
Работе нервных центров торможение необходимо для
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
1. ЯВЛЕНИЕ ИЗМЕНЕНИЯ КОЛИЧЕСТВА НЕРВНЫХ ИМПУЛЬСОВ В ЭФФЕРЕНТНЫХ ВОЛОКНАХ РЕФЛЕКТОРНОЙ ДУГИ ПО СРАВНЕНИЮ С АФФЕРЕНТНЫМИ ОБУСЛОВЛЕНО
1) трансформацией ритма в нервном центре
2) наличием доминантного очага возбуждения
3) посттетанической потенциацией
4) рефлекторным последействием
5) наличием общего конечного пути
2. ПОД ТРАНСФОРМАЦИЕЙ РИТМА ВОЗБУЖДЕНИЯ ПОНИМАЮТ
1) направленное распространение возбуждения в ЦНС
2) циркуляцию импульсов в нейронной ловушке
3) увеличение или уменьшение числа импульсов
4) беспорядочное распространение возбуждения в ЦНС
5) увеличение или уменьшение числа синапсов
3. С УВЕЛИЧЕНИЕМ СИЛЫ РАЗДРАЖИТЕЛЯ ВРЕМЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ
4) увеличивается до определенного предела
5) либо увеличивается, либо уменьшается
4. ПРИ УТОМЛЕНИИ ВРЕМЯ РЕФЛЕКСА
4) уменьшается до определенного предела
5) либо увеличивается, либо уменьшается
5. В ОСНОВЕ РЕФЛЕКТОРНОГО ПОСЛЕДЕЙСТВИЯ ЛЕЖИТ
1) пространственная суммация импульсов
2) циркуляция импульсов в нейронной ловушке
3) последовательная суммация импульсов
4) трансформация импульсов
5) замедленное распространение возбуждения по ЦНС
6. ПОД ДИФФУЗНОЙ ИРРАДИАЦИЕЙ ВОЗБУЖДЕНИЯ ПОНИМАЮТ
1) ненаправленное распространение возбуждения по ЦНС
2) изменение ритма возбуждения
3) замедленное распространение возбуждения по ЦНС.
4) направленное распространение возбуждения по ЦНС
5) циркуляцию импульсов в нейронной ловушке
7. ПОВЫШАЮЩАЯ ТРАНСФОРМАЦИЯ РИТМА ВОЗБУЖДЕНИЯ В НЕРВНОЙ СИСТЕМЕ ОБУСЛОВЛЕНА
1) дисперсией возбуждений и низкой лабильностью нервных центров
2) синаптической задержкой
3) утомляемостью нервных центров и дисперсией возбуждений
4) дисперсией и мультипликацией возбуждений
5) дисперсией и высокой чувствительностью нервных центров
1) являются местом возникновения возбуждения в ЦНС
2) формируют потенциал покоя нервной клетки
3) передают возбуждение с нейрона на нейрон
4) проводят токи покоя
5) синтезируют медиаторы
8. В РЕФЛЕКТОРНОЙ ДУГЕ С НАИМЕНЬШЕЙ СКОРОСТЬЮ ВОЗБУЖДЕНИЕ РАСПРОСТРАНЯЕТСЯ ПО ПУТИ
4) обратной афферентации
5) скорость везде одинаковая
9. ЗА ВРЕМЯ РЕФЛЕКСА ПРИНИМАЮТ ВРЕМЯ ОТ НАЧАЛА ДЕЙСТВИЯ РАЗДРАЖИТЕЛЯ ДО
1) конца действия раздражителя
2) появления ответной реакции
3) достижения полезного приспособительного результата
4) возникновения первого потенциала действия
5) все ответы неправильные
10. В ОСНОВЕ ОККЛЮЗИИ ЛЕЖАТ ПРОЦЕССЫ
5) циркуляции импульсов в замкнутых нейронных сетях
11.ВРЕМЯ РЕФЛЕКСА ЗАВИСИТ ПРЕЖДЕ ВСЕГО
1) от силы раздражителя и функционального состояния ЦНС
2) от физических и химических свойств эффектора
3) от физиологических свойств эффектора
4) от иррадиации возбуждения
5) от физических и химических свойств раздражителя
12. ВОЗБУЖДЕНИЕ В НЕРВНОМ ЦЕНТРЕ РАСПРОСТРАНЯЕТСЯ
1) от эфферентных нейронов через промежуточные к афферентным
2) от промежуточных нейронов через эфферентные нейроны к афферентным
3) от афферентных нейронов через промежуточные к эфферентным
4) от промежуточных нейронов через афферентные нейроны к эфферентным
5) все ответы неправильные, т.к. действует закон двустороннего проведения возбуждения
13. РОЛЬ ЗВЕНА ОБРАТНОЙ АФФЕРЕНТАЦИИ ЗАКЛЮЧАЕТСЯ В ОБЕСПЕЧЕНИИ
1) морфологического соединения нервного центра с эффектором
2) распространения возбуждения от афферентного звена к эфферентному
3) оценки результата рефлекторного акта
4) закона двухстороннего распространения возбуждения
5) морфологического соединения эффектора с афферентным нейроном
14. НЕРВНАЯ КЛЕТКА ВЫПОЛНЯЕТ ВСЕ ФУНКЦИИ, КРОМЕ
1) приема информации
2) хранения информации
3) кодирования информации
4) выработки медиатора
5) инактивации медиатора
15. ОСНОВНОЙ ФУНКЦИЕЙ ДЕНДРИТОВ ЯВЛЯЕТСЯ
1) проведение возбуждения к телу нейрона
2) выработка медиатора
3) проведение возбуждения от тела клетки к эффектору
4) инактивация медиатора
5) кодирование и хранение информации
16. В ЕСТЕСТВЕННЫХ УСЛОВИЯХ ПОТЕНЦИАЛ ДЕЙСТВИЯ В НЕЙРОНЕ ВОЗНИКАЕТ
1) в области дендритов
3) в соме нервной клетки
4) в начальном сегменте аксона
5) в терминали аксона
16. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ В ЦНС ОСУЩЕСТВЛЯЕТСЯ ПРЕИМУЩЕСТВЕННО С УЧАСТИЕМ СИНАПСОВ
4) всех перечисленных
5) правильного ответа нет
17. ИНТЕГРАТИВНАЯ ДЕЯТЕЛЬНОСТЬ НЕЙРОНА ЗАКЛЮЧАЕТСЯ В
1) суммации всех постсинаптических потенциалов, возникающих на мембране нейрона
2) связи с другими нейронами посредством отростков
3) посттетанической потенциации
4) генерации потенциала покоя
5) кодировании и хранении информации
18. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ ВОЗНИКАЕТ ПРИ ЛОКАЛЬНОЙ
3) католической депрессии
4) статической поляризации
5) передаче информации
19. ВОЗБУЖДАЮЩИЙ ПОСТСИИАПТИЧЕСКИЙ ПОТЕНЦИАЛ РАЗВИВАЕТСЯ В РЕЗУЛЬТАТЕ ОТКРЫТИЯ НА ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ КАНАЛОВ ДЛЯ ИОНОВ
4) магния
5) водорода
20. ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ - ЭТО ЛОКАЛЬНЫЙ ПРОЦЕСС ДЕПОЛЯРИЗАЦИИ, РАЗВИВАЮЩИЙСЯ НА МЕМБРАНЕ
1) аксонного холмика
21. С БОЛЕЕ ВЫСОКОЙ ЧАСТОТОЙ ГЕНЕРИРУЮТ ИМПУЛЬСЫ ТЕ НЕЙРОНЫ, У КОТОРЫХ СЛЕДОВАЯ ГИПЕРПОЛЯРИЗАЦИЯ ДЛИТСЯ
5) связи между следовой гиперполяризацией и частотой импульсов нет
22. КОМПЛЕКС СТРУКТУР, НЕОБХОДИМЫХ ДЛЯ ОСУЩЕСТВЛЕНИЯ РЕФЛЕКТОРНОЙ РЕАКЦИИ, НАЗЫВАЮТ
1) функциональной системой
2) нервным центром
3) рефлекторной дугой
4) доминантным очагом возбуждения
5) нервно-мышечным препаратом
23. ПРИ ДЛИТЕЛЬНОМ РАЗДРАЖЕНИИ КОЖИ ЛАПКИ ЛЯГУШКИ РЕФЛЕКТОРНОЕ ОТДЕРГИВАНИЕ ЛАПКИ ПРЕКРАЩАЕТСЯ ИЗ-ЗА
РАЗВИТИЯ УТОМЛЕНИЯ
1) в мышцах лапки
2) в нервно-мышечных синапсах
3) в нервном центре рефлекса
4) в звене обратной афферентации
5) в вегетативных ганглиях
24. УВЕЛИЧЕНИЕ ЧИСЛА ВОЗБУЖДЕННЫХ НЕЙРОНОВ В ЦНС ПРИ УСИЛЕНИИ РАЗДРАЖЕНИЯ ПРОИСХОДИТ ВСЛЕДСТВИЕ
4) пространственной суммации
5) временной суммации
25. РАСПРОСТРАНЕНИЕ ВОЗБУЖДЕНИЯ ОТ ОДНОГО АФФЕРЕНТНОГО НЕЙРОНА НА МНОГИЕ ИНТЕРНЕЙРОНЫ НАЗЫВАЕТСЯ ПРОЦЕССОМ
1) трансформации ритма
4) проторения пути
5) пространственной суммации
1) афферентного синтеза
2) последовательной суммации .
5) трансформации ритма
26. УСИЛЕНИЕ РЕФЛЕКТОРНОЙ РЕАКЦИИ НЕ МОЖЕТ ВОЗНИКНУТЬ В РЕЗУЛЬТАТЕ
1) торможения рефлекса-антагониста
2) посттетанической потенциации
3) последовательной суммации
27. ПОСТТЕТАНИЧЕСКАЯ ПОТЕНЦИАЦИЯ ЗАКЛЮЧАЕТСЯ В УСИЛЕНИИ РЕФЛЕКТОРНОЙ РЕАКЦИИ НА РАЗДРАЖЕНИЕ, КОТОРОМУ ПРЕДШЕСТВОВАЛО
1) ритмическое раздражение нервного центра
2) пространственная суммация импульсов
3) понижающая трансформация ритма импульсов
4) торможение нервного центра
5) все перечисленное
28. ПРОСТРАНСТВЕННАЯ СУММАЦИЯ ИМПУЛЬСОВ ОБЕСПЕЧИВАЕТСЯ
1) дивергенцией возбуждения
2) наличием доминантного очага возбуждения
3) конвергенцией возбуждения
4) наличием обратной связи
5) пространственным облегчением
29. ДЛЯ НЕЙРОНОВ ДОМИНАНТНОГО ОЧАГА ХАРАКТЕРНА
1) способность к суммации возбуждений
2) все перечисленное
3) высокая лабильность
5) способность к трансформации ритма
30. НЕРВНЫЕ ЦЕНТРЫ ОБЛАДАЮТ
2) высокой чувствительностью к химическим раздражителям
3) способностью к суммации возбуждений
4) способностью к трансформации ритма
5) всеми перечисленными свойствами
31. УСИЛЕНИЕ РЕФЛЕКТОРНОЙ РЕАКЦИИ ВОЗНИКАЕТ В РЕЗУЛЬТАТЕ
1) торможения рефлекса-антагониста
2) посттетанической потенциации
3) последовательной суммации
5) всех перечисленных процессов
32. УЧАСТИЕ В РАЗЛИЧНЫХ РЕФЛЕКТОРНЫХ РЕАКЦИЯХ ОДНИХ И ТЕХ ЖЕ ЭФФЕРЕНТНЫХ НЕЙРОНОВ И ЭФФЕКТОРОВ ОБУСЛОВЛЕНО НАЛИЧИЕМ
1) пластичности нервных центров
2) полифункциональности нейронов
3) общего конечного пути
4) проторения пути
5) дивергенции возбуждений
33. ПРЕВЫШЕНИЕ ЭФФЕКТА ОДНОВРЕМЕННОГО ДЕЙСТВИЯ ДВУХ СЛАБЫХ АФФЕРЕНТНЫХ ВОЗБУЖДЕНИЙ НАД СУММОЙ ИХ РАЗДЕЛЬНЫХ ЭФФЕКТОВ НАЗЫВАЮТ
34. БОЛЕЕ СЛАБЫЙ ЭФФЕКТ ОДНОВРЕМЕННОГО ДЕЙСТВИЯ ДВУХ СИЛЬНЫХ АФФЕРЕНТНЫХ ВХОДОВ В ЦНС, ЧЕМ СУММА ИХ РАЗДЕЛЬНЫХ ЭФФЕКТОВ, НАЗЫВАЕТСЯ
2) понижающей трансформацией
4) отрицательной индукцией
35. ДЛЯ РАЗВИТИЯ ТОРМОЖЕНИЯ В ЦНС НЕОБХОДИМО ВСЕ КРОМЕ
1) нарушения целостности нервного центра
3) открытия хлорных каналов
4) открытия калиевых каналов
36. МЕДИАТОР ТОРМОЗНОГО НЕЙРОНА, КАК ПРАВИЛО, НА ПОСТ- СИНАПТИЧЕСКОЙ МЕМБРАНЕ ВЫЗЫВАЕТ
1) статическую поляризацию
5) католическую депрессию
37. ВРЕМЯ РЕФЛЕКСА В ОПЫТЕ И.М.СЕЧЕНОВА
1) не изменяется
2) в этом опыте не определяется
5) может увеличиваться, может уменьшаться
38. В ОПЫТЕ И.М.СЕЧЕНОВА РАЗРЕЗ МОЗГА ПРОВОДИТСЯ МЕЖДУ
1) грудными и поясничными отделами спинного мозга
2) между зрительными буграми и вышележащими отделами
3) продолговатым и спинным мозгом
4) на любом уровне ЦНС
5) правильного ответа нет
39. ТОРМОЖЕНИЕ БЫЛО ОТКРЫТО И.М. СЕЧЕНОВЫМ ПРИ РАЗДРАЖЕНИИ
1) спинного мозга
2) зрительных бугров
3) коры головного мозга
5) продолговатого мозга
40. ПРИ РАЗВИТИИ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ МЕМБРАНА НЕЙРОНА НАХОДИТСЯ В СОСТОЯНИИ
1) статической поляризации
3) устойчивой длительной деполяризации
4) католической депрессии
41. ЯВЛЕНИЕ, ПРИ КОТОРОМ ВОЗБУЖДЕНИЕ ОДНОЙ МЫШЦЫ СОПРОВОЖДАЕТСЯ ТОРМОЖЕНИЕМ ЦЕНТРА МЫШЦЫ-АНТАГОНИСТА, НАЗЫВАЕТСЯ
1) отрицательной индукцией
5) реципрокным торможением
42. ТОРМОЖЕНИЕ-ЭТО ПРОЦЕСС
2)распространяющийся, если ТПСП достигает критического уровня
3) всегда распространяющийся
4) может быть локальным, может быть распространяющимся
5) правильного ответа нет
43. К СПЕЦИФИЧЕСКИМ ТОРМОЗНЫМ НЕЙРОНАМ ОТНОСЯТСЯ
1) нейроны черной субстанции и красного ядра среднего
мозга
2) пирамидные клетки коры больших полушарий
3) нейроны ядра Дейтерса продолговатого мозга
4) клетки Пуркинье и Реншоу
5) нейроны солнечного сплетения
44. ЯВЛЕНИЕ СОПРЯЖЕННОГО ТОРМОЖЕНИЯ МОЖНО НАБЛЮДАТЬ
1) в опыте И.М.Сеченова
2) в опыте, когда при развитии одного рефлекса раздражается рецепторное поле антагонистического рефлекса
3) при одновременном раздражении рецепторных полей двух спинальных рефлексов
4) в опыте с односторонним разрушением мозжечка
5) в опыте с введением стрихнина
1) в обеспечении координации работы центров-антагонистов
2) в освобождении ЦНС от переработки несущественной информации
3) в выполнении защитной функции
4) в проторении пути
5) в формировании общего конечного пути
46. ТПСП ВОЗНИКАЕТ ВСЛЕДСТВИЕ ИЗМЕНЕНИЯ ПРОНИЦАЕМОСТИ МЕМБРАНЫ ДЛЯ ИОНОВ
2) натрия и хлора
3) калия и хлора
47. ВОЗНИКНОВЕНИЕ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ ВЕРОЯТНО
1) при низкой частоте импульсов
2) при секреции тормозных медиаторов
3) при возбуждении вставочных тормозных нейронов
4) при увеличении частоты импульсов
5) при блокаде тормозных каналов
48. ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ ОСУЩЕСТВЛЯЕТСЯ ПОСРЕДСТВОМ СИНАПСОВ
49. МЕХАНИЗМ ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ СВЯЗАН СО СЛЕДУЮЩИМ ЯВЛЕНИЕМ
1) длительной деполяризацией
2) работой К - Na-насоса
3) работой Са-насоса
50. С ТОЧКИ ЗРЕНИЯ БИНАРНО-ХИМИЧЕСКОЙ ТЕОРИИ ПРОЦЕСС ТОРМОЖЕНИЯ ВОЗНИКАЕТ
1) вследствие инактивации холинэстеразы
2) при функционировании специальных тормозных нейронов, вырабатывающих специальные медиаторы
3) в тех же структурах и с помощью тех же медиаторов,
что и процесс возбуждения
4) при уменьшении синтеза возбуждающего медиатора
5) при блокаде тормозных синапсов
51.. С ТОЧКИ ЗРЕНИЯ УНИТАРНО-ХИМИЧЕСКОЙ ТЕОРИИ ТОРМОЖЕНИЕ ВОЗНИКАЕТ
1) вследствие инактивации холинэстеразы
2) при уменьшении синтеза возбуждающего медиатора
3) в тех же структурах и с помощью тех же медиаторов,
что и процесс возбуждения
4) при функционировании специальных тормозных нейронов, вырабатывающих специальные медиаторы
5) при блокаде тормозных синапсов
52. ЯВЛЕНИЕ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ БЫЛО ОТКРЫТО
1) Ч. Шеррингтоном
4) братьями Вебер
5) Н.Е. Введенским
53. ЯВЛЕНИЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ БЫЛО ОТКРЫТО
1) братьями Вебер
54. ТОРМОЖЕНИЕ - ЭТО ПРОЦЕСС
1) возникающий в результате утомления нервных клеток
2) приводящий к снижению КУД нервной клетки
3) возникающий в рецепторах при чрезмерно сильных раздражителях
4) препятствующий возникновению возбуждения или ослабляющий уже возникшее возбуждение
5) возникающий в рецепторах при чрезмерно длительном раздражении
55. В РАБОТЕ НЕРВНЫХ ЦЕНТРОВ ТОРМОЖЕНИЕ НЕОБХОДИМО ДЛЯ
1) замыкания дуги рефлексов в ответ на раздражение
2) обеспечения сохранности, регуляции и координации функций
3) объединения клеток ЦНС в нервные центры
4) защиты нейронов от чрезмерного возбуждения
5) всего перечисленного
56.ДИФФУЗНАЯ ИРРАДИАЦИЯ МОЖЕТ БЫТЬ ПРЕКРАЩЕНА В РЕЗУЛЬТАТЕ
1) латерального торможения
2) увеличения силы раздражителя
3) введения стрихнина
4) реципрокного торможения
5) всего перечисленного
57. О РАЗВИТИИ ТОРМОЖЕНИЯ В ОПЫТЕ И.М.СЕЧЕНОВА НА ЛЯГУШКЕ СУДЯТ ПО
1) появлению судорожных сокращений лапок
2) изменению времени спинального рефлекса
3) снижению частоты сердцебиений с последующей остановкой сердца
4) вовлечению в рефлекторный ответ мышц передних
конечностей
5) появлению манежных движений
58. СОКРАЩЕНИЕ МЫШЦ-СГИБАТЕЛЕЙ ПРИ ОДНОВРЕМЕННОМ
РАССЛАБЛЕНИИ МЫШЦ-РАЗГИБАТЕЛЕЙ ВОЗМОЖНО В РЕЗУЛЬТАТЕ
1) активного отдыха
2) облегчения распространения возбуждения
3) реципрокного торможения
4) пессимального торможения
5) отрицательной идукции
59. ТОРМОЖЕНИЕ НЕЙРОНОВ СОБСТВЕННЫМИ ИМПУЛЬСАМИ, ПОСТУПАЮЩИМИ ПО КОЛЛАТЕРАЛЯМ АКСОНА К ТОРМОЗНЫМ КЛЕТКАМ, НАЗЫВАЮТ
60.. С ПОМОЩЬЮ ТОРМОЗНЫХ ВСТАВОЧНЫХ КЛЕТОК РЕНШОУ
ОСУЩЕСТВЛЯЕТСЯ ТОРМОЖЕНИЕ
61.. ТОРМОЖЕНИЕ МОТОНЕЙРОНОВ МЫШЦ-АНТАГОНИСТОВ
ПРИ СГИБАНИИ И РАЗГИБАНИИ КОНЕЧНОСТЕЙ НАЗЫВАЮТ
5) правильного ответа нет
244. ПРИ СГИБАНИИ КОНЕЧНОСТИ ВСТАВОЧНЫЕ ТОРМОЗНЫЕ НЕЙРОНЫ ЦЕНТРА МЫШЦ-РАЗГИБАТЕЛЕЙ ДОЛЖНЫ БЫТЬ
3) в состоянии покоя
4) поочередно в каждом из перечисленных состояний
5) состояние не установлено
62.. ТОРМОЗНОЙ ЭФФЕКТ СИНАПСА, РАСПОЛОЖЕННОГО ВБЛИЗИ АКСОННОГО ХОЛМИКА, ПО СРАВНЕНИЮ С ДРУГИМИ УЧАСТКАМИ НЕЙРОНА, БОЛЕЕ
4) сильный и продолжительный
5) слабый, но продолжительный
63.. РАЗВИТИЮ ТОРМОЖЕНИЯ НЕЙРОНОВ СПОСОБСТВУЕТ
1) гиперполяризация мембраны аксонного холмика
2) деполяризация сомы и дендритов
3) деполяризация мембраны аксонного холмика и начального сегмента
4) реполяризация сомы и дендритов
5) реполяризация аксонных холмиков
64.. ПО СВОЕМУ МЕХАНИЗМУ ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ МОЖЕТ БЫТЬ
1) и де-, и гиперполяризационным
2) только гиперполяризационным
3) только деполяризационным
4) только реполяризационным
5) и де-, и реполяризационным
65.. ПО СВОЕМУ МЕХАНИЗМУ ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ МОЖЕТ БЫТЬ
Выберите один правильный ответ.
212. ДЛЯ РАЗВИТИЯ ТОРМОЖЕНИЯ В ЦНС НЕОБХОДИМО ВСЕ, КРОМЕ
3) открытия хлорных каналов
4) открытия калиевых каналов
5) нарушения целостности нервного центра
213. МЕДИАТОР ТОРМОЗНОГО НЕЙРОНА, КАК ПРАВИЛО, НА ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ ВЫЗЫВАЕТ
1) статическую поляризацию
3) гиперполяризацию
214. ВРЕМЯ РЕФЛЕКСА В ОПЫТЕ СЕЧЕНОВА
1) не изменяется
2) в этом опыте не определяется
4) увеличивается
215. В ОПЫТЕ СЕЧЕНОВА РАЗРЕЗ МОЗГА ПРОВОДИТСЯ МЕЖДУ
1) грудными и поясничными отделами спинного мозга
2) продолговатым и спинным мозгом
3) между зрительными буграми и вышележащими отделами
216. ТОРМОЖЕНИЕ БЫЛО ОТКРЫТО СЕЧЕНОВЫМ ПРИ РАЗДРАЖЕНИИ
1) спинного мозга
2) продолговатого мозга
3) коры головного мозга
5) зрительных бугров
217. ПРИ РАЗВИТИИ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ МЕМБРАНА НЕЙРОНА НАХОДИТСЯ В СОСТОЯНИИ
1) статической поляризации
3) устойчивой длительной деполяризации
218. ЯВЛЕНИЕ, ПРИ КОТОРОМ ВОЗБУЖДЕНИЕ ОДНОЙ МЫШЦЫ СОПРОВОЖДАЕТСЯ ТОРМОЖЕНИЕМ ЦЕНТРА МЫШЦЫ-АНТАГОНИСТА, НАЗЫВАЕТСЯ
1) отрицательной индукцией
5) реципрокным торможением
219. ТОРМОЖЕНИЕ - ЭТО ПРОЦЕСС
1) всегда распространяющийся
2) распространяющийся, если ТПСП достигает критического уровня
3) локальный
220. К СПЕЦИФИЧЕСКИМ ТОРМОЗНЫМ НЕЙРОНАМ ОТНОСЯТСЯ
1) нейроны черной субстанции и красного ядра среднего мозга
2) пирамидные клетки коры больших полушарий
3) нейроны ядра Дейтерса продолговатого мозга
4) клетки Пуркинье и Реншоу
221. ЯВЛЕНИЕ СОПРЯЖЕННОГО ТОРМОЖЕНИЯ МОЖНО НАБЛЮДАТЬ
1) в опыте Сеченова
2) при одновременном раздражении рецептивных полей двух спинальных рефлексов
3) в опыте, когда при развитии одного рефлекса раздражается рецептивное поле антагонистического рефлекса
222. ЗНАЧЕНИЕ РЕЦИПРОКНОГО ТОРМОЖЕНИЯ ЗАКЛЮЧАЕТСЯ
1) в выполнении защитной функции
2) в освобождении ЦНС от переработки несущественной информации
3) в обеспечении координации работы центров-антагонистов
223. ТПСП ВОЗНИКАЕТ ВСЛЕДСТВИЕ ИЗМЕНЕНИЯ ПРОНИЦАЕМОСТИМЕМБРАНЫ ДЛЯ ИОНОВ
2) натрия и хлора
3) калия и хлора
224. ВОЗНИКНОВЕНИЕ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ ВЕРОЯТНО
1) при низкой частоте импульсов
2) при секреции тормозных медиаторов
3) при возбуждении вставочных тормозных нейронов
4) при увеличении частоты импульсов
225. ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ ОСУЩЕСТВЛЯЕТСЯ ПОСРЕДСТВОМ СИНАПСОВ
4) аксо-аксональных
226. МЕХАНИЗМ ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ СВЯЗАН
1) с гиперполяризацией
2) с работой К - Nа насоса
3) с работой Са насоса
4) с длительной деполяризацией
227. С ТОЧКИ ЗРЕНИЯ БИНАРНО-ХИМИЧЕСКОЙ ТЕОРИИ ПРОЦЕСС ТОРМОЖЕНИЯ ВОЗНИКАЕТ
1) вследствие инактивации холинэстеразы
2) при уменьшении синтеза возбуждающего медиатора
3) в тех же структурах и с помощью тех же медиаторов, что и процесс возбуждения
4) при функционировании специальных тормозных нейронов,вырабатывающих специальные медиаторы
228. С ТОЧКИ ЗРЕНИЯ УНИТАРНО-ХИМИЧЕСКОЙ ТЕОРИИ ТОРМОЖЕНИЕ ВОЗНИКАЕТ
1) вследствие инактивации холинэстеразы
2) при уменьшении синтеза возбуждающего медиатора
3) при функционировании специальных тормозных нейронов,вырабатывающих специальные медиаторы
4) в тех же структурах и с помощью тех же медиаторов, что и процесс возбуждения
229. ЯВЛЕНИЕ ПЕССИМАЛЬНОГО ТОРМОЖЕНИЯ БЫЛО ОТКРЫТО
1) Ч. Шеррингтоном
4) братьями Вебер
5) Н.Е. Введенским
230. ЯВЛЕНИЕ ЦЕНТРАЛЬНОГО ТОРМОЖЕНИЯ БЫЛО ОТКРЫТО
1) братьями Вебер
2) Ч. Шеррингтоном
231. ТОРМОЖЕНИЕ - ЭТО ПРОЦЕСС
1) возникающий в результате утомления нервных клеток
2) приводящий к снижению КУД нервной клетки
3) возникающий в рецепторах при чрезмерно сильных раздражителях
4) препятствующий возникновению возбуждения или ослабляющийуже возникшее возбуждение
232. В РАБОТЕ НЕРВНЫХ ЦЕНТРОВ ТОРМОЖЕНИЕ НЕОБХОДИМО
1) для замыкания дуги рефлексов в ответ на раздражение
2) для защиты нейронов от чрезмерного возбуждения
3) для объединения клеток ЦНС в нервные центры
4) для обеспечения сохранности, регуляции и координации функций
233. ДИФФУЗНАЯ ИРРАДИАЦИЯ МОЖЕТ БЫТЬ ПРЕКРАЩЕНА В РЕЗУЛЬТАТЕ
1) введения стрихнина
2) увеличения силы раздражителя
3) латерального торможения
234. О РАЗВИТИИ ТОРМОЖЕНИЯ В ОПЫТЕ СЕЧЕНОВА НА ЛЯГУШКЕ СУДЯТ ПО
1) появлению судорожных сокращений лапок
2) урежению сердцебиений с последующей остановкой сердца
3) изменению времени спинального рефлекса
235. СОКРАЩЕНИЕ МЫШЦ-СГИБАТЕЛЕЙ ПРИ ОДНОВРЕМЕННОМ РАССЛАБЛЕНИИ МЫШЦ-РАЗГИБАТЕЛЕЙ ВОЗМОЖНО В РЕЗУЛЬТАТЕ
1) активного отдыха
3) отрицательной идукции
4) пессимального торможения
5) реципрокного торможения
236. ТОРМОЖЕНИЕ НЕЙРОНОВ СОБСТВЕННЫМИ ИМПУЛЬСАМИ, ПОСТУПАЮЩИМИ ПО КОЛЛАТЕРАЛЯМ АКСОНА
К ТОРМОЗНЫМ КЛЕТКАМ, НАЗЫВАЮТ
5) возвратным
237. С ПОМОЩЬЮ ТОРМОЗНЫХ ВСТАВОЧНЫХ КЛЕТОК РЕНШОУ ОСУЩЕСТВЛЯЕТСЯ ТОРМОЖЕНИЕ
4) возвратное
238. ТОРМОЖЕНИЕ МОТОНЕЙРОНОВ МЫШЦ-АНТАГОНИСТОВ ПРИ СГИБАНИИ И РАЗГИБАНИИ КОНЕЧНОСТЕЙ НАЗЫВАЮТ
4) реципрокным
239. ПРИ СГИБАНИИ КОНЕЧНОСТИ ВСТАВОЧНЫЕ ТОРМОЗНЫЕ НЕЙРОНЫ ЦЕНТРА МЫШЦ-РАЗГИБАТЕЛЕЙ ДОЛЖНЫ БЫТЬ
1) в состоянии покоя
3) возбуждены
240. ТОРМОЗНОЙ ЭФФЕКТ СИНАПСА, РАСПОЛОЖЕННОГО ВБЛИЗИ АКСОННОГО ХОЛМИКА,
ПО СРАВНЕНИЮ С ДРУГИМИ УЧАСТКАМИ НЕЙРОНА БОЛЕЕ
2) сильный
241. РАЗВИТИЮ ТОРМОЖЕНИЯ НЕЙРОНОВ СПОСОБСТВУЕТ
1) деполяризация мембраны аксонного холмика и начального сегмента
2) деполяризация сомы и дендритов
3) гиперполяризация мембраны аксонного холмика
242. ПО СВОЕМУ МЕХАНИЗМУ ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ МОЖЕТ БЫТЬ
1) только деполяризационным
2) только гиперполяризационным
3) и де- , и гиперполяризационным
243. ПО СВОЕМУ МЕХАНИЗМУ ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ МОЖЕТ БЫТЬ
1) и де- , и гиперполяризационным
2) только гиперполяризационным
3) только деполяризационным
ПРИ ТОРМОЖЕНИИ. НА СУБСИНАПТИЧЕСКОЙ МЕМБРАНЕВОЗНИКАЕТ
А.2 Пресинаптическом 1. Кратковременная деполяризация.
Б.3 Постсинаптическом 2. Длительная деполяризация.
3. Гиперполяризация или длительная деполяризация.
ТЕОРИИ ТОРМОЖЕНИЯ. ЗАКЛЮЧАЮТСЯ В ТОМ, ЧТО
А.3 Унитарно-химическая 1. Торможение является следствием утомления.
Б.2 Бинарно-химическая 2. Торможение возникает в результате функционирования тормозных нейронов.
3. Торможение проявляется в тех же структурах и с помощью тех же медиаторов, что и возбуждение.
НЕРВНЫЙ ПРОЦЕСС. ХАРАКТЕРИЗУЮТ ПРИЗНАКИ
А.2 Возбуждение 1. Всегда локальный процесс, проявляющийся
Б.1 Торможение в длительной устойчивой деполяризацииили гиперполяризации мембраны нейрона.
2. Местный или распространяющийся процесс, обусловленный открытием натриевых каналов.
ЯВЛЕНИЕ. РАЗВИВАЕТСЯ ВСЛЕДСТВИЕ
А.4 Пессимального 1. Длительного действия постоянного тока
торможения в области приложения катода.
Б.1 Катодической 2. Кратковременного действия постоянного токав области приложения катода.
депрессии 3. Раздражения блуждающего нерва.
4. Увеличении частоты импульсации.
5. Одновременного раздражения рецептивных полей двух спинальных рефлексов.
ИССЛЕДОВАТЕЛИ. ФИЗИОЛОГИИ ЦНС ВНЕСЛИ СЛЕДУЮЩИЙ ВКЛАД В РАЗВИТИЕ
А.2 А.А.Ухтомский 1. Сформулировал принципы общего
Б.3 Бергер конечного пути и реципрокности.
В.1 Ч.Шеррингтон 2. Разработал учение о доминанте.
3. Впервые заригистрировал ЭЭГ у человека.
А.2 Является 1. Исчезновения коленного рефлексапри травме поясничного отдела позвоночника.
Б.1 Не является 2. Прекращения слюноотделения в процессе приема пищи при появлении сильной боли в животе.
ВИД ТОРМОЖЕНИЯ. ВЫПОЛНЯЕТ ФУНКЦИЮ
А.2 Латеральное 1. Подавляет возбуждение центра
Б.4 Возвратное антагонистической функции.
В.1 Реципрокное 2. Устраняет диффузную иррадиацию возбуждения.
3. Прекращает выход медиатора в синаптическую щель.
4. Ослабляет возбуждение мотонейронов ихсобственными импульсами через клетки Реншоу.
ВИДЫ НЕЙРОНОВ. ПРЕДСТАВЛЯЮТ СОБОЙ
А.3 Альфа-мотонейрон 1. Нейрон моторной зоны коры большого мозга.
Б.2 Гамма-мотонейрон 2. Нейрон передних рогов спинного мозга,
В.1 Гигантская пира- иннервирующий интрафузальные волокна мидальная клетка скелетных мышц.
Беца 3. Нейрон передних рогов спинного мозга,
Г.5 Клетка Реншоу иннервирующий экстрафузальные волокна скелетных мышц.
4. Тормозный нейрон коры мозжечка.
5. Тормозный интернейрон спинного мозга.
ВИДЫ ПОСТСИНАПТИЧЕСКИХ ПОТЕНЦИАЛОВ НЕЙРОНА. ОБУСЛОВЛЕНЫ ОТКРЫТИЕМ КАНАЛОВ ДЛЯ ИОНОВ
А.1 ВПСП 1. Натрия.
Б.23 ТПСП 2. Калия.
ПРИ АКТИВАЦИИ ХЛОРНЫХ КАНАЛОВ. НАБЛЮДАЕТСЯ ТОК ИОНОВ ХЛОРА .
А.1 Пресинаптических 1. Наружу из клетки.
Б.2 Постсинаптических 2. Из внешней среды в клетку.
Определите верны или неверны утверждения и связь между ними.
254. Торможение спинального рефлекса в опыте Сеченова вызывают раздражением зрительных бугров кристалликом хлористого натрия, потому что ионы натрия и хлора вызывают гиперполяризацию нейронов.
5) ВНН
255. Пресинаптическое торможение очень эффективно при обработке поступающей к нейрону информации, потому что при пресинаптическом торможении возбуждение может быть подавлено избирательно на одном синаптическом входе, не влияя на другие синаптические входы.
5) ВВВ
256. Для демонстрации роли торможения лягушке вводят стрихнин, потому что стрихнин активирует тормозные синапсы.
5) ВНН
257. Для демонстрации роли торможения лягушке вводят стрихнин, потому что стрихнин блокирует тормозные синапсы.
5) ВВВ
258. Для демонстрации роли торможения лягушке вводят стрихнин, потому что после введения стрихнина у лягушки наблюдается
диффузная иррадиация возбуждения.
5) ВВВ
259. Нейрон может находиться в состоянии покоя, возбуждения или торможения, потому что на одном нейроне могут суммироваться
либо возбуждающие, либо тормозные постсинаптические потенциалы.
5) ВНН
260. На одном нейроне могут суммироваться только ВПСП или только ТПСП,потому что согласно принципу Дейла, один нейрон использует
во всех своих терминалях только один вид медиатора.
5) НВН
261. По аксону нейрона может распространяться либо возбуждение, либо торможение, потому что при суммации ВПСП
и ТПСП суммарный потенциал может быть либо положительным, либо отрицательным.
5) НВН
262. Опыт Сеченова проводится на спинальной лягушке, потому что в опыте Сеченова измеряют время спинального рефлекса.
5) НВН
263. Опыт Сеченова проводится на таламической лягушке, потому что для проявления спинального рефлекса в опыте Сеченова необходимо положить на зрительные бугры кристаллик соли.
Регуляция нервной деятельности представляет собой процессы возбуждения и торможения в ЦНС. Вначале она возникает как элементарная реакция на раздражение. В процессе эволюции произошло усложнение нейрогуморальных функций, приводящее к образованию основных отделов нервной и эндокринной систем. В данной статье мы изучим один из главных процессов – торможение в ЦНС, виды и механизмы его осуществления.
Нервная ткань, её строение и функции
Одна из разновидностей животных тканей, названная нервной, имеет особое строение, обеспечивающее как процесс возбуждения, так и приводящее в действие функции торможения в ЦНС. Нервные клетки состоят из тела и отростков: коротких (дендритов) и длинного (аксона), который обеспечивает передачу нервных импульсов от одного нейроцита к другому. Окончание аксона нервной клетки контактирует с дендритами следующего нейроцита в местах, называемых синапсами. Они обеспечивают передачу биоэлектрических импульсов по нервной ткани. Причем возбуждение всегда движется в одном направлении – с аксона на тело или дендриты другого нейроцита.
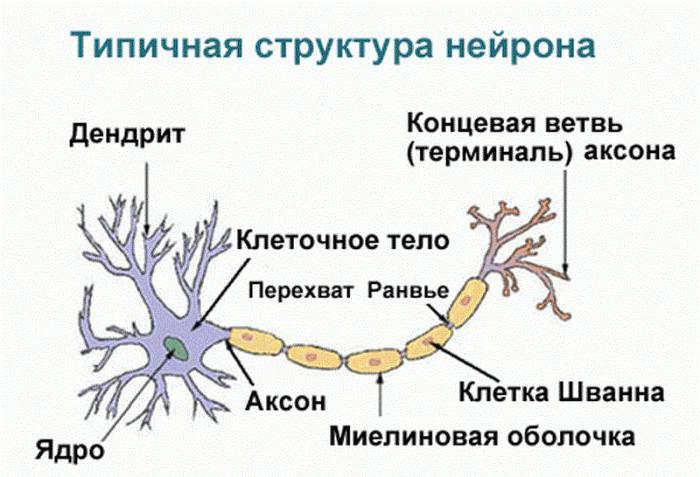
Еще одно свойство, кроме возбуждения, протекающее в нервной ткани, – торможение в ЦНС. Оно является ответной реакцией организма на действие раздражителя, ведущей к снижению или полному прекращению двигательной или секреторной активности, в которой участвуют центробежные нейроны. Торможение в нервной ткани может возникать и без предварительного возбуждения, а только лишь под воздействием тормозного медиатора, например ГАМК. Он является одним из главных трансмиттеров торможения. Здесь же можно назвать такое вещество, как глицин. Эта аминокислота участвует в усилении тормозных процессов и стимулирует в синапсах выработку молекул гаммааминомаслянной кислоты.
И. М. Сеченов и его работы в нейрофизиологии
Выдающийся российский ученый, создатель теории рефлекторной деятельности головного мозга доказал наличие в центральных отделах нервной системы особых комплексов клеток, способных к инактивации биоэлектрических процессов. Открытие центров торможения в ЦНС стало возможным благодаря применению И. Сеченовым трех видов экспериментов. К ним относятся: перерезание участков коры в различных зонах головного мозга, стимуляция отдельных локусов серого вещества физическими или химическими факторами (электрическим током, раствором хлорида натрия), а также метод физиологического возбуждения мозговых центров. И. М. Сеченов был прекрасным экспериментатором, проводя сверхточные разрезы в зоне между зрительными буграми и непосредственно в самом таламусе лягушки. Он наблюдал уменьшение и полное прекращение двигательной активности конечностей животного.
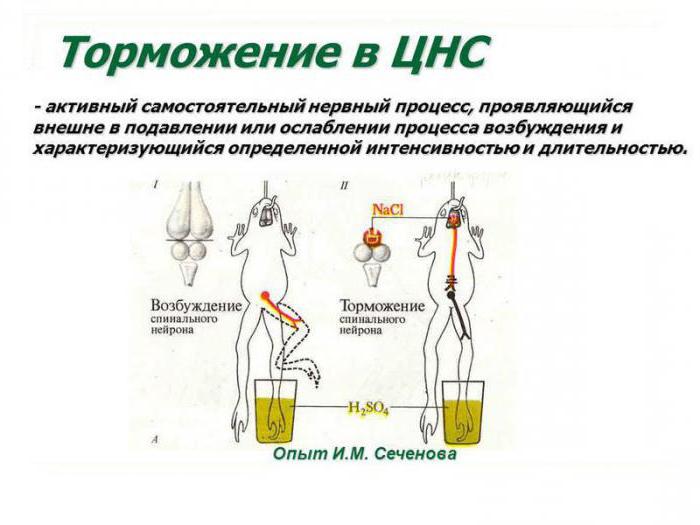
Взаимодействуют ли нервные процессы?
Исследования выдающихся российских физиологов И. П. Павлова и И. М. Сеченова доказали, что работа центральной нервной системы характеризуется координацией рефлекторных реакций организма. Взаимодействие процессов возбуждения и торможения в ЦНС приводит к согласованной регуляции функций организма: двигательной активности, дыхания, пищеварения, выделения. Биоэлектрические процессы одновременно происходят в нервных центрах и могут последовательно меняться во времени. Это обеспечивает корреляцию и своевременное прохождение ответных рефлексов на сигналы внутренней и внешней среды. Многочисленные опыты, проведенные нейрофизиологами, подтвердили тот факт, что возбуждение и торможение в ЦНС – это ключевые нервные явления, в основе которых лежат некоторые закономерности. Остановимся на них подробнее.
Нервные центры коры головного мозга способны распространять оба вида процессов по всей нервной системе. Это свойство называется иррадиацией возбуждения или торможения. Противоположное явление – уменьшение или ограничение участка мозга, распространяющего биоимпульсы. Оно названо концентрацией. Оба вида взаимодействий ученые наблюдают в течение образования условных двигательных рефлексов. Во время начальной стадии формирования двигательных навыков, вследствие иррадиации возбуждения одновременно сокращаются сразу несколько групп мышц, не обязательно участвующих в выполнении формируемого двигательного акта. Только после многократных повторений формируемого комплекса физических движений (катания на коньках, лыжах, велосипеде), в результате концентрации процессов возбуждения в конкретных нервных очагах коры, все движения человека становятся высококоординированными.

Переключения в работе нервных центров могут происходить также вследствие индукции. Она проявляется при выполнении следующего условия: сначала происходит концентрация торможения или возбуждения, причем эти процессы должны быть достаточной силы. В науке известны два вида индукции: S-фаза (центральное торможение в ЦНС усиливает возбуждение) и отрицательная форма (возбуждение вызывает процесс торможения). Встречается также последовательная индукция. В этом случае нервный процесс меняется на противоположный в самом нервном центре. Исследования нейрофизиологов доказали тот факт, что поведение высших млекопитающих и человека определяется явлениями индукции, иррадиации и концентрации нервных процессов возбуждения и торможения.
Безусловное торможение
Рассмотрим более подробно виды торможения в ЦНС и остановимся на такой его форме, которая присуща как животным, так и человеку. Сам термин был предложен И. Павловым. Ученый считал этот процесс одним из врождённых свойств нервной системы и выделил два его вида: гаснущее и постоянное. Остановимся на них детальнее.
Допустим, в коре существует очаг возбуждения, генерирующий импульсы к рабочему органу (к мышцам, секреторным клеткам желез). Вследствие изменения условий внешней или внутренней среды возникает еще один возбужденный участок коры головного мозга. Он вырабатывает биоэлектрические сигналы большей интенсивности, что тормозит возбуждение в ранее активном нервном центре и его рефлекторной дуге. Гаснущее торможение в ЦНС приводит к тому, что интенсивность ориентировочного рефлекса постепенно уменьшается. Объяснение этому следующее: первичный раздражитель уже не вызывает процесса возбуждения в рецепторах афферентного нейрона.
Другой вид торможения, наблюдаемого как у человека, так и у животных, демонстрирует опыт, проведенный лауреатом нобелевской премии в 1904 году И. П. Павловым. Во время кормления собаки (с выведенной из щеки фистулой) экспериментаторы включали резкий звуковой сигнал – выделение слюны из фистулы прекращалось. Такой вид торможения ученый назвал запредельным.
Являясь врождённым свойством, торможение в ЦНС протекает по безусловно-рефлекторному механизму. Оно достаточно пассивно и не вызывает расхода большого количества энергии, приводя к прекращению условных рефлексов. Постоянное безусловное торможение сопровождает многие психосоматические заболевания: дискинезии, спастический и вялый параличи.
Что такое гаснущий тормоз
Продолжая изучать механизмы торможения в ЦНС, рассмотрим, что представляет собой один из его видов, названный гаснущим тормозом. Хорошо известно, что ориентировочный рефлекс представляет собой реакцию организма на воздействие нового постороннего сигнала. В этом случае в коре мозга образуется нервный центр, находящийся в состоянии возбуждения. Он и формирует рефлекторную дугу, отвечающую за реакцию организма и называемую ориентировочным рефлексом. Этот рефлекторный акт вызывает торможение условного рефлекса, происходящего в данный момент. После многоразового повторения постороннего раздражителя рефлекс, называемый ориентировочным, постепенно снижается и наконец исчезает. А значит, не вызывает больше торможения условного рефлекса. Такой сигнал и получил название гаснущего тормоза.
Таким образом, внешнее торможение условных рефлексов связано с влиянием на организм постороннего сигнала и является врождённым свойством центральной и периферической нервной системы. Внезапный или новый раздражитель, например, болевое ощущение, посторонний звук, изменение освещенности, не только вызывает ориентировочный рефлекс, но также способствует ослаблению или даже полному прекращению условно-рефлекторной дуги, активной в данный момент. Если посторонний сигнал (кроме болевого) действует повторно, торможение условного рефлекса проявляется меньше. Биологическая роль безусловной формы нервного процесса заключается в проведении ответной реакции организма на раздражитель, наиболее важный в данный момент.
Внутреннее торможение
Его другое название, используемое в физиологии высшей нервной деятельности, – условное торможение. Главная предпосылка возникновения такого процесса – отсутствие подкрепления сигналов, поступающих из внешнего мира, врождёнными рефлексами: пищеварительным, слюноотделительным. Возникшие в этих условиях процессы торможения в ЦНС требуют определенного временного интервала. Рассмотрим их виды более подробно.
Например, дифференцировочное торможение возникает как ответ на сигналы окружающей среды, совпадающие по амплитуде, интенсивности и силе к условному раздражителю. Эта форма взаимодействия нервной системы и окружающего мира позволяет организму более тонко различать раздражители и вычленять из их совокупности тот, который получает подкрепление врожденным рефлексом. Например, на звук звонка с силой 15 Гц, подкрепленный кормушкой с пищей, у собаки выработали условную слюноотделительную реакцию. Если к животному применить еще один звуковой сигнал, силой 25 Гц, не подкрепляя его пищей, в первой серии опытов у собаки из фистулы слюна будет выделяться на оба условных раздражителя. Через некоторое время у животного произойдет дифференциация этих сигналов, и на звук, силой 25 Гц слюна из фистулы перестанет выделяться, то есть разовьется дифференцировочное торможение.
Освободить мозг от информации, потерявшей жизненно значимую роль для организма, – эту функцию как раз и выполняет торможение в ЦНС. Физиология опытным путем доказала, что условные двигательные реакции, хорошо закрепленные выработанными навыками, могут сохраняться на протяжении всей жизни человека, например, катание на коньках, езда на велосипеде.
Подводя итог, можно сказать, что процессы торможения в ЦНС – это ослабление или прекращение определенных реакций организма. Они имеют очень большое значение, так как все рефлексы организма корригируются в соответствии с измененными условиями, а если условный сигнал потерял свое значение, то даже полностью могут исчезать. Различные виды торможения в ЦНС являются базовыми для таких способностей психики человека, как сохранение самообладания, различение раздражителей, ожидание.
Запаздывающий вид нервного процесса
Опытным путем можно создать ситуацию, при которой ответ организма на условный сигнал из внешней среды проявляется еще до воздействия безусловного раздражителя, например пищи. При увеличении промежутка времени между началом воздействия условного сигнала (свет, звук, например, удары метронома) и моментом подкрепления до трех минут выделение слюны на вышеназванные условные раздражители все более запаздывает и проявляется только в момент, когда перед животным появляется кормушка с пищей. Отставание ответа на условный сигнал характеризует процессы торможения в ЦНС, названные запаздывающим видом, при котором его время протекания соответствует интервалу запаздывания безусловного раздражителя, например пищи.
Значение торможения в ЦНС
Биологические основы поведения человека
Оба вида активности нервной ткани, как возбуждение, так и торможение в ЦНС, являются основой высшей нервной деятельности. Она обуславливает физиологические механизмы психической деятельности человека. Учение высшей нервной деятельности было сформулировано И. П. Павловым. Современная его трактовка звучит следующим образом:
- Возбуждение и торможение в ЦНС, происходящие во взаимодействии, обеспечивают сложные психические процессы: память, мышление, речь, сознание, а также формируют сложные поведенческие реакции человека.
Чтобы составить научно обоснованный режим учебы, труда, отдыха, ученые применяют знания закономерностей высшей нервной деятельности.
Биологическое значение такого активного нервного процесса, как торможение, можно определить следующим образом. Изменение условий внешней и внутренней среды (отсутствие подкрепления условного сигнала врождённым рефлексом) влечет за собой адекватные изменения приспособительных механизмов в организме человека. Поэтому приобретенный рефлекторный акт угнетается (гаснет) или вовсе исчезает, так как становится для организма нецелесообразным.
Что такое сон?
И. П. Павлов в своих работах экспериментально доказал тот факт, что процессы торможения в ЦНС и сон имеют единую природу. В период бодрствования организма на фоне общей активности коры головного мозга все же диагностируются отдельные её участки, охваченные внутренним торможением. Во время сна оно иррадиирует по всей поверхности больших полушарий, достигая подкорковых образований: зрительных бугров (таламуса), гипоталамуса, ретикулярной формации и лимбической системы. Как указывал выдающийся нейрофизиолог П. К. Анохин, все вышеперечисленные части центральной нервной системы, ответственные за поведенческую сферу, эмоции и инстинкты, во время сна снижают свою активность. Это влечет за собой снижение генерирования нервных импульсов, поступающих из-под корки. Таким образом, активизация коры снижается. Это обеспечивает возможность покоя и восстановления обмена веществ как в нейроцитах большого мозга, так и во всем организме в целом.

Опытами других ученых (Гесса, Экономо) были установлены особые комплексы нервных клеток, входящие в неспецифические ядра зрительных бугров. Процессы возбуждения, диагностируемые в них, вызывают снижение частоты биоритмов коры, которые можно расценивать как переход от активного состояния (бодрствования) ко сну. Исследования таких участков головного мозга, как Сильвиев водопровод и ІІІ желудочек, подтолкнули ученых к идее о наличии центра регуляции сна. Он анатомически связан с участком мозга, ответственным за бодрствование. Поражение этого локуса коры вследствие травмы или в результате наследственных нарушений у человека приводит к патологическим состояниям бессонницы. Также отметим тот факт, что регуляция такого жизненно важного для организма процесса торможения, как сон, осуществляется нервными центрами промежуточного мозга и подкорковых ядер: хвостатого, миндалевидного, ограды и чечевицеобразного.
Читайте также:
