
Синапс понятие о нервном центре
Нервный центр – это совокупность нейронов, обеспечивающих регуляцию какого-либо конкретного физиологического процесса или функции.
Нервный центр в узком смысле – это совокупность нейронов, без которых данная конкретная функция не может регулироваться. Например, без нейронов дыхательного центра продолговатого мозга дыхание прекращается. Нервный центр в широком смысле — это совокупность нейронов, которые участвуют в регуляции конкретной физиологической функции, но не являются строго обязательными для ее осуществления! Например, в регуляции дыхания кроме нейронов продолговатого мозга участвуют нейроны пневмотаксического центра варолиевого моста, отдельные ядра гипоталамуса, кора больших полушарий и другие образования головного мозга.
Все нейроны нервного центра разделяют на 2 неравные по количеству и качеству группы.
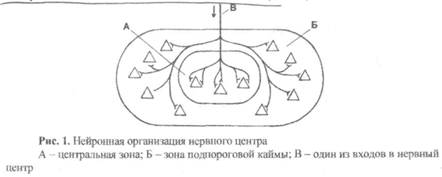
Первая группа – нейроны центральной зоны. Это наиболее возбудимые нейроны, которые возбуждаются в ответ на поступление порогового (для нервного центра) сигнала. Таких нейронов около 15-20%, и они не обязательно располагаются в середине нервного центра, как это изображено на рис.1. Особенностью их является то, что они имеют на своем теле больше синаптических терминалей от сенсорных и вставочных нейронов.
Вторая группа – нейроны подпороговой каймы. Это менее возбудимые нейроны, которые не возбуждаются в ответ на поступление пороговых им-пульсов, но при действии более сильных раздражителей они возбуждаются и включаются в работу нервного центра, обеспечивая ее усиление. Таких нейронов большинство (80-85%), и они не обязательно располагаются на периферии нервного центра, но все имеют значительно меньше синаптических терминалей от сенсорных и вставочных нейронов по сравнению с нейронами центральной зоны.
На рис. 1 нейроны центральной зоны условно поставлены в центр внутреннего круга (А), а нейроны подпороговой каймы – в пространство между внутренним и наружным кругами (Б). Таким образом, если к нервному центру по афферентному входу (В) придет пороговый импульс, то возбудятся три нейрона центральной зоны, а на десяти нейронах подпороговой каймы потенциалы действия не возникнут, но появится местная деполяризация – возбуждающий постсинаптический потенциал (ВПСП).
От структуры нервного центра зависят его свойства, а они, в свою очередь, влияют на процесс проведения возбуждения через нервный центр, на его скорость и степень выраженности. От свойств нервных центров во многом зависит процесс распространения возбуждения по ЦНС, что имеет важное значение в интегративной деятельности организма.
Свойства нервных центров обусловлены описанной выше нейронной организацией нервного центра, а также химическим способом передачи возбуждения в синапсах. При электрическом способе передачи возбуждения нервные центры не имели бы подобных свойств.
Свойства нервных центров: 1 одностороннее проведение возбуждения; 2 задержка проведения возбуждения; 3 суммация; 4 облегчение; 5 окклюзия; 6 мультипликация; 7 трансформация; 8 последействие; 9 посттетаническая потенциация; 10 утомление; 11 тонус; 12 высокая чувствительность к изменению состояния внутренней среды организма; 13 пластичность.
3) Суммация – это возникновение возбуждения в нервном центре при поступлении к нему нескольких допороговых импульсов, каждый из которых в отдельности не может возбуждения (рис. 2). Фактически этот процесс происходит на нейронах подпороговой каймы. Различают два вида суммации: пространственную и временною.
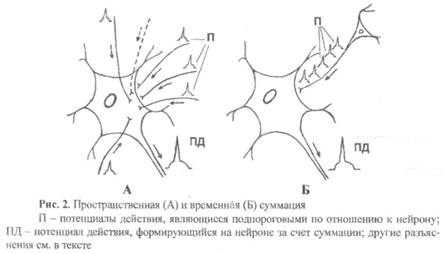
Пространственная суммация возникает в том случае, когда к нервному центру (к его нейронам) приходят одновременно, несколько допороговых импульсов. На рисунке 2А видно, что к нейрону подпороговой каймы, имеющему пороговый потенциал 30 мВ одновременно по пяти различным афферентным входам (их аксоны обозначены сплошной линией) приходят пять импульсов, каждый из которых деполяризует мембрану нейрона на 5 мВ (то есть возникают пять отдельных ВПСП). В этом случае возбуждение нейрона не наступает, так как суммарная деполяризация мембраны нейрона составляет лишь 25 мВ (суммированный ВПСП мал для достижения КУД). Но если к нейрону придет еще один подобный импульс по шестому входу (его аксон обозначен пунктирной линией), то суммированный ВПСП будет достаточен по величине и мембрана нейрона в зоне аксонного холмика деполяризуется до критического уровня, в результате чего нейрон из состояния покоя перейдет в состояние возбуждения. На постсинаптической мембране происходит суммация ВПСП в пространстве.
Временная (последовательная) суммация возникает в том случае, когда к нейронам нервного центра по одному афферентному входу приходит не один, а серия импульсов с очень небольшими по времени межимпульсными промежутками (рис. 2Б). Два механизма временной суммации:
1) интервалы между отдельными импульсами настолько малы, что за это время медиатор, выделившийся в синаптическую щель, не успевает полностью разрушиться и вернуться в пресинаптический аппарат. В этом случае возникает постепенное накопление медиатора до критического объема, необходимого для возникновения достаточного по амплитуде ВПСП, а значит, и для возникновения ПД;
2) интервалы между отдельными импульсами настолько малы, что возникший за это время на постсинаптической мембране ВПСП не успевает исчезнуть и усиливается за счет новой порции медиатора – суммируется. На постсинаптической мембране происходит суммация ВПСП во времени.
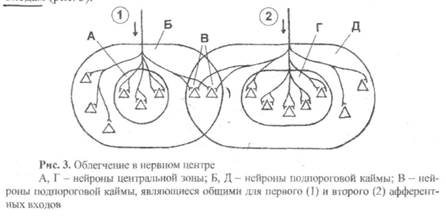
4) Облегчение – это увеличение количества возбужденных нейронов в нервном центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному, а по двум или более афферентным входам. На рис. 3 рассмотрен случай, когда при отдельном раздражении первого афферентного входа возбуждается только три нейрона центральной зоны (А), а на пяти нейронах подпороговой каймы (Б) возникают ВПСП. Если раздражать отдельно только второй афферентный вход, то возбуждены будут пять нейронов (Г), а четыре нейрона подпороговой каймы (Д) не возбудятся. Раздражая и первый, и второй афферентные входы одновременно (!), мы ожидаем вовлечения в процесс возбуждения восьми нейронов. И они, естественно, будут возбуждаться, но кроме них (сверх ожидания!) могут возбуждаться еще некоторые нейроны подпороговой каймы. Это произойдет потому, что один или несколько нейронов подпороговой каймы являются общими как для первого, так и для второго афферентных входов (в нашем случае это два нейрона, обозначенные буквой В), и при одновременном поступлении возбуждения к этим нейронам дни возбудятся за счет возникновения пространственной суммации.
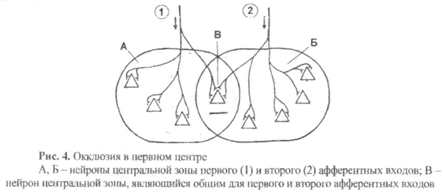
5) Окклюзия – это уменьшение количества возбужденных нейронов в нервной центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному. а по двум или более афферентным входам (рис. 4).
6) Мультипликационное возбуждение (мультипликация) заключается в том, что по разветвлениям аксона вставочного нейрона возбуждение поступает одновременно не на один, а на несколько моторных нейронов (рис. 6). В связи с этим эффект на рабочем органе усиливается в несколько раз, или в работу вовлекаются не одна, а несколько рабочих структур, Это свойство особенно ярко проявляется в ганглиях автономной (вегетативной) нервной системы.
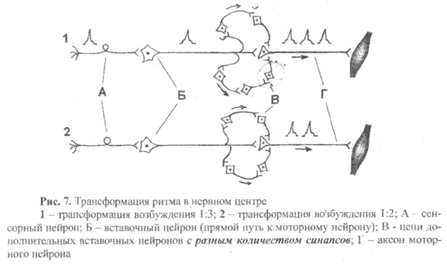
7) Трансформация ритма возбуждения – это изменение частоты импульсов на выходе из нервного центра по сравнению с частотой импульсов на входе в нервный центр.
а) наличием дублирующих цепей вставочных нейронов, связывающих сенсорные и моторные нейроны;
б) разным количеством синапсов в каждой из этих цепей.


Например, на рис.7 представлены два варианта трансформации, которые, на первый взгляд, не отличаются друг от друга, так как в том и в другом случае показаны две дополнительные цепи вставочных нейронов (кроме прямого пути), с помощью которых возбуждение может передаваться по цепи нейронов А—Б—В. Рассмотрим эти схемы.
Вариант 1. Верхняя цепь состоит из двух дополнительных вставочных нейронов, а значит, по сравнению с прямым путем передачи возбуждения с нейрона Б на нейрон В, имеет два дополнительных синапса. Поэтому возбуждение, проходя по верхней цепи, задержится на 2 мс (время синаптической задержки в одном синапсе составляет
1 мс) и придет на нейрон В после того, как пройдет возбуждение по прямому пути. В нижней цепи три дополнительных вставочных нейрона (то есть три дополнительных синапса), значит, возбуждение будет доходить до нейрона В еще дольше, чем по верхней цепи (задержка составит 3 мс). Следовательно, по нижней цепи возбуждение на нейрон В придет после того, как пройдет возбуждение по верхней цепи. В результате на один импульс, пришедший по сенсорному нейрону А, на моторном нейроне В возникнет три потенциала действия (трансформация 1:3).
Вариант 2. В этом случае и верхняя и нижняя цепи вставочных нейронов состоят из двух дополнительных нейронов. Возбуждение по обеим цепям придет к нейрону В одновременно в виде одного потенциала действие, который появится на нейроне В только после прохождения возбуждения к нему от нейрона Б по прямому пути. В этом варианте мы тоже получим трансформацию ритма, но уже в соотношении 1:2.
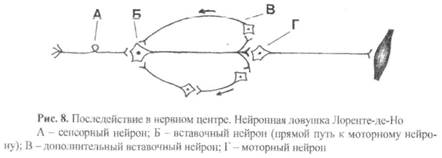
8) Последействие – это продолжение возбуждения моторного нейрона в течение некоторого времени после прекращения действия раздражителя.
Другими причинами последействия могут быть:
а) возникновение высокоамплитудного ВПСП, в результате которого возникает не один, а несколько потенциалов действия то есть ответ длится большее время;
б) длительная следовая деполяризация постсинаптической мембраны, в результате чего возникают несколько потенциалов действия, вместо одного.
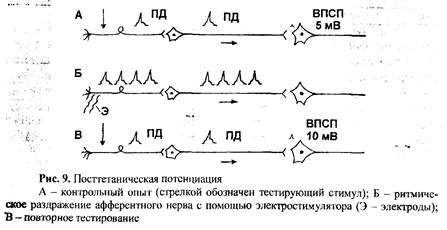
9) Посттетаническая потенциация (синаптическое облегчение) – это улучшение проведения в синапсах после короткого раздражения афферентных путей.
Если в качестве контроля вызвать одиночное раздражение афферентного нерва тестирующим раздражителем (рис. 9А), то на моторном нейроне мы получим ВПСП вполне определенной амплитуды (в нашем случае 5 мВ). Если после этого тот же афферентный нерв раздражать некоторое время серией частых импульсов (рис. 9Б), а потом вновь подействовать тестирующим раздражителем (рис. 9В), то величина ВПСП будет больше (в нашем случае 10 мВ). Причем она будет тем больше, чем более частыми импульсами мы раздражали афферентный нерв.
Длительность синаптического облегчения зависит от свойств синапса и характера раздражения: после одиночных стимулов оно выражено слабо, после раздражающей серии потенциация (облегчение) может продолжаться от нескольких минут до нескольких часов. Объясняется он тем, что при частом раздражении афферентного волокна в его пресинаптической терминали (окончании) накапливаются ионы кальция, а значит, улучшается выделение медиатора. Кроме того, показано, что частое раздражение нерва приводит к усилению синтеза медиатора, мобилизации пузырьков медиатора, к усилению синтеза белков-рецепторов на постсинаптической мембране и увеличению их чувтствительности. Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах.
10) Утомление нервного центра (посттетаническая депрессия, синаптическая депрессия) – это уменьшение или прекращение импульсной активности нервного центра в результате длительной стимуляции его афферентными импульсами (или произвольного вовлечения его в процесс возбуждения посредством импульсов, идущих из коры больших полушарий). Причинами утомления нервного центра могут быть:
• истощение запасов медиатора в афферентном или вставочном нейроне;
• снижение возбудимости постсинаптической мембраны (то есть мембраны моторного или вставочного нейрона) из-за накопления, например, продуктов метаболизма.
Утомляемость нервных центров продемонстрировал Н.Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения п. tibialis и п. peroneus. В этом случае ритмическое раздражение одного нерва вызывает ритмические сокращения мышцы, приводящие к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги. Синаптическая депрессия при длительной активации центра выражается в снижении постсинаптических потенциалов.
11) Тонус нервного центра – это длительное, умеренное возбуждение нервного центра без видимо утомления Причинами тонуса могут быть:
• потоки афферентных импульсов, постоянно поступающие с неадаптирующихся рецепторов;
• гуморальные факторы, постоянно присутствующие в плазме крови;
• спонтанная биоэлектрическая активность нейронов (автоматия);
• циркуляция (реверберация) импульсов в ЦНС.
12) Нервный центр состоит из нейронов, а они очень чувствительны к изменению состава внутренней среды организма, что и отражается на свойствах нервных центров. Наиболее важными факторами, влияющими на работу нервных центров, являются: гипоксия; недостаток питательных веществ (например, глюкозы); изменение температуры; воздействие продуктов метаболизма; воздействие различных токсических и фармакологических препаратов.
Разные нервные центры имеют неодинаковую чувствительность к воздействию названных факторов. Так, нейроны коры больших полушарий наиболее чувствительны к гипоксии, недостатку глюкозы, продуктам метаболизма; клетки гипоталамуса – к изменению температуры, содержанию глюкозы, аминокислот, жирных кислот и др.; различные участки ретикулярной формации выключаются разными фармакологическими препаратами, различные нервные центры избирательно активируются или тормозятся разными медиаторами.
13) Пластичность нервного центра означает его способность изменять при определенных обстоятельствах свои функциональные свойства. В основе этого явления лежит поливалентность нейронов нервных центров. Особенно ярко проявляется это свойство при всевозможных повреждениях ЦНС, когда организм компенсирует утраченные функции за счет сохранившихся нервных центров. Особенно хорошо свойство пластичности выражено в коре больших полушарий. Например, центральные параличи, связанные с патологией двигательных центров коры, иногда полностью компенсируются, и ранее утраченные двигательные функции восстанавливаются.
Дата добавления: 2015-01-21 ; просмотров: 21322 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Рефлексом (от лат. reflecto - отражение) называется ответная реакция организма, возникающая на раздражение рецепторов и осуществляемая с участием ЦНС.
В основе рефлекторной теории Сеченова лежат четыре принципа:
1. Структурности (структурной основой рефлекса является рефлекторная дуга)
2. Детерминизма (принцип причинно-следственных отношений). Ни одна ответная реакция организма не бывает без причины.
3. Анализа и синтеза (любое воздействие на организм сначала анализируется, затем обобщается).
4. Обратной связи (отображающий точность реакций и адаптацию), особенно развитый в трудах академика П.К. Анохина.
Итак, структурной основой рефлекса является рефлекторная дуга.
Функционально она состоит из: афферентного, центрального и эфферентного звеньев, связанных между собой синаптическими соединениями.
Морфологически она состоит из:
- рецепторных образований, назначение которых заключается в трансформации энергии внешних раздражений (информации) в энергию нервного импульса; В слизистой оболочке рта сосредоточено множество рецепторов, воспринимающих раздражения внешней и внутренней среды. Раздражение рецепторов полости рта вызывает ответные реакции в сердце, органах пищеварения, эндокринных железах и др., изменяя их деятельность.
- афферентного (чувствительного) нейрона, проводящего нервный импульс в нервный центр;
- интернейрона (вставочного) нейрона или нервного центра, представляющего собой центральную часть рефлекторной дуги;
- эфферентного (двигательного) нейрона, проводящего нервный импульс до эффектора;
- эффектора (рабочего органа), осуществляющего соответствующую деятельность. Полость рта и лицевая область могут являются эффекторным полем рефлекторного влияния с внутренних органов.
На основании принципа обратной связи необходимо дополнить рефлекторную дугу еще одним звеном – петлей обратной связи, которая устанавливает связь между исполнительной структурой и центром рефлекторной дуги или рецепторными структурами (морфологически – коллатераль аксона двигательного нейрона или другой нейрон?). Такая структура рефлекторной дуги превращает открытую рефлекторную дугу в закрытую, т.е. в самонастраивающийся нервный контур регуляции физиологической функции, оптимизируя поведение организма.
В зависимости от структуры рефлекторной дуги (т.е. количества нейронов или синаптических связей) различают моносинаптические (двухнейронные) рефлексы, например, коленный рефлекс, и полисинаптические (три и более нейронов) рефлексы.
На основании рассмотренного морфологического строения рефлекторной дуги мы уже можем заключить, что структурно-функциональной единицей нервной системы является нейрон.Действительно, в основе представления о структуре и функции ЦНС лежит нейронная теория, которая получила признание лишь на рубеже 20 века. Нервная система состоит из двух типов клеток: нейронов и клеток глии. Однако именно нейроны обеспечивают все многообразие процессов, связанных с передачей и обработкой информации.
Структура и функция нейронов и синапсов.
В каждом нейроне можно выделить 2 основных элемента:
2. отростки – дендриты и аксон с пресинаптическими окончаниями.
Тело нейрона содержит различные внутриклеточные органеллы, необходимые для обеспечения жизнедеятельности всей клетки: ядро, рибосомы, эндоплазматический ретикулум, аппарат Гольджи, митохондрии. Мембрана покрыта синапсами. Функции – синтез макромолекул, восприятие, интеграция сигналов, поступающих от других нейронов.
Дендриты – самая вариабельная часть нейрона. Сильно ветвятся, образуют различные выросты – шипики, значительно увеличивая поверхность тела нейрона. Это создает условия для размещения на дендритах большого числа синапсов. Функция – восприятие информации, передача информации к телу клетки.
Аксон – менее вариабельная часть нейрона. Начальный участок аксона называется аксональным холмиком, обладает низким порогом возбуждения по сравнению с другими участками мембраны нейрона. Аксон может образовывать коллатерали. Заканчивается аксон пресинаптическими окончаниями. Функция – проведение нервного импульса от тела клетки к другим клеткам или исполнительным органам; транспортная – внутриклеточный антероградный (синтезированные в соме активные вещества, митохондрии) и ретроградный транспорт (вирус герпеса, полиемиелита).
1. По количеству отростков –уни -, би-, мультиполярные.
2. По функции: сенсорные (чувствительные, афферентные), интернейроны (вставочные), моторные (двигательные, эфферентные).
Функции нейронов: Функции глии: 1). Защитная, опорная,
1. Интегративная; изолирующая.
2. Координирующая Трофическая ф. не доказана, но
3. Трофическая выявлена роль в процессах памяти.
Итак, уникальными особенностями нейрона являются способность генерировать электрические разряды и передавать информацию с помощью специализированных межклеточных соединений – синапсов. Являясь главным механизмом связи между нейронами, синапсы во многом обеспечивают все многообразие функций мозга.
Понятие синапсбыло введено в физиологию англ. физиологом Ч. Шеррингтоном в 1897 году для обозначения функционального контакта между нейронами. Синапсы различаются по:
- механизму действия (электрический, химический, смешанный);
- локализации на поверхности клетки (аксосоматические, аксодендрические, реже - аксоаксонные, дендродендрические);
- функции (возбуждающие или тормозящие);
- способностью к модуляции?[K1]
Синапс представляет собой сложное функциональное образование и состоит из:
- постсинаптической мембраны (субсинаптическая мембрана).
Мы разберем строение и работу наиболее часто встречающихся у человека химических синапсов.
Пресинаптическая мембрана, как мы уже говорили, это окончание аксона.
Синаптическая щель широкая и составляет в среднем 10-20 нм, подавляющая часть пресинаптического тока шунтируется низким сопротивлением щели. Поэтому в химических синапсах существует особый механизм, способный изменить мембранный потенциал постсинаптической мембраны и таким образом передать информацию от клетки к клетке. Таким механизмом является выделение пресинаптическим окончанием аксона особых химических веществ – нейротрансмиттеров (или медиаторов), которые, воздействуя на специфические рецепторы постсинаптической мембраны, способны изменять состояние ионных каналов постсинаптической мембраны.
В настоящее время выявлено более 15 нейротрансмиттеров (ацетилхолин, норадреналин, серотонин, глицин, гамма-аминомаслянная кислота и др.). Существует правило Дейла – каждый нейрон во всех своих пресинаптических окончаниях выделяет один и тот же медиатор, поэтому нейроны или синапсы иногда обозначают по типу медиатора (холинергические, адренергические, серотонинергические и др.). Однако как из каждого правила существуют исключения, так и в данном случае правило Дейла не абсолютно. Экспериментально обнаружена возможность выделения одним и тем же пресинаптическим окончанием множества других нейротрансмиттеров.
Постсинаптическая мембрана (а именно субсинаптическая) имеет специализированные рецепторы и хемочувствительные ионные каналы.
Разберем механизм синаптической передачи.
В пресинаптических окончаниях аксона депонируется определенное количество нейротрансмиттеров (медиаторов) в синаптических пузырьках или везикулах, каждая из которых содержит один квант медиатора, состоящий из нескольких тысяч молекул.
Деполяризация пресинаптической мембраны под действием распространяющегося импульса увеличивает проницаемость для Са 2+ . При наличии Са 2+ везикула, подойдя к внутренней поверхности мембраны пресинаптического окончания, сливается с пресинаптической мембраной. В результате происходит выброс кванта медиатора в синаптическую щель. Таким образом, выход кванта нейротрансмиттера (медиатора) из везикул представляет собой процесс экзоцитоза. После спадения везикулы окружающая его мембрана включается в мембрану пресинаптического окончания. В дальнейшем в результате эндоцитоза небольшие участки пресинаптической мембраны впячиваются внутрь, вновь образуя везикулы, которые снова способны включать нейротрансмиттер (медиатор) и вступать в цикл его освобождения.
Участие Са 2+ определяет ряд важных особенностей работы химических синапсов. Прежде всего, время, необходимое для вхождения Са 2+ внутрь клетки, определяет синаптическую задержку проведения, составляющую 0,2-0,5 мс. Кроме того, накопление Са 2+ внутри пресинаптического окончания в результате предшествующего поступления улучшает эффективность работы химического синапса, т.е. вызывает высвобождение большего числа квантов медиатора, что проявляется увеличением амплитуды постсинаптических потенциалов.
В возбуждающем синапсе происходит увеличение проницаемости для Nа + или Са 2+ , в результате чего происходит деполяризация мембраны и возникает локальный ответ – ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ (ВПСП).
В тормозном синапсе происходит увеличение проницаемости для К + , в результате чего происходит гиперполяризация мембраны и возникает локальный ответ - ТОРМОЗНОЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ (ТПСП).
Характер действия медиатора определяется свойствами субсинаптической мембраны.
Следует еще раз подчеркнуть, что на субсинаптической мембране возникает лишь локальный ответ. Возникновение потенциала действия (ПД) за счет суммации локальных ответов происходит в специальной низкопороговой области (зона аксонного холмика), откуда ПД распространяется по аксону и на мембрану соседних участков клетки.
Неиспользованный медиатор или его фрагменты всасываются обратно в пресинаптическую часть синапса.
Нейрон имеет множественные синаптические контакты с другими нейронами. Два принципа: дивергенции и конвергенции.
Дивергенция – это способность нервной клетки устанавливать многочисленные синаптические связи с различными нервными клетками. В результате афферентная информация поступает одновременно к разным участкам ЦНС, один нейрон может участвовать в нескольких различных реакциях, передавать возбуждение значительному числу других нейронов, обеспечивая широкую иррадиацию возбудительного процесса в центральных нервных образованиях (кашлевой рефлекс).
Конвергенция – это схождение различных импульсных потоков от нескольких нервных клеток к одному нейрону. Интегративная функция. Мотонейрон – общий конечный путь двигательной системы (англ. физиол. Шеррингтон)
Понятие о нервном центре, его свойства.
Если нейрон можно рассматривать как структурную единицу нервной системы, то физиологической системной единицей является нервный центр.
Нервный центр – это динамическая совокупность нейронов, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт.
Функциональный нервный центр может быть локализован в разных анатомических структурах. Например, дыхательный центр представлен нервными клетками, расположенными в спинном, продолговатом, промежуточном мозге, в коре головного мозга.
Нервные центры имеют ряд общих свойств, определяемых наличием синаптических образований и структурой нейронных сетей, образующих эти центры:
1. Низкая лабильность. Обусловлена скоростью развития синаптической передачи импульса в химическом синапсе. 50-100 имп/с.
2. Высокая утомляемость. Утомление – временное снижение работоспособности в результате проведенной работы, которое исчезает после отдыха. Причины: а). истощение и несвоевременный синтез медиатора; б). адаптация постсинаптического рецептора к медиатору; в). инактивация рецепторов в результате длительной деполяризации постсинаптической мембраны.
3. Высокая чувствительность к недостатку кислорода. Мозг в 22 раза больше потребляет кислорода, чем мышечная ткань. Необратимые изменения наступают в коре через 4-5 мин, в стволовых клетках – через 15-20 мин.
4. Высокая чувствительность к фармакологическим веществам (блокаторы нервно-мышечной передачи, психомиметические средства), ядам.
Токсин Cl. вotulinum – блокада высвобождения возбуждающего медиатора.
Столбнячный токсин – блокада высвобождения тормозного медиатора.
Удаление Са 2+ или замещение на Мg 2+ - блокада высвобождения медиатора
Гемихолиний – нарушение синтеза медиатора.
Бунгаротоксин – необратимое связывание с АХ-рецепторами
Яд кураре – конкурентное связывание с АХ-рецепторами.
Стрихнин – конкурентное связывание с глицин-рецепторами.
Бикулин, пикротоксин (судорожные яды), пенициллин – конкурентное связывание с ГАМК-рецепторами.
Фосфоорганические соединения – угнетение холинэстеразы и вследствие этого продолжительная субсинаптическая деполяризация и инактивация рецепторов.
Релаксанты (сукцинилхолин) – аналоги АХ, но не расщепляющиеся АХЭ, продолжительная деполяризация субсинаптической мембраны и инактивация рецепторов.
Алкоголизм, привыкание, наркомания.
5. Пластичность. Основа: изменение структуры и функции синапсов. Поэтому синапсы участвуют в таких функциях ЦНС как научение и память, компенсации функции при нарушении за счет формирования новых нейронных связей; синтеза специфических белков.
Особенности проведения возбуждения по нервному центру определяют закономерности проведения возбуждения по рефлекторной дуге:
1. Одностороннее проведение. Обусловлено особенностями проведения возбуждения по синапсу. Медиаторы выделяются в пресинаптическом окончании, рецепторы находятся в постсинаптической мембране.
2. Замедленное проведение, обусловленное синаптической задержкой в центральной части рефлекторной дуги. Составляет 0.2-0.5 мс и определяет время рефлекса (от начала раздражения до начала ответной реакции).
3. Суммация подпороговых раздражений. Например, чихательный рефлекс. И.М. Сеченов открыл, изучил. Два типа:
1. Суммация последовательная (временная);
2. Суммация пространственная (одновременная)
П/п раздражения наносятся одновременно на несколько точек рецептивного поля, в результате конвергенции происходит суммация локальных ответов.
4. Трансформация ритма возбуждения.
При ритмическом возбуждении нервный центр перестраивает ритм как понижая, так и повышая частоту следования импульсов. Понижение связано с низкой лабильностью синапса (максимально – 100 имп/с). Повышение обусловлено: - возникновением повторных разрядов на фоне длительной следовой деполяризации; - наличием полисинаптических нервных цепей; - циркуляцией импульсов в замкнутых нейронных цепях.
5. Рефлекторное последействие – продолжение рефлекторной реакции после окончания действия раздражителя. Механизмы те же, что и механизмы повышающей трансформации.
6. Ритмическая активация синапса часто сопровождается значительным увеличением амплитуды синаптических потенциалов. Посттетаническая потенциация – это усиление рефлекторного ответа после тетанических раздражений. Длительность посттетанической потенциации может составлять от нескольких минут до нескольких часов. С функциональной точки зрения посттетаническая потенциация представляет собой процесс облегчения в ЦНС, связанный с приобретением опыта, т.е. процесс научения, памяти.
1). Ритмическая активация мембраны пресинаптического окончания аксона ведет к увеличению амплитуды ПД, что вызовет высвобождение большего количества медиатора в синаптическую щель.
2). Ритмическая активация сопровождается увеличением запаса доступного медиатора, готового к выделению.
3). Ритмическая активация приводит к вхождению и накоплению Са 2+ в пресинаптическом окончании(мы уже с вами об этом упоминали при рассмотрении работы синапса), что увеличивает высвобождение медиатора в синаптическую щель. Это самый важный фактор для посттетанической потенциации.
Читайте также:
