
Проведение сигнала по нерву
Нервная система человека выступает своеобразным координатором в нашем организме. Она передаёт команды от мозга мускулатуре, органам, тканям и обрабатывает сигналы, идущие от них. В качестве своеобразного носителя данных используется нервный импульс. Что он собой представляет? С какой скоростью работает? На эти, а также на ряд других вопросов можно будет найти ответ в этой статье.
Чем является нервный импульс?
Исследование строения и работы
Впервые прохождение нервного импульса было продемонстрировано немецкими учеными Э. Герингом и Г. Гельмгольцем на примере лягушки. Тогда же и было установлено, что биоэлектрический сигнал распространяется с указанной ранее скоростью. Вообще, такое является возможным благодаря особенному построению нервных волокон. В некотором роде они напоминают электрический кабель. Так, если проводить параллели с ним, то проводниками являются аксоны, а изоляторами – их миелиновые оболочки (они являют собой мембрану шванновской клетки, которая намотана в несколько слоев). Причем скорость нервного импульса зависит в первую очередь от диаметра волокон. Вторым по важности считается качество электрической изоляции. Кстати, в качестве материала организмом используется липопротеид миелин, который обладает свойствами диэлектрика. При прочих равных условиях, чем больше будет его слой, тем быстрее будут проходить нервные импульсы. Даже на данный момент нельзя сказать, что эта система полноценно исследована. Многое, что относится к нервам и импульсам, ещё остаётся загадкой и предметом исследования.
Особенности строения и функционирования
Где они создаются?
Типы клеток
- Рецепторные (чувствительные). Ими кодируются и превращаются в нервные импульсы все температурные, химические, звуковые, механические и световые раздражители.
- Вставочные (также называются кондукторными или замыкательными). Они служат для того, чтобы перерабатывать и переключать импульсы. Наибольшее их число находится в головном и спинном мозге человека.
- Эффекторные (двигательные). Они получают команды от центральной нервной системы на то, чтобы были совершены определённые действия (при ярком солнце закрыть рукой глаза и так далее).
Каждый нейрон имеет тело клетки и отросток. Путь нервного импульса по телу начинается именно с последнего. Отростки бывают двух типов:
- Дендриты. На них возложена функция восприятия раздражения расположенных на них рецепторов.
- Аксоны. Благодаря им нервные импульсы передаются от клеток к рабочему органу.
Интересный аспект деятельности
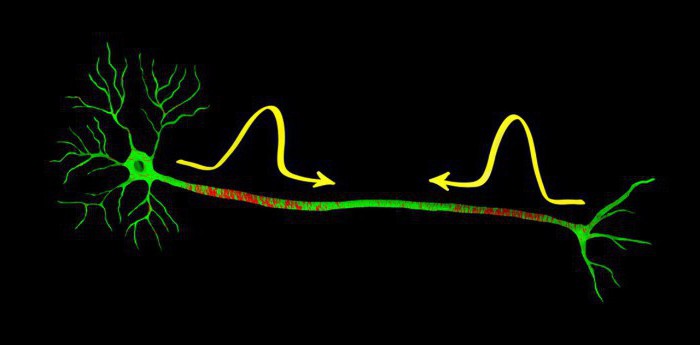
О потенциале действия
Как всё работает в мозгу?
Работа нейромедиаторов
Когда они передают нервные импульсы, то существует несколько вариантов, что произойдёт с ними:
- Они будут диффундированы.
- Подвергнутся химическому расщеплению.
- Вернутся назад в свои пузырьки (это называется обратным захватом).
В конце 20-го века сделали поразительное открытие. Ученые узнали, что лекарства, что влияют на нейромедиаторы (а также их выброс и обратный захват), могут изменять психическое состояние человека коренным образом. Так, к примеру, ряд антидепрессантов вроде "Прозака" блокируют обратный захват серотонина. Есть определённые причины считать, что в болезни Паркинсона виноват дефицит в головном мозге нейромедиатора дофамина.
Если кратко, то они могут работать с тысячами нейромедиаторов, которые посылаются их соседями. Детали относительно обработки и интеграции данного типа импульсов нам почти не известны. Хотя над этим работает много исследовательских групп. На данный момент получилось узнать, что все полученные импульсы интегрируются, а нейрон выносит решение – необходимо ли поддерживать потенциал действия и передавать их дальше. На этом фундаментальном процессе базируется функционирование головного мозга человека. Ну что ж, тогда это неудивительно, что мы не знаем ответа на эту загадку.
Некоторые теоретические особенности
Где же создаются нервные импульсы?
Откуда они начинают свой путь? Ответ на этот вопрос может дать любой студент, который прилежно изучал физиологию возбуждения. Есть четыре варианта:
- Рецепторное окончание дендрита. Если оно есть (что не факт), то возможным является наличие адекватного раздражителя, что создаст сначала генераторный потенциал, а потом уже и нервный импульс. Подобным образом работают болевые рецепторы.
- Мембрана возбуждающего синапса. Как правило, такое возможно только при наличии сильного раздражения или их суммирования.
- Триггерная зона дентрида. В этом случае локальные возбуждающие постсинаптические потенциалы формируются как ответ на раздражитель. Если первый перехват Ранвье миелинизирован, то они на нём суммируются. Благодаря наличию там участка мембраны, которая обладает повышенной чувствительностью, здесь возникает нервный импульс.
- Аксонный холмик. Так называют место, где начинается аксон. Холмик – это наиболее частый создать импульсов на нейроне. Во всех остальных местах, которые рассматривались ранее, их возникновение гораздо менее вероятное. Это происходит из-за того, что здесь мембрана имеет повышенную чувствительность, а также пониженный критический уровень деполяризации. Поэтому, когда начинается суммирование многочисленных возбуждающих постсинаптических потенциалов, то раньше всего на них реагирует холмик.
Пример распространяющегося возбуждения
Вспомните сводки из новостей прошлого лета (также это скоро можно будет услышать опять). Пожар распространяется! При этом деревья и кустарники, которые горят, остаются на своих местах. А вот фронт огня идёт всё дальше от места, где был очаг возгорания. Аналогичным образом работает нервная система.
Часто бывает необходимо успокоить начавшееся возбуждение нервной системы. Но это не так легко сделать, как и в случае с огнем. Для этого совершают искусственное вмешательство в работу нейрона (в лечебных целях) или используют различные физиологические средства. Это можно сравнить с заливанием пожара водой.

Кандидат биологических наук Л. Чайлахян, научный сотрудник Института биофизики АН СССР
Мозг человека, без сомнения, высшее достижение природы.
Велика и заманчива цель, но неимоверно сложен объект исследования. Шутка сказать, этот килограмм ткани представляет собой сложнейшую систему связи десятков миллиардов нервных клеток.
Однако первый существенный шаг к познанию работы мозга уже сделан. Может быть, он один из самых легких, но он чрезвычайно важен для всего дальнейшего.
Я имею в виду исследование механизма передачи нервных импульсов — сигналов, бегущих по нервам, как по проводам. Именно эти сигналы являются той азбукой мозга, с помощью которой органы чувств посылают в центральную нервную систему сведения-депеши о событиях во внешнем мире. Нервными импульсами зашифровывает мозг свои приказы мышцам и различным внутренним органам. Наконец, на языке этих сигналов говорят между собой отдельные нервные клетки и нервные центры.
В проблеме изучения механизма нервного импульса и его распространения можно выделить два основных вопроса: природа проведения нервного импульса или возбуждения в пределах одной клетки — по волокну и механизм передачи нервного импульса от клетки к клетке — через синапсы.
Какова природа сигналов, передающихся от клетки к клетка по нервным волокнам?
Этой проблемой человек интересовался уже давно, Декарт предполагал, что распространение сигнала связано с переливанием жидкости по нервам, как по трубкам. Ньютон думал, что это чисто механический процесс. Когда появилась электромагнитная теория, ученые решили, что нервный импульс аналогичен движению тока по проводнику со скоростью, близкой к скорости распространения электромагнитных колебаний. Наконец, с развитием биохимии появилась точка зрения, что движение нервного импульса — это распространение вдоль по нервному волокну особой биохимической реакции.
И всё же ни одно из этих представлений не оправдалось.
В настоящее время природа нервного импульса раскрыта: это удивительно тонкий электрохимический процесс, в основе которого лежит перемещение ионов через оболочку клетки.
Большой вклад в раскрытие этой природы внесли работы трех ученых: Алана Ходжкина, профессора биофизики Кембриджского университета; Эндрью Хаксли, профессора физиологии Лондонского университета, и Джона Экклса, профессора физиологии австралийского университета в Канберре. Им присуждена Нобелевская премия в области медицины за 1963 год,
Впервые предположение об электрохимической природе нервного импульса высказал известный немецкий физиолог Бернштейн в начале нашего столетия.
К началу двадцатого века было довольно многое известно о нервном возбуждении. Ученые уже знали, что нервное волокно можно возбудить электрическим током, причем возбуждение всегда возникает под катодом — под минусом. Было известно, что возбужденная область нерва заряжается отрицательно по отношению к невозбужденному участку. Было установлено, что нервный импульс в каждой точке длится всего 0,001—0,002 секунды, что величина возбуждения не зависит от силы раздражения, как громкость звонка в нашей квартире не зависит от того, как сильно мы нажимаем на кнопку. Наконец, ученые установили, что носителями электрического тока в живых тканях являются ионы; причем внутри клетки основной электролит — соли калия, а в тканевой жидкости — соли натрия. Внутри большинства клеток концентрация ионов калия в 30—50 раз больше, чем в крови и в межклеточной жидкости, омывающей клетки.
И вот на основании всех этих данных Бернштейн предположил, что оболочка нервных и мышечных клеток представляет собой особую полупроницаемую мембрану. Она проницаема только для ионов К + ; для всех остальных ионов, в том числе и для находящихся внутри клетки отрицательно заряженных анионов, путь закрыт. Ясно, что калий по законам диффузии будет стремиться выйти из клетки, в клетке возникает избыток анионов, и по обе стороны мембраны появится разность потенциалов: снаружи — плюс (избыток катионов), внутри — минус (избыток анионов). Эта разность потенциалов получила название потенциала покоя. Таким образом, в покое, в невозбужденном состоянии внутренняя часть клетки всегда заряжена отрицательно по сравнению с наружным раствором.
Бернштейн предположил, что в момент возбуждения нервного волокна происходят структурные изменения поверхностной мембраны, ее поры как бы увеличиваются, и она становится проницаемой для всех ионов. При этом, естественно, разность потенциалов исчезает. Это и вызывает нервный сигнал.
Мембранная теория Бернштейма быстро завоевала признание и просуществовала свыше 40 лет, вплоть до середины нашего столетия.
Но уже в конце 30-х годов теория Бернштейна встретилась с непреодолимыми противоречиями. Сильный удар ей был нанесен в 1939 году тонкими экспериментами Ходжкина и Хаксли. Эти ученые впервые измерили абсолютные величины мембранного потенциала нервного волокна в покое и при возбуждении. Оказалось, что при возбуждении мембранный потенциал не просто уменьшался до нуля, а переходил через ноль на несколько десятков милливольт. То есть внутренняя часть волокна из отрицательной становилась положительной.
Но мало ниспровергнуть теорию, надо заменить ее другой: наука не терпит вакуума. И Ходжкин, Хаксли, Катц в 1949—1953 годах предлагают новую теорию. Она получает название натриевой.
Здесь читатель вправе удивиться: до сих пор о натрии не было речи. В этом все и дело. Ученые установили с помощью меченых атомов, что в передаче нервного импульса замешаны не только ионы калия и анионы, но и ионы натрия и хлора.
В организме достаточно ионов натрия и хлора, все знают, что кровь соленая на вкус. Причем натрия в межклеточной жидкости в 5—10 раз больше, чем внутри нервного волокна.
Что же это может означать? Ученые предположили, что при возбуждении в первый момент резко увеличивается проницаемость мембраны только для натрия. Проницаемость становится в десятки раз больше, чем для ионов калия. А так как натрия снаружи в 5—10 рез больше, чем внутри, то он будет стремиться войти в нервное волокно. И тогда внутренняя часть волокна станет положительной.
А через какое-то время — после возбуждения — равновесие восстанавливается: мембрана начинает пропускать и ионы калия. И они выходят наружу. Тем самым они компенсируют тот положительный заряд, который был внесен внутрь волокна ионами натрия.
Совсем нелегко было прийти к таким представлениям. И вот почему: диаметр иона натрия в растворе раза в полтора больше диаметра ионов калия и хлора. И совершенно непонятно, каким образом больший по размеру ион проходит там, где не может пройти меньший.
Нужно было решительно изменить взгляд на механизм перехода ионов через мембраны. Ясно, что только рассуждениями о порах в мембране здесь не обойтись. И тогда была высказана идея, что ионы могут пересекать мембрану совершенно другим способом, с помощью тайных до поры до времени союзников — особых органических молекул-переносчиков, спрятанных в самой мембране. С помощью такой молекулы ионы могут пересекать мембрану в любом месте, а не только через поры. Причем эти молекулы-такси хорошо различают своих пассажиров, они не путают ионы натрия с ионами калия.
Интересно, что нервные волокна тратят на свою основную работу — проведение нервных импульсов — всего около 15 минут в сутки. Однако готовы к этому волокна в любую секунду: все элементы нервного волокна работают без перерыва — 24 часа в сутки. Нервные волокна в этом смысле подобны самолетам-перехватчикам, у которых непрерывно работают моторы для мгновенного вылета, однако сам вылет может состояться лишь раз в несколько месяцев.
Мы познакомились сейчас с первой половиной таинственного акта прохождения нервного импульса — вдоль одного волокна. А как же передается возбуждение от клетки к клетке, через места стыков — синапсы. Этот вопрос был исследован в блестящих опытах третьего нобелевского лауреата, Джона Экклса.
Возбуждение не может непосредственно перейти с нервных окончаний одной клетки на тело или дендриты другой клетки. Практически весь ток вытекает через синаптическую щель в наружную жидкость, и в соседнюю клетку через синапс попадает ничтожная его доля, неспособная вызвать возбуждение. Таким образом, в области синапсов электрическая непрерывность в распространении нервного импульса нарушается. Здесь, на стыке двух клеток, в силу вступает совершенно другой механизм.
Когда возбуждение подходит к окончанию клетки, к месту синапса, в межклеточную жидкость выделяются физиологически активные вещества — медиаторы, или посредники. Они становятся связующим звеном в передаче информации от клетки к клетке. Медиатор химически взаимодействует со второй нервной клеткой, изменяет ионную проницаемость ее мембраны — как бы пробивает брешь, в которую устремляются многие ионы, в том числе и ионы натрия.
Итак, благодаря работам Ходжкина, Хаксли и Экклса важнейшие состояния нервной клетки — возбуждение и торможение — можно описать в терминах ионных процессов, в терминах структурно-химических перестроек поверхностных мембран. На основании этих работ уже можно делать предположения о возможных механизмах кратковременной и долговременной памяти, о пластических свойствах нервной ткани. Однако это разговор о механизмах в пределах одной или нескольких клеток. Это лишь, азбука мозга. По-видимому, следующий этап, возможно, гораздо более трудный, — вскрытие законов, по которым строится координирующая деятельность тысяч нервных клеток, распознание языка, на котором говорят между собой нервные центры.
Мы сейчас в познании работы мозга находимся на уровне ребенка, который узнал буквы алфавита, но не умеет связывать их в слова. Однако недалеко время, когда ученые с помощью кода — элементарных биохимических актов, происходящих в нервной клетке, прочтут увлекательнейший диалог между нервными центрами мозга.
Детальное описание иллюстраций
Специалисты, работающие в клинических нейрофизиологических лабораториях, преимущественно имеют дело с двумя группами вопросов: оценкой функционального состояния периферической нервной системы (ПНС) и оценкой состояния коры головного мозга. Для оценки состояния ПНС применяют методы исследования нервной проводимости (ИНП). При этом исследуемый нерв стимулируют электрическим током с одновременной регистрацией электрических волн, которые нерв генерирует в ответ на раздражение. Второй метод оценки состояния ПНС — электромиография (ЭМГ), при которой в исследуемых мышцах регистрируют волны, возникающие при их произвольном сокращении. Комплекс методов ИНП и ЭМГ называют электродиагностическим обследованием.
При подозрении на поражение ПНС после клинического осмотра проводят исследование нервной проводимости (ИНП). С помощью электрической стимуляции нерва и последующей регистрации деполяризации связанного с этим нервом мышечного волокна можно определить, поражает ли заболевание нерв, нервно-мышечный синапс или саму мышцу. ИНП также позволяет установить, является ли заболевание фокальным или диффузным, повреждает чувствительные и/или двигательные аксоны, поражает миелиновую оболочку или сам аксон.
а) Исследование проведения по нервам верхней конечности. Основной нерв верхней конечности, исследование проводимости которого позволяет выявить наличие фокального (в отличие от генерализованного) поражения периферической нервно-мышечной системы,— срединный нерв. Срединный нерв — одновременно двигательный и чувствительный — обладает тремя основными преимуществами в плане проведения электрофизиологической диагностики.
1. В области локтевого сустава и запястья он расположен близко к поверхности, поэтому технически просто проводить стимуляцию нерва и выполнять регистрацию его реакции.
2. Срединный нерв иннервирует короткую мышцу, отводящую большой палец кисти, которая хорошо подходит для проведения поверхностной и игольчатой ЭМГ (двигательное ИНП).
1. Проведение по двигательным нервам:
- Стимуляция. Обычный стимулирующий электрод имеет катод и анод в форме двух тупых штырей, которые прикладывают к поверхности кожи над нервом. На рисунке ниже изображен электрод, размещенный над срединным нервом у запястья (сразу латеральнее сухожилия длинной ладонной мышцы). Для того чтобы анод не заблокировал проведение нервного импульса, катод должен быть расположен к месту регистрации ближе, чем анод. Когда между катодом и анодом возникает электрический ток достаточной силы, изменение трансмембранного тока ионов вызывает появление нервного импульса, который распространяется по нерву в обоих направлениях. Сначала деполяризация возникает на крупных миелинизированных нервных волокнах, расположенных вблизи катода; к ним относят Аα-аксоны мотонейронов переднего рога спинного мозга. Обычно стимул в 20-40 mА продолжительностью 0,1 мс достаточен для возбуждения всех двигательных единиц короткой мышцы, отводящей большой палец.
- Регистрация. Активный поверхностный датчик, имеющий в нашем случае форму диска, устанавливают к двигательной точке. Это место, расположенное в центре исследуемой мышцы, имеет наибольшую концентрацию двигательных концевых пластинок. Второй, референтный электрод устанавливают на нейтральную область на некотором удалении от первого электрода. Для записи разницы потенциалов между этими двумя электродами используют усилитель, который также увеличивает выраженность вызванной двигательной реакции мышцы. Система настроена таким образом, что при регистрации активным электродом отрицательного значения кривая на мониторе отклоняется вверх.
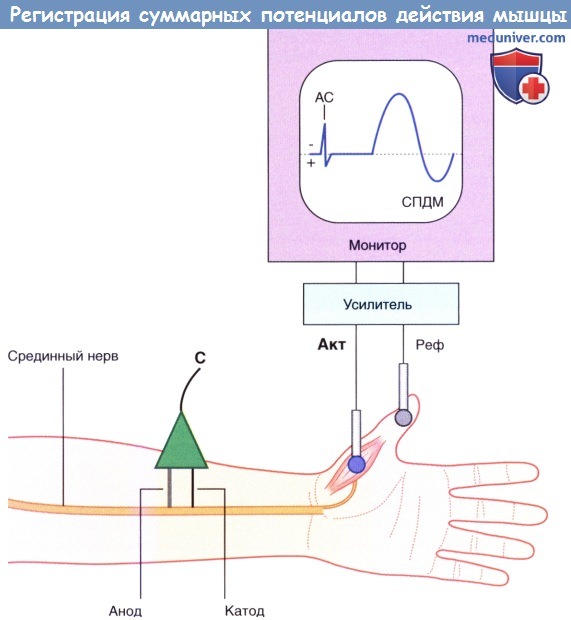
Базовая установка для регистрации СПДМ. АС — артефакт стимуляции.
Стимулирующий электрод (С) расположен над срединным нервом.
Активный электрод (Акт) расположен над короткой мышцей, отводящей большой палец.
Референтный электрод (Реф) установлен дистальнее.
При подаче слабого стимула на мониторе будет отображаться только плоская линия, которая иногда прерывается стимуляционными артефактами. При увеличении интенсивности стимула появляются небольшие суммарные потенциалы действия мышцы (СПДМ). Они возникают за счет активации крупных миелинизированных аксонов, расположенных вблизи стимулирующего электрода. Волна деполяризации, передающаяся по аксону, приводит к деполяризации всех мышц, которые иннервирует этот аксон. В двигательных единицах внутренних мышц руки, в том числе в короткой мышце, отводящей большой палец, один мотонейрон иннервирует 200-300 мышечных волокон. Для крупных мышц, которые не совершают точных мелких движений (например, дельтовидная, икроножная), минимальное отклонение кривой на мониторе будет в несколько раз больше. Это связано с двумя факторами: соотношение мотонейронов к двигательным волокнам составляет примерно 1:1000, а их крупные мышечные волокна генерируют потенциалы действия большей амплитуды.
Следует еще раз подчеркнуть, что кривая, возникающая на мониторе, не отображает сокращения мышц, а представляет результат внеклеточного потенциала, который возникает за счет деполяризации мембран мышечных клеток и передается через подкожные ткани и кожу. Несмотря на это, при большинстве мышечных заболеваний патологический процесс также будет нарушать процесс деполяризации клеточной мембраны, что приведет к появлению аномалий на кривой.
При повышении вольтажа происходит возбуждение дополнительных двигательных единиц. Это происходит до тех пор, пока все двигательные единицы не будут активироваться одним импульсом. Такой стимул называют максимальным. Для точности окончательный стимул обычно делают супрамаксималъным, т.е. на 5-10 % превышающим максимальный. Окончательная наблюдаемая волна и представляет собой СПДМ — сумму потенциалов действия отдельных мышечных волокон.
Результат измерения конечного СПДМ изображен на рисунке ниже. К измеряемым параметрам относят латентность (временной интервал) между временем подачи стимула и началом деполяризации, амплитуду и длительность отрицательной фазы волны. (Появление последней, положительной фазы, обусловлено движением ионов внутрь клетки во время общей реполяризации мышечных волокон.)
- Повторное измерение. Целесообразно повторно измерить СПДН, но уже с другого нерва. Обычно для повторного измерения используют локтевой нерв. Точка С1 расположена на запястье сразу латеральнее локтевого сгибателя запястья, а С2 — на уровне выхода нерва у медиального надмыщелка. Активный электрод устанавливают над мышцами гипотенара у медиального края ладони.
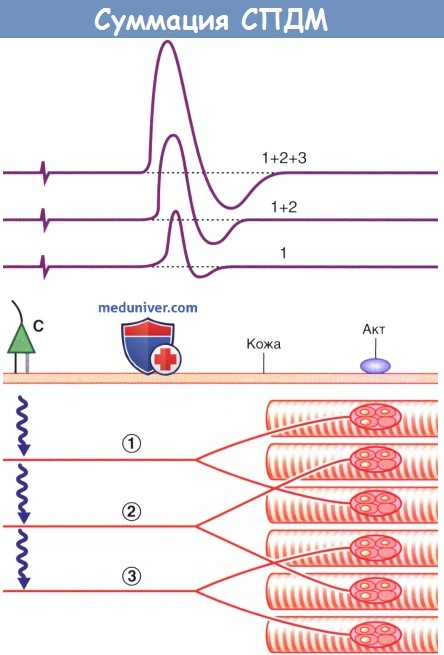
Суммация СПДМ. Двигательные единицы представлены перекрещивающимися парами мышечных волокон.
При легкой (1), средней (2) и максимальной (3) стимуляции на экране возникают волны все большей амплитуды, хотя они и являются разными с точки зрения физиологии феноменами. 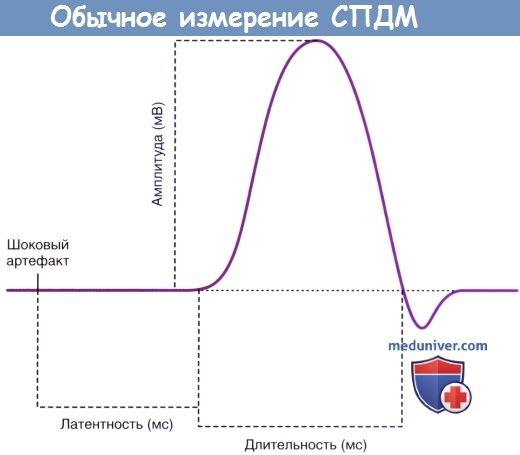
Обычное измерение СПДМ 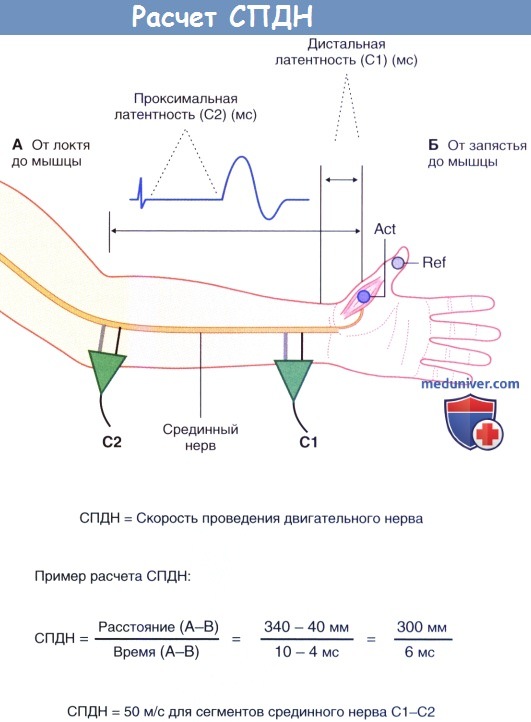
Расчет скорости проведения двигательного нерва (СПДН). Нерв стимулируют дважды: С1—первый стимул, С2 — второй стимул.
Двунаправленными стрелками отмечены два измерения длины.
Исходное значение времени не представлено. Внизу показан пример расчета нормальной скорости проведения.
2. Проведение по чувствительным нервам. Для измерения скорости проведения чувствительных нервов (СПЧН) также удобнее всего использовать срединный нерв. Точно также при стимуляции происходит возбуждение крупных миелинизированных нервных волокон, места и характер стимуляции в локтевой ямке и у запястья остаются прежними. Однако в данном случае избирательно записывают антидромные потенциалы кожных чувствительных волокон, в частности пальцевых ветвей срединного нерва, идущих к коже указательного пальца. Для этого на указательный палец надевают активное записывающее устройство в форме кольца.
1. Физиологическая временная дисперсия. Подобно тому, как во время гонки расстояние между бегунами прогрессивно увеличивается, наиболее быстрый проводник импульса выходит вперед, а самый медленный остается позади. Из-за этого при увеличении расстояния измерения кривая СЧПД удлиняется. Данное явление и называют временной дисперсией (рассеивание во времени).
- Скорость проведения чувствительных нервов. Основные режимы и принципы измерения остаются теми же, что и при исследовании суммарных потенциалов действия мышцы (СПДМ). На рисунке ниже изображен пример, на котором показан феномен отмены фазы при наличии физиологической временной дисперсии.
- Повторное измерение. Обычно проводят с локтевым нервом. Как и ранее, нерв стимулируют у запястья и у локтевой ямки, кольцо надевают на мизинец.
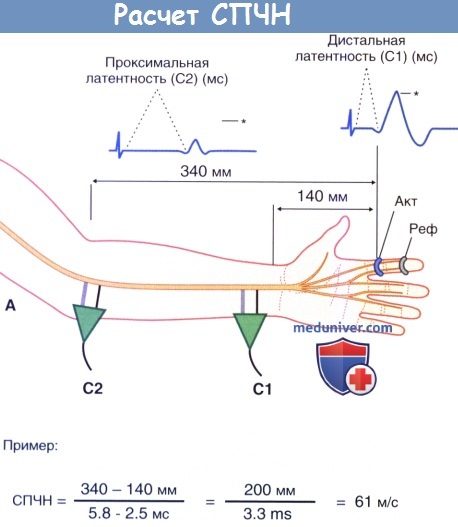
Расчет скорости проведения чувствительных нервов (СПЧН). Представлены пальцевые ветви срединного нерва. Основные принципы расчета те же, что и в случае скорости проведения двигательного нерва (СПДН).
б) Проведение по нервам нижней конечности:
1. Проведение по двигательным нервам. При исследовании СПДМ на нижней конечности чаще всего используют глубокий малоберцовый нерв, регистрацию осуществляют с короткого разгибателя пальцев на подошвенной поверхности стопы. Сначала глубокий малоберцовый нерв стимулируют на передней поверхности лодыжки, а затем на уровне шейки малоберцовой кости. Иногда регистрацию осуществляют также с передней большеберцовой мышцы; в этом случае общий малоберцовый нерв сначала стимулируют у шейки малоберцовой кости, а затем у латерального края подколенной ямки вблизи сухожилия двуглавой мышцы бедра.
Повторное исследование СПДМ проводят со стимуляцией большеберцового нерва, запись осуществляют с мышцы, приводящей большой палец, расположенной у медиального края стопы.
2. Проведение по чувствительным нервам. Для исследования СЧПД используют икроножный нерв. Он начинается от большеберцового нерва и получает ветвь от общего малоберцового нерва; данный нерв иннервирует кожу вдоль латерального края стопы. Запись осуществляют с кожи, расположенной немного ниже латеральной лодыжки, далее нерв антидромно стимулируют в точках, показанных на рисунке ниже.
• на уровне шеи, где корешки спинномозговых нервов С6 и С7 предрасположены к сдавливанию остеофитами, возникающими при шейном спондилезе;
• на уровне поясницы, где корешки спинномозгового нерва S1 могут сдавливаться при пролапсе межпозвоночного диска L5/S1;
• радикулопатия может быть компонентом генерализованной периферической нейропатии.
Н-рефлекс. Поскольку корешки спинномозговых нервов залегают глубоко, оценить проводимость по ним можно лишь косвенно за счет активации сенсомоторных чувствительных дуг соответствующего уровня. Стандартный тест, названный в честь Hoffman, который впервые его описал, называют Н-ответом или Н-рефлексом. Его часто используют для оценки общей скорости проведения по рефлекторной дуге S1; оценивают те же самые нейроны, которые клинически исследуют путем ахиллова рефлекса. Большеберцовый нерв стимулируют длительным током минимальной силы, достаточной для сокращения мышцы. Цель исследования — возбуждение самых крупных афферентных миелинизированных волокон (предпочтительно длительными стимулами), преимущественно тех, которые иннервируют аннулоспиральные окончания мышечных веретен. За счет этого активируется моносинаптический рефлекс с минимальной латентностью, происходит сокращение трехглавой мышцы голени (икроножной/камбаловидной). Минимальная латентность оказывается достаточно долгой — до 35 мс в зависимости от роста пациента, поскольку сегмент S1 спинного мозга расположен за телом позвонка L1; за счет этого нервный импульс должен пройти путь порядка 130-150 см вниз и вверх. При последовательном повышении силы тока возникает точка, в которой появляются М-волны. М-волны возникают благодаря прямой ортодром-ной активации двигательных концевых пластинок. Антидромное проведение обусловливает постепенное затухание потенциалов действия, нисходящих по эфферентному звену Н-рефлекса.
На верхней конечности проведение по нервным корешкам нерва С6 можно исследовать за счет стимуляции срединного нерва и регистрации сокращений лучевого сгибателя запястья. Корешок С7 можно исследовать путем стимуляции заднего кожного нерва предплечья и регистрации сокращений трехглавой мышцы плеча.
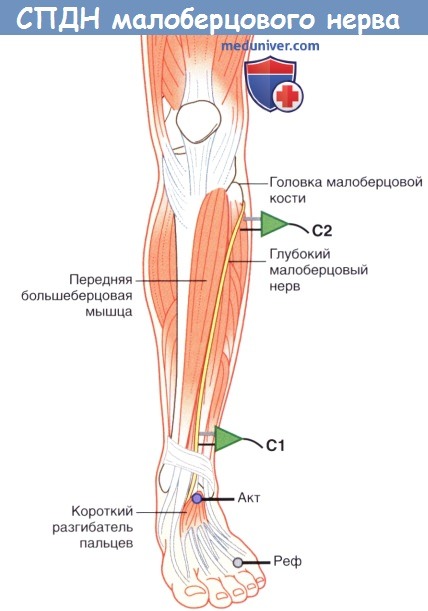
Положение датчика и усилителя при измерении скорости проведения двигательного нерва (СПДН) малоберцового нерва. 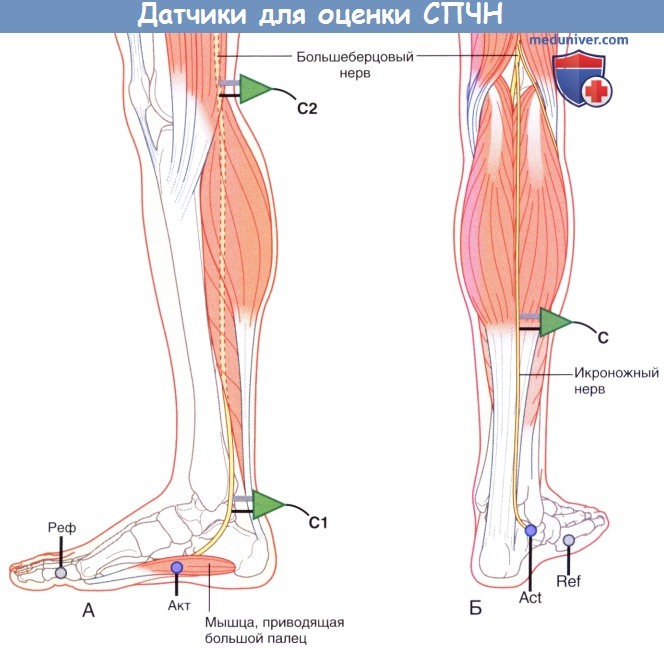
(А) Положение датчика и стимулятора при измерении скорости проведения чувствительного нерва (СПЧН) с большеберцового нерва.
(Б) Положение электродов при записи антидромной скорости проведения чувствительного нерва (СПЧН) с икроножного нерва. 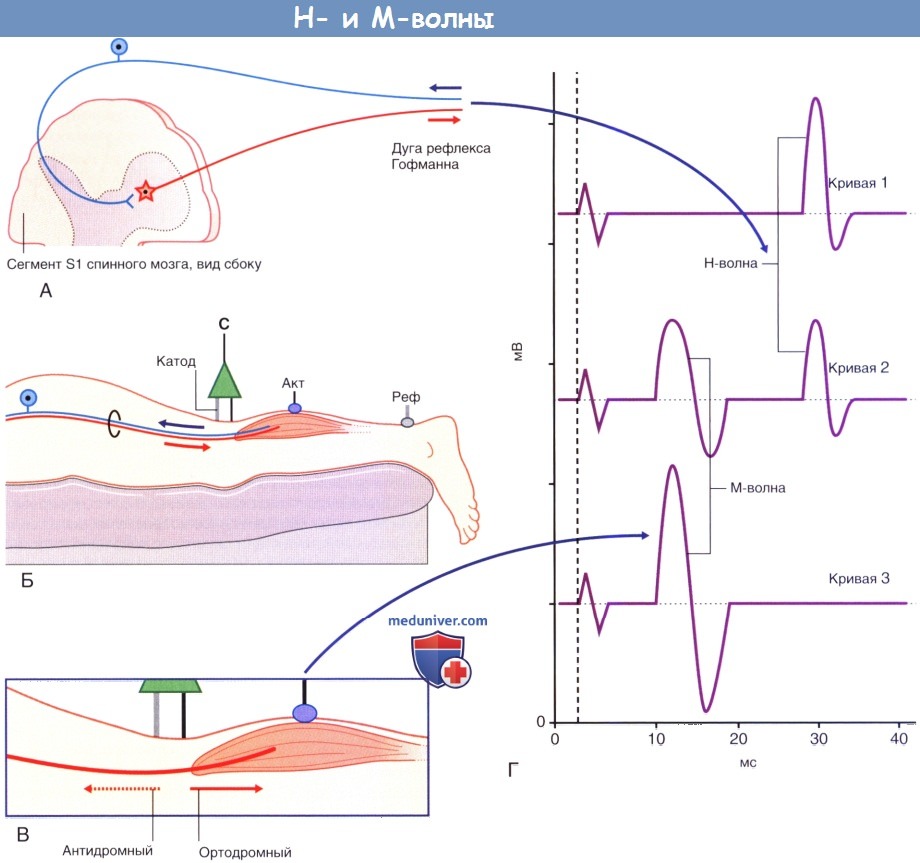
Анатомические предпосылки появления Н- и М-волн.
(А) Появление Н-волны опосредованно моносинаптической рефлекторной дугой, как показано на рисунке.
(Б) Запись СПДМ с S1 сегмента рефлекторной дуги Ахиллова рефлекса. Стимулирующий электрод расположен над большеберцовым нервом; записывающий электрод расположен над трехглавой мышцей.
Оба звена рефлекторной дуги образованы большеберцовой ветвью седалищного нерва.
(В) При повышении силы тока происходит непосредственная активация аксонов, иннервирующих мышцу, из-за чего возникают коротколатентные М-волны.
(Г) Обратите внимание, что по мере увеличение интенсивности стимула Н-волна постепенно исчезает (от кривой 1 до кривой 3) вследствие того,
что ортодромные двигательные импульсы из (А) подавляются антидромными импульсами катода, которые представлены в (В) пунктирной линией.
Редактор: Искандер Милевски. Дата публикации: 13.11.2018
Читайте также:
