического тока открыл Пфлюгер в 1859 г.
1. Постоянный ток проявляет свое раздражающее действие только в момент замыкания и размыкания цепи.
2. При замыкании цепи постоянного тока возбуждение возникает под катодом; при размыкании по анодом.
Изменение возбудимости под катодом.
При замыкании цепи постоянного тока под катодом (действуют допороговым, но продолжительным раздражителем) на мембране возникает стойкая длительная деполяризация, которая не связана с изменением ионной проницаемости мембраны, а обусловлена перераспределением ионов снаружи (они привносятся на электроде) и внутри – катион перемещается к катоду.
Вместе со смещением мембранного потенциала уровень критической деполяризации (УКД) смещается к нулю. При размыкании цепи постоянного тока под катодом мембранный потенциал быстро возвращается к исходному уровню, а УКД медленно, следовательно, порог увеличивается, возбудимость снижается и отмечается католическая депрессия Вериго. Таким образом, возбуждение возникает только при замыкании цепи постоянного тока под катодом.
Изменение возбудимости под анодом.
При замыкании цепи постоянного тока под анодом (действуют допороговым, но продолжительным раздражителем) на мембране развивается гиперполяризация за счет перераспределения ионов по обе стороны мембраны (без изменения ионной проницаемости мембраны) и возникающее за ней смещение уровня критической деполяризации в сторону мембранного потенциала. Следовательно, порог уменьшается, возбудимость повышается – анодическая экзальтация.
При размыкании цепи мембранный потенциал быстро восстанавливается к исходному уровню и достигает сниженного уровня критической деполяризации, генерируется потенциал действия. Таким образом, возбуждение возникает только при размыкании цепи постоянного тока под анодом. Сдвиги мембранного потенциала вблизи полюсов постоянного тока получили название электротонических. Сдвиги мембранного потенциала не связанные с изменением ионной проницаемости мембраны клетки называют пассивными.
Проведение возбуждения.
Потенциал действия – это волна возбуждения, распространяющаяся по мембранам нервных и мышечных клеток.
ПД обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам. Синоним ПД – нервный импульс или спайк. Сложная информация о действующих на организм раздражениях кодируется в виде отдельных групп потенциалов действия – рядов.
Согласно закону “все или ничего” амплитуда и длительность отдельных потенциалов действия постоянны, а частота и количество в ряду зависит от интенсивности раздражения. Такой способ кодирования информации и ее передачи является наиболее психоустойчивым.
В живых организмах информация может передаваться и гуморальным путем.
Преимущества ПД:
1. Информация более целенаправлена;
2. Передается быстро;
3. Адресат точно известен;
4. Информация может быть точнее закодирована.
ПД распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками. Из-за перезарядки мембраны во время генерации потенциала действия последний обладает способностью к самораспространению. Возникнув на одном участке, является стимулом для соседних. Наступающая после возбуждения в данном участке мембраны рефрактерностъ, обусловливает поступательное движение ПД.
Конкретные особенности распространения возбуждения связаны со строением мембраны клетки, нервных волокон. По мембранам мышечных клеток и в безмякотных нервных волокнах возбуждение распространяется непрерывно вдоль всей мембраны.
В волокнах, покрытых миелиновой оболочкой, потенциал действия может распространяться только скачкообразно (сальтаторно), перепрыгивая через участки волокна, покрытые шванновскими клетками, с одного перехвата Ранвье на другой.
Перехваты Ранвье представляют собой своеобразные станции ретрансляции, постоянно усиливающие сигнал, не давая ему угаснуть.
Причины сальтаторного проведения:
1. В перехватах Ранвье, свободных от миелина, сопротивление электрическому току минимально;
2. Порог раздражения в перехватах Ранвье минимальный;
3. Амплитуда ПД в каждом перехвате в 5 – 6 раз превышает порог в соседнем перехвате;
4. Велика плотность натриевых каналов на мембране перехвата.
Следовательно, возбуждение, возникающее в одном перехвате Ранвье, вызывает смещение электронов во внешней среде данного волокна и этого смещения достаточно, чтобы вызвать возбуждение в соседнем участке. Таким образом, скорость проведения возбуждения по нервному волокну зависит от диаметра волокон и наличия перехватов Ранвье.
Различают декрементное и бездекрементное распространение волны возбуждения.
ДЕКРЕМЕНТНОЕ проведение:
1. Наблюдается в безмиелиновых волокнах;
2. Скорость проведения невелика;
3. По мере удаления от места возникновения раздражающее действие местных токов постепенно уменьшается вплоть до полного угасания;
4. Свойственно волокнам, которые иннервируют внутренние органы, обладающие низкой функциональной активностью.
БЕЗДЕКРЕМЕНТНОЕ проведение:
1. ПД проходит весь путь от места раздражения до места реализации без затухания.
2. Характерно для миелиновых и тех безмиелиновых волокон, которые передают сигналы к органам, обладающими высокой реактивностью (сердце).
Распространение одиночного потенциала действия само по себе не требует энергетических затрат. Однако, восстановление исходного состояния мембраны и поддержание ее готовности к проведению нового импульса связано с затратой энергии.
Законы проведения возбуждения в нервах
Закон анатомической и физиологической непрерывности волокна.
Любая травма волокна нарушает проводимость. При действии новокаина (дикаина, кокаина) блокируются натриевые и калиевые каналы мембраны. Возникновение возбуждения и его проведение в этом случае становится невозможным.
Закон двустороннего проведения возбуждения
В целом организме по рефлекторной дуге возбуждение всегда распространяется в одном направлении: от рецептора к эффектору.
Причины:
1. Возбуждение всегда возникает при раздражении специфических рецепторов;
2. Рефрактерность во время возбуждения обусловливает поступательное движение;
3. В рефлекторной дуге возбуждение с одной нервной клетки на другую передастся в синапсах с помощью медиатора, который может выделяться только в одном направлении.
Закон изолированного проведения возбуждения в нервных стволах.
Передача возбуждения на большие расстояния невозможна из-за значительной потери тока во внеклеточной среде.
Физиология нейронов, глиальных клеток, рецепторов и синапсов
Классическая рефлекторная дуга состоит из:
– рецептора;
– афферентного пути (афферентного нейрона, который располагается в спинальном ганглии);
– нервного центра, где возбуждение с афферентного нейрона переходит на вставочную нервную клетку.
Затем возбуждение переходит на эффекторный орган (эффектор), в роли которого может выступать мышца. Многие нервные волокна покрыты глиальными клетками (миелиновая оболочка). Между этими Шванновскими клетками есть промежутки – перехваты Ранвье. Возбуждение с одного нейрона на другой и с мотонейрона на мышцу передается в синапсах с помощью медиатора.
Нервная клетка – структурная и функциональная единица ДНС, которая окружена клетками нейроглии.
Нейроглия (глиоциты) – совокупность всех клеточных элементов нервной ткани кроме нейронов.
В мозге взрослого человека 1150 – 200 млрд. глиальных клеток, что в 10 раз больше нервных.
Нейроглия:
1. макроглия:
– астроциты;
– олигодендроциты;
– эиендимоциты.
2. микроглия: глиальные макрофаги.
Астроциты составляют 45 – 60% серого вещества мозга. Покрывают 85% поверхности капилляров мозга (сосудистые ножки астроцитов), крупные отростки астроцитов контактируют с телами нейронов. Основная функция – трофическая.
Олигодендроциты образуют миелин в нервной системе к поддерживают его целостность.
Эпендимоциты – клетки, выстилающие стенки спинномозгового канала и всех желудочков головного мозга. Это граница между спинномозговой жидкостью (ликвор) и тканью мозга.
Функции нейроглии:
1. Опорная – вместе с сосудами и мозговыми оболочками образуют строму ткани мозга.
2. Трофическая – обеспечивают метаболизм нервных клеток (связь с кровеносными сосудами). В глиоцитах сосредоточен весь гликоген ЦНС.
3. Участие в интегративной деятельности мозга:
– формирование следов воздействия (память), а значит и условного рефлекса;
– без глиоцитов (блокада антиглиальным гамма-глобулином) меняется электрическая активность нейронов.
Особенности глиальных клеток:
1. Более чувствительны к ионным изменениям среды;
2. Высокая активность калий – натриевой АТФ-азы;
3. Высокая проницаемость для ионов калия;
4. Мембранный потенциал равен 90 мВ, у нейронов 60 – 80 мВ;
5. На раздражение отвечает только медленной деполяризацией не более 10 мВ;
6. Потенциал действия в глиальных клетках не генерируется.
Читайте также:
