
Сохранение следа памяти в нервной системе
Сохранение следов в нервной системе
Лишь за последние два десятилетия положение дела существенно изменилось:
Явления длительного сохранения следов раздражителя отмечались исследователями на всем протяжении развития животного мира.
Многократно наблюдался факт, что однократное раздражение электрическим ударом нервной системы полипов вызывало появление ритмических электрических импульсов, которые могли сохраняться в течение многих часов.
Подобные же явления можно было наблюдать при исследовании работы центральной нервной системы животных. Так, однократное раздражение вспышкой света вызывало в верхнем двухолмии кролика ритмические электрические разряда, которые можно было регистрировать в течение достаточно длительного времени, причем такие реакции можно было наблюдать даже при отведении токов действия от изолированного нейрона.
Продолжение электрических разрядов, возникающих после единичного раздражения, показывает, что нейроны являются не только аппаратами, принимающими сигналы и реагирующими на них соответствующими отделами, но и то, что они сохраняют следы раздражителя, продолжая давать запущенные этим раздражителем ритмические ответы долгое время после того, как этот раздражитель прекратил свое влияние.
Это последействие влияний раздражителя и есть наиболее элементарное проявление психологической памяти, которое можно наблюдать как на отдельном нейроне, так и на работе всей нервной системы в целом.
Наиболее элементарные физиологические проявления памяти можно наблюдать и другим путем, о котором мы уже упоминали в предыдущей главе.
Как показали исследования, длительное повторение одного и того же сигнала приводит к привыканию к нему, которое проявляется в исчезновении ориентировочных рефлексов на этот раздражитель, ставший привычным. Как показал советский психолог Е. Н. Соколов, такие явления привыкания можно наблюдать даже при исследовании ответов изолированного нейрона на многократно повторяемые раздражители.
Наиболее характерным является тот факт, что при небольшом изменении интенсивности или характера раздражителя признаки ориентировочного рефлекса появляются снова.
Данные, полученные Е. Н. Соколовым и его сотрудниками, показали, что явление растормаживания ранее угасшего ориентировочного рефлекса можно наблюдать не только сразу после изменения характера раздражителя, но и через некоторые, иногда довольно значительные промежутки времени. Так, если у испытуемого вырабатывалось явление привыкания к определенному раздражителю, то достаточно было изменить интенсивность, продолжительность или характер раздражителя, чтобы вегетативные или электрофизиологические симптомы ориентировочного рефлекса восстановились, причем это растормаживание (восстановление) знаков ориентировочного рефлекса наблюдалось после достаточно значительных промежутков времени после угашения. Этот факт можно было наблюдать как при регистрации симптомов ориентировочного рефлекса нервной системы в целом, так и на уровне отдельного нейрона. [8]
Нервная система может с удивительной тонкостью сохранять следы прежних раздражителей, что можно иллюстрировать и целым рядом дальнейших наблюдений.
Факты дальнейших исследований показали, что нервная система человека может сохранять следы отдельных сигналов с очень большой степенью точности и хранить их длительное время. Иллюстрацией этого может служить опыт, проведенный в лаборатории Е. Н. Соколова.
Испытуемому однократно предъявлялся звуковой сигнал определенной высоты (500 гц) и интенсивности (20 дБ). В ответ на этот сигнал он должен был сжимать руки, причем его предупреждали, что он должен отвечать движением только в ответ на этот сигнал и не двигать рукой при предъявлении любого отличающегося сигнала. Затем испытуемому в беспорядке предъявлялись разные звуки той же высоты, но варьирующие по интенсивности (от 5 дБ до 30 дБ). Регистрировалась электроэнцефалограмма, эдектромиограмма и кожно-гальваническая реакция. Такой же опыт повторялся на 2-й, 4-й и 25-й день, причем однократно показанный эталон (звук в 500 гц интенсивностью в 20 дБ) более ни разу не предъявлялся.
Результаты опыта показали, что один раз показанный эталон сохранялся в течение длительного времени, и после длительных промежутков (от двух до 25 суток) испытуемый продолжал давать четкие электрофизиологические и двигательные реакции только на сигналы, соответствующие данному эталону, и ни на какие другие.
Приведенный опыт показывает, что мозг человека оказывается в состоянии сохранять четкие следы раз предъявленного раздражителя на очень длительное время, причем точность этих следов не только не исчезает со временем, но, возможно, возрастает.
Это делает мозг человека тончайшим прибором не только для улавливания раздражителей и их выделения из числа других, доходящих до него раздражителей, но и для сохранения в памяти следов тех воздействий, которые ранее воспринимались им. [8]
Физиологические основы памяти
Явления длительного сохранения следов раздражителя отмечались исследователями на всем протяжении развития животного мира.
Многократно наблюдался факт, что однократное раздражение электрическим ударом нервной системы полипов вызывало появление ритмических электрических импульсов, которые могли сохраняться в течение многих часов.
Подобные же явления можно было наблюдать при исследовании работы центральной нервной системы животных. Так, однократное раздражение вспышкой света вызывало в верхнем двухолмии кролика ритмические электрические разряда, которые можно было регистрировать в течение достаточно длительного времени, причем такие реакции можно было наблюдать даже при отведении токов действия от изолированного нейрона.
Продолжение электрических разрядов, возникающих после единичного раздражения, показывает, что нейроны являются не только аппаратами, принимающими сигналы и реагирующими на них соответствующими отделами, но и то, что они сохраняют следы раздражителя, продолжая давать запущенные этим раздражителем ритмические ответы долгое время после того, как этот раздражитель прекратил свое влияние. Это последействие влияний раздражителя и есть наиболее элементарное проявление психологической памяти, которое можно наблюдать как на отдельном нейроне, так и на работе всей нервной системы в целом.
Наиболее элементарные физиологические проявления памяти можно наблюдать и другим путем, о котором мы уже упоминали в предыдущей главе.
Как показали исследования, длительное повторение одного и того же сигнала приводит к привыканию к нему, которое проявляется в исчезновении ориентировочных рефлексов на этот раздражитель, ставший привычным. Как показал советский психолог Е. Н. Соколов, такие явления привыкания можно наблюдать даже при исследовании ответов изолированного нейрона на многократно повторяемые раздражители.
Наиболее характерным является тот факт, что при небольшом изменении интенсивности или характера раздражителя признаки ориентировочного рефлекса появляются снова.
Тот факт, что нервная система может с удивительной тонкостью сохранять следы прежних раздражителей, можно иллюстрировать и целым рядом дальнейших наблюдений, из которых мы приведем только два.
Известно, что чем чаще встречается определенный сигнал, чем больше испытуемый привыкает к нему, тем быстрее он дает на него двигательную реакцию (тем меньше латентный период этой реакции). Тщательное исследование показало, что в самых простых условиях этот закон сохраняется и быстрота реакции на сигнал прямо пропорциональна частоте, с которой он предъявляется.
Факты дальнейших исследований показали, что нервная система человека может сохранять следы отдельных сигналов с очень большой степенью точности и хранить их длительное время. Иллюстрацией этого может служить опыт, проведенный в лаборатории Е. Н. Соколова.
Испытуемому однократно предъявлялся звуковой сигнал определенной высоты (500 гц) и интенсивности (20 дБ). В ответ на этот сигнал он должен был сжимать руки, причем его предупреждали, что он должен отвечать движением только в ответ на этот сигнал и не двигать рукой при предъявлении любого отличающегося сигнала. Затем испытуемому в беспорядке предъявлялись разные звуки той же высоты, но варьирующие по интенсивности (от 5 дБ до 30 дБ). Регистрировалась электроэнцефалограмма, эдектромиограмма и кожно-гальваническая реакция. Такой же опыт повторялся на 2-й, 4-й и 25-й день, причем однократно показанный эталон (звук в 500 гц интенсивностью в 20 дБ) более ни разу не предъявлялся.
Результаты опыта показали, что один раз показанный эталон сохранялся в течение длительного времени, и после длительных промежутков (от двух до 25 суток) испытуемый продолжал давать четкие электрофизиологические и двигательные реакции только на сигналы, соответствующие данному эталону, и ни на какие другие.
Приведенный опыт показывает, что мозг человека оказывается в состоянии сохранять четкие следы раз предъявленного раздражителя на очень длительное время, причем точность этих следов не только не исчезает со временем, но, возможно, возрастает.
Мы привели некоторые факты, говорящие о том, что нервная система обладает способностью длительно сохранять следы предъявленного раздражителя, оценивать частоту, с которой он предъявлялся, и удерживать в памяти с большой точностью те эталоны раздражителей, которые предъявлялись хотя бы один раз.
Это делает мозг человека тончайшим прибором не только для улавливания раздражителей и их выделения из числа других, доходящих до него раздражителей, но и для сохранения в памяти следов тех воздействий, которые ранее воспринимались им.
Явления длительного сохранения следов раздражителя отмечались исследователями на всем протяжении развития животного мира.
Многократно наблюдался факт, что однократное раздражение электрическим ударом нервной системы полипов вызывало появление ритмических электрических импульсов, которые могли сохраняться в течение многих часов.
Подобные же явления можно было наблюдать при исследовании работы центральной нервной системы животных. Так, однократное раздражение вспышкой света вызывало в верхнем двухолмии кролика ритмические электрические разряда, которые можно было регистрировать в течение достаточно длительного времени, причем такие реакции можно было наблюдать даже при отведении токов действия от изолированного нейрона.
Продолжение электрических разрядов, возникающих после единичного раздражения, показывает, что нейроны являются не только аппаратами, принимающими сигналы и реагирующими на них соответствующими отделами, но и то, что они сохраняют следы раздражителя, продолжая давать запущенные этим раздражителем ритмические ответы долгое время после того, как этот раздражитель прекратил свое влияние.
Наиболее элементарные физиологические проявления памяти можно наблюдать и другим путем, о котором мы уже упоминали в предыдущей главе.
Как показали исследования, длительное повторение одного и того же сигнала приводит к привыканию к нему, которое проявляется в исчезновении ориентировочных рефлексов на этот раздражитель, ставший привычным. Как показал советский психолог Е. Н. Соколов, такие явления привыкания можно наблюдать даже при исследовании ответов изолированного нейрона на многократно повторяемые раздражители.
Наиболее характерным является тот факт, что при небольшом изменении интенсивности или характера раздражителя признаки ориентировочного рефлекса появляются снова.
Тот факт, что нервная система может с удивительной тонкостью сохранять следы прежних раздражителей, можно иллюстрировать и целым рядом дальнейших наблюдений, из которых мы приведем только два.
Известно, что чем чаще встречается определенный сигнал, чем больше испытуемый привыкает к нему, тем быстрее он дает па него двигательную реакцию (тем меньше латентный период этой реакции).
Факты дальнейших исследований показали, что нервная система человека может сохранять следы отдельных сигналов с очень большой степенью точности и хранить их длительное время. Иллюстрацией этого может служить опыт, проведенный в лаборатории Е. Н. Соколова.
Испытуемому однократно предъявлялся звуковой сигнал определенной высоты (500 гц) и интенсивности (20 дБ). В ответ на этот сигнал он должен был сжимать руки, причем его предупреждали, что он должен отвечать движением только в ответ на этот сигнал и не двигать рукой при предъявлении любого отличающегося сигнала. Затем испытуемому в беспорядке предъявлялись разные звуки той же высоты, но варьирующие по интенсивности (от 5 дБ до 30 дБ). Регистрировалась электроэнцефалограмма, эдектромиограмма и кожно-гальваническая реакция. Такой же опыт повторялся на 2-й, 4-й и 25-й день, причем однократно показанный эталон (звук в 500 гц интенсивностью в 20 дБ) более ни разу не предъявлялся. Результаты опыта показали, что один раз показанный эталон сохранялся в течение длительного времени, и после длительных промежутков (от двух до 25 суток) испытуемый продолжал давать четкие электрофизиологические и двигательные реакции только на сигналы, соответствующие данному эталону, и ни на какие другие.
Приведенный опыт показывает, что мозг человека оказывается в состоянии сохранять четкие следы раз предъявленного раздражителя на очень длительное время, причем точность этих следов не только не исчезает со временем, но, возможно, возрастает.
Мы привели некоторые факты, говорящие о том, что нервная система обладает способностью длительно сохранять следы предъявленною раздражителя, оценивать частоту, с которой он предъявлялся, и удерживать в памяти с большой точностью те эталоны раздражителей, которые предъявлялись хотя бы один раз.
Это делает мозг человека тончайшим прибором не только для улавливания раздражителей и их выделения из числа других, доходящих до него раздражителей, но и для сохранения в памяти следов тех воздействий, которые ранее воспринимались им.
Факт запечатления следов тех раздражителей, которые воздействовали на мозг человека, заставляет поставить важные вопросы:
• как протекает процесс закрепления этих следов?
• закрепляются ли они сразу или для их закрепления нужно некоторое время? Эти вопросы были предметом целого ряда исследований.
Уже раньше было сделано наблюдение, что в тех случаях, когда человек испытывает травму черепа, следы от раздражителей, которые воздействовали на человека за короткое время до травмы и в некоторый промежуток после травмы, не сохраняются. Человек, получивший массивную травму черепа с потерей сознания, обычно не сохраняет никаких воспоминаний о том, что непосредственно предшествовало травме и что следовало за ней. Этот факт широко известен и получил название антероградной и ретроградной амнезии. Он указывает на то, что сильный шок, который испы-
тывает нервная система, делает мозг на некоторое время неспособным запечатлевать следы дошедших до него раздражений.
Описанные факты были толчком для специальных экспериментов, в которых человеку давался искусственный ослабленный шок, и просматривалось, какой промежуток времени выпадает из его мозга.
Примером могут служить опыты, проведенные советским психофизиологом Ф. Д. Горбовьш.
Испытуемый помещался перед окошечком, через которое плавно проходили простые цифры с арифметическими знаками (+4, -1, -8, +5 и т. д.). Испытуемый должен был произвести соответствующие арифметические операции, прибавляя данное число к результату ранее полученных операций или вычитая из него соответствующее число. Естественно, что для выполнения этой задачи он должен был прочно удерживать в памяти следы ранее полученного результата.
приводило к грубому разрушению следов, применение его через 10 мин — к относительно слабому сохранению следов, а применение его через 20 мин оставляло навык сохранным.
Первый вопрос, который возникает перед исследователями нервной системы и когнитивных процессов это, что такое память? Что такое память в биологическом аспекте? Как проявляется память на уровне отдельного нейрона? И в какой форме хранится информация в нервной системе?
Что такое память в биологическом аспекте и на уровне отдельного нейрона? Уже давно бытует мнение в нейронауке, что память связана с изменением состояний и силы синапсов, а также чувствительности самого нейрона. О том, как может происходить изменения этих состояний, мы говорили в прошлом выпусках, когда говорили о привыкании и сенсибилизации на примере моллюска Аплизии а также когда изучали механизмы образования условных рефлексов на базе ассоциативного нейроэлемента. Такое представление памяти породило понятие коннектом, что представляет собой полное описание всех связей в нервной системе с учётом качества и свойств этих связей. Можно сказать что Вы, ваша личность это ваш коннектом. Если когда-то представится возможность полностью повторить Ваш коннектом, то получатся личность идентичная Вам с теми же воспоминаниями и навыками.
В какой форме хранится информация в нервной системе? Если говорить о нервной системе моллюска, то все ограниченно настройками синапсов в цепочках нейронов уже имеющихся рефлексов. Но если обратится к нервной системе млекопитающих, то она обладает инструментом позволяющим формировать рефлексы, которые небыли заданны у животного от рождения. Рефлексы в нервной системе представлены виде рефлекторных дуг или цепочек нейронов, по которым последовательно передается нервное возбуждение. Можно представить, что цепочки нейронов формируют целые схемы путей, по которым распространяется возбуждение, одни пути очень крупные подобно магистралям другие более мелкие подробно тропинкам и все это подвластно некоторым перестройкам и изменением.
Когда говорим о рефлекторных дугах, то представляются последовательная цепочка единичных нейронов, но на самом деле для центральной нервной системы свойственно, что в рефлекторную дугу вовлечены сразу множество нейронов и эти передающие цепочки состоят из целого массива клеток. Хотя, по сути, каждая клетка из этого массива содержит практически одинаковую информацию и, удалив не одну, а целые регионы клеток можно сохранить рефлекс, к которому они принадлежали. Это может показаться нерациональным сточки зрения экономии пространства и ресурсов, но с позиции надежности сохранности информации это оправданно. Ведь нервная клетка может быть подвержена гибели из-за различных неблагоприятных условий, гипоксии, недостатка питательных веществ или токсичного отравления и не желательно, чтобы смерть одной клетки приводил к возможной потери важной информации для организма. Получается, что невозможно оценить достоверно объемы информации оперируемой мозгом по количеству нейронов и связей между ними.
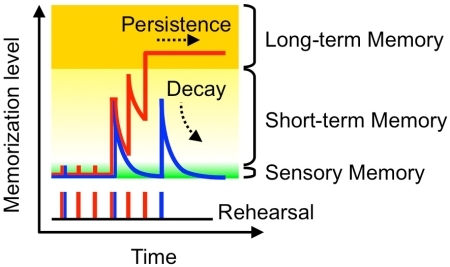
В чем природа различной степени глубины запоминания? Физиологами уже давно было выделено два вида памяти временная или оперативная память и долговременная память. То есть в некоторых ситуациях мы запоминаем информацию на определенный небольшой срок и если не возвращаемся к ней, то мы ее забываем. А в других ситуациях мы запоминаем информацию на неограниченный срок, если эта информация для нас важна, то она может сохраниться даже сразу. Эволюционно эти два вида памяти обоснованны тем, что организму не требуется хранить все, что с ним происходит, особенно ели эта информация не важна для выживания и успешности и никогда больше не повторяется, ведь хранение информации требует траты ресурсов.
Переход информации из временной памяти в долговременную память называется консолидацией памяти. На эту тему существует множество исследований, и в работе над моделью я стремился обосновать наличие и имитировать различную глубину запоминания. И изучая материалы этих исследований, я пришел к выводу, что возможно мы имеем дело не с одним видом консолидации памяти, а как минимум с двумя.
Первое это консолидация на физиологическом уровне, это эксперименты, в которых, к примеру, птенцов обучает некоторому рефлексу и если ничего не происходит, то рефлекс сохранится в памяти, но если в течение двадцати минут будет оказанное действия электрошока или токсина, то рефлекс не будет сохранен в памяти. Это значит, что в течение этих двадцати минут происходят процессы, связанные с сохранением информации в памяти, которые можно нарушить, тем самым предотвратить это сохранение.
Изменения в структуре клетки не происходят моментально при её обучении, на это требуется некоторое время, связанное с целым рядом химических реакций и превращений, производством необходимых белков и ростом новых отростков. Но следствие обучения мы наблюдаем иногда практически моментальным, достаточно нам услышать какую-то информацию и мы уже ей обладаем и можем оперировать ей. Предположительно это связанно с тем, что нервная клетка обладает неким резервом к изменениям, к примеру, в синапсах может храниться дополнительные везикулы с медиатором, которые будут выделяться только в условиях обучения. Но после обучения для сохранения информации потребуется увеличение синапса и общего содержания в нем медиаторов с учётом резерва на возможное обучение. В таком случае синапс маркируется специальными белками, а так же запускается определенный сложный каскад химических реакций, который и приводит к росту синапса. Нарушение хода этого сложного каскада химических реакций в течение необходимых для этого времени и приводит к нарушению запоминания.
Но в компьютерной модели нет необходимости отражать физиологическую консолидацию памяти, потому что все изменения с виртуальными синапсами мы можем проводить моментально.
Следующая форма консолидации памяти является примером того, что повторенная несколько раз информация усваивается надолго, а информация, которая не имеет эмоционального окраса и была представлена единожды или довольно малое количество, через некоторое время будет забыта. Данный вид консолидации я называю иерархической, давайте рассмотрим данный вид консолидации на примере усвоения обычного условного рефлекса и с использованием модели.
В предыдущей части демонстрировался пример того как образуется условный рефлекс на базе ассоциативных нейроэлементов. Теперь давайте разберемся, как формируется условный рефлекс на уровне центральной системы. Обычно безусловные рефлексы имеют представительства сразу в нескольких областях нервной системы, сначала это представительство в области спинного мозга, если двигаться дальше по иерархии, то представительства в каких-нибудь ганглиях, затем в древнем мозге, к примеру, в гиппокампе и наконец, представительство в самой коре.
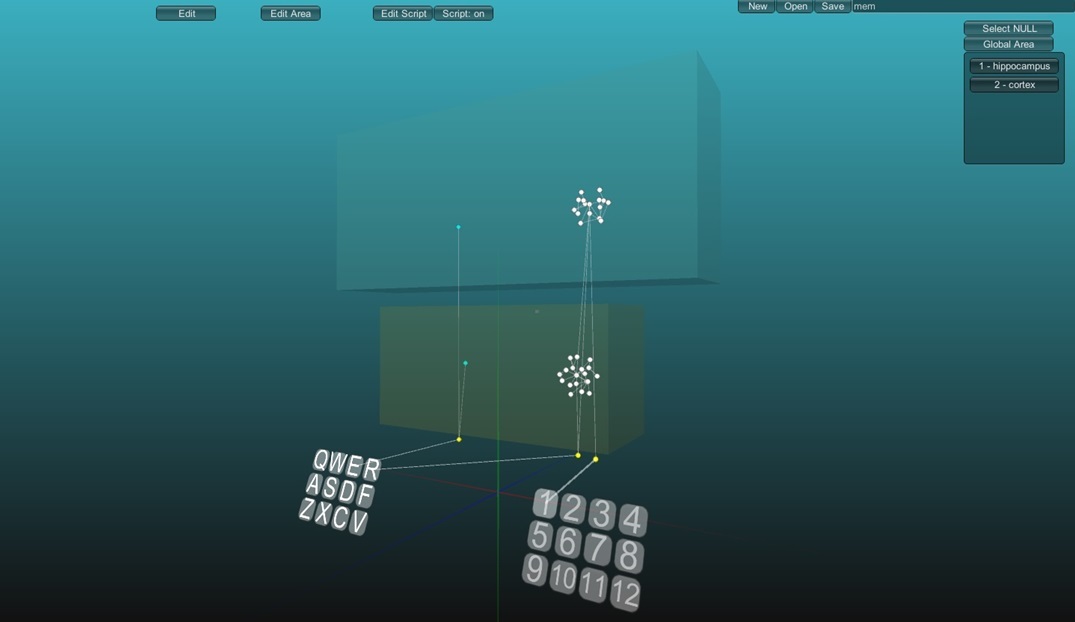
На модели Вы видите две выделенные области, они изолированы, т.е. нейроэлементы принадлежащие этим областям не оказывают друг на друга действия, если конечно не связанны синапсами напрямую. Я условно обозначил эти две области как “hippocampus” (гиппокамп), область находящиеся ниже и “cortex” (кора) область находящиеся сверху. Так же существует рефлекс с заголовком “R” приводящий к рефлекторному ответу “1”. У этого рефлекса существуют два представительства в гиппокампе и коре, это две группы белых нейроэлементов. А также есть два рецепторных нейроэлемента связанных с рецептором “E” в каждой области, эти нейроэлементы индифферентны, и активация их не приведет ни к какому ответу, а создаст случайную активность. Главное это то, что нейпропластичность для этих двух областей различная, в области обозначенной как гиппокамп пластичность выше, чем в области коры.
Если мы активизируем рефлекс с заголовком “R” и безусловный раздражитель “E”, примерно в одном временном интервале, то произойдет образование новой рефлекторной дуги, но быстрее всего это произойдет в области с высокой пластичностью.
Рефлекс образованный только в области с высокой пластичностью будет полностью функционален, но может быть утерян потому, что под влиянием других раздражителей нейроны могут переобучаться. Гиппокамп значительно меньше коры, но в нем присутствуют практически все представительства, что и в коре. Необходимость в образовании новых связей всё время заставляет использовать клетки, которые уже участвовали в образовании рефлексов.
Если мы продолжим обучение сочетанием раздражителей “E” и “R”, то через некоторое количество повторений сформируется рефлекторная дуга и в области с низкой пластичностью.
Теперь рефлекс защищен более надежно, даже если он пропадет из области с высокой пластичностью, он все равно будет выполняться.
Конечно, переобучение нейронов в области с низкой пластичностью возможно, но для этого потребуется больше времени и усилий.
Таким образом, существует два этапа запоминания: до образования рефлекса в коре и после. Сохранение копии рефлекса не только в гиппокампе но и в коре головного мозга является иерархической консолидацией памяти.
Но как тогда обстоит дело с информацией, которая запоминается сразу и надолго, обычно такая информация сопровождается какими-либо эмоциональными переживаниями. К примеру, человеку достаточно однажды обжечься от кастрюли на плите, что бы помнить об этом всю жизнь.
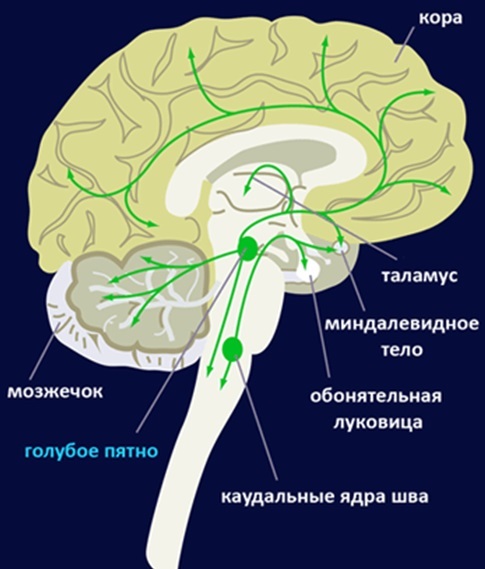
Норадреналин в нашем случае является сигналом к повышению нейропластичности. Это своего рода команда “now print” (“теперь пиши”) и под влиянием норадреналина нервные клетки стараются как можно быстрее производить изменения в себе. И соответственно, когда действия норадреналина прекращаются клетки возвращаются к своему рабочему состоянию, сохраняя все изменения, которые с ними происходили.
Итак, давайте рассмотрим это в модели.
Области в программе не только логически отделяют и изолируют нейроэлементы, но и существует возможность настраивать некоторые сценарии взаимодействия этих областей.
Система организации в модели можно охарактеризовать как нечто подобное гиперсети (сеть сетей) из лекций Константина Анохина. Но объекты сети более высокого уровня – это различные области мозга. Характер их взаимодействия в системе особый, активность одной области может приводить к торможению всех нейронов другой, или оказывать модулирующий характер, или влиять на пластичность. Чувство страха (активность миндалины) приводит к повышению чувствительности нейронов моторной коры, т.е. происходит модуляция по снижению порога активации нейронов. Поэтому, когда нас охватывает страх, мы можем быстрее убегать от опасности, наши мускулы не становятся при этом сильнее, просто требуется меньше внутреннего мотива к действию.
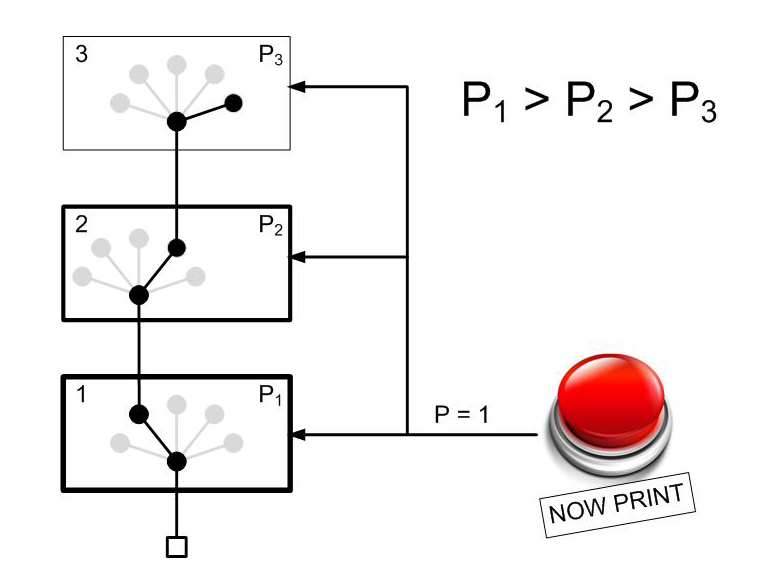
Существуют сгруппированные сети нейроэлементов, а так же эти группы можно организовать в сети уже со своими характерами взаимодействия. В данном случае существует сценарий, по которому при возникновении активности в области именуемой “amigdala” (миндалина) в области “cortex” происходит увеличение пластичности на две секунды.
Без активации миндалины:
С активацией миндалины:
Таким образом, получается, что в состоянии стресса обучение происходит в коре так же быстро, как и в областях с высокой пластичностью.
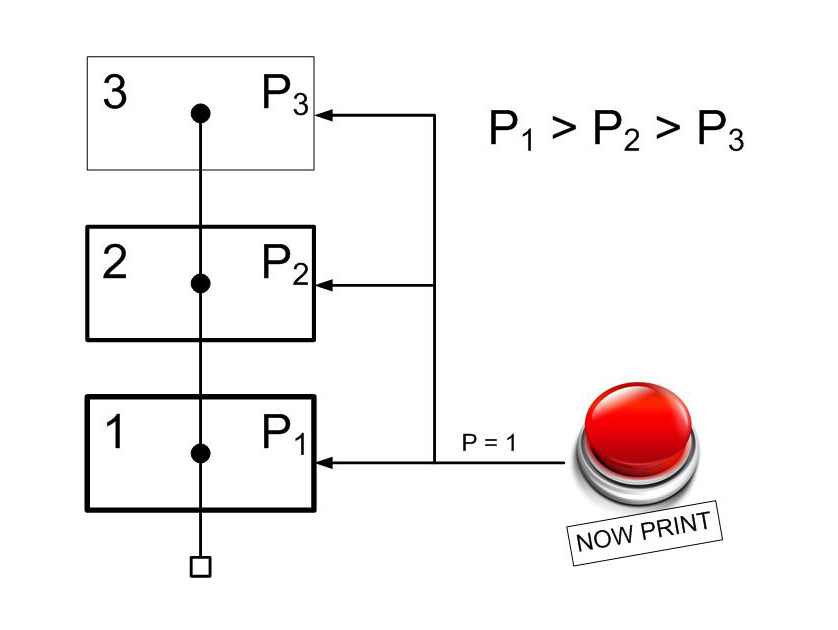
Иерархическую систему памяти можно представить следующим образом. Существует несколько изолированных областей, в каждой последующей пластичность будет меньше, а значит, каждая следующая область будет в меньшей степени подвергаться изменениям. В каждой такой области будет существовать представительство раздражителя. А так же будет возможность управления скоростью запоминания посредством сигналов, приводящих к кратковременному изменению пластичности.
Образы
Рассматривать человеческую память только на основе простейших условных рефлексов неправильно, для многих это может показаться очень примитивным в отношении той информации, которой может оперировать человек. И поэтому мы поговорим о таком понятии как образ, и о том, как образуются образы на уровне нервных клеток или нейроэлементов.
Давайте рассмотрим механизм получения специализации нейронами на модели.
У нас имеется рецепторное поле из 12 рецепторов (Q, W, E, R…V), все рецепторы являются одинаковыми, находятся в равных условиях. И каждый рецептор имеет представительство (рецепторный нейрон) в группе взаимосвязанных клеток расположенных в плоскости, подобно тому, как это организованно во фрагменте коры обрабатывающей сигналы от органов чувств. Это будет область первичной обработки и поэтому нейроэлементы этой области имеют очень высокую пластичность (P = 1).
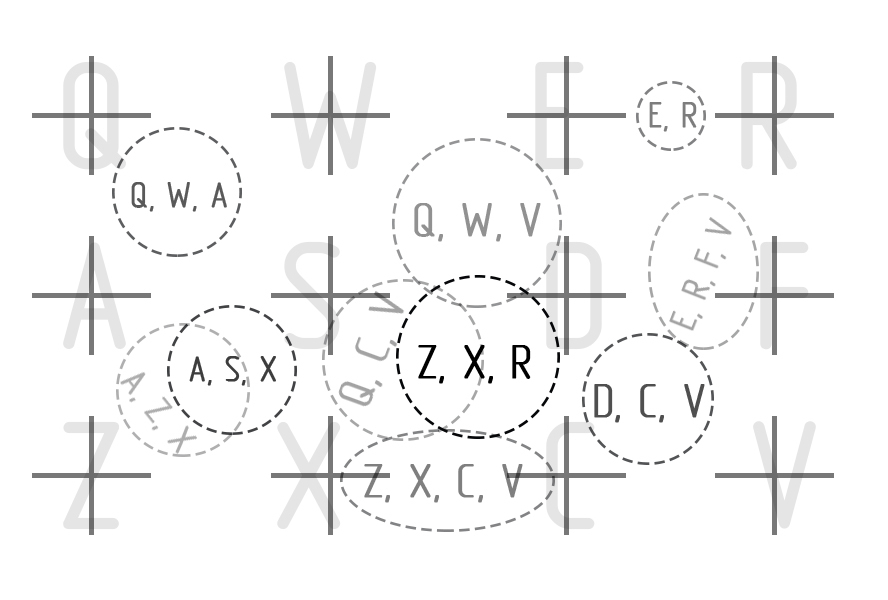
И конечно можно заметить, что эта система по формированию областей образов работает очень не точно. Области образов могут пересекаться, а так же возможно, что одной и той же области могут, привязаны совершенно разные группы раздражителей. И такая неточность в работе мозга может оказывать скорее положительный характер. Ведь в распознавании образов точность может только мешать, а так же это раскрывает потенциал к творчеству ведь совмещение ранее несовместимого или увидеть в образе нечто иное возможно только потому, что эти образы не так однозначны и не имеют четких границ. Еще это объясняет то, почему человеческое восприятие склонно к обманчивости.
Но для выделения этих областей повышенного уровня активирующего воздействия, необходимо добавить еще одну группу нейроэлементов которые будут активироваться только в случае получения такого повышенного воздействия. О структуре коры и организации коры мы подробно поговорим в посвященному этому выпуску, а так же о том как качественно улучшить механизм формирования образов.
Теперь давайте соединим иерархическую систему памяти и систему формирования образов.
Возбуждение от одного раздражителя, прежде чем попасть на следующий уровень обработки может пойти различными путями. Так путь, который выбирает возбуждение, определяется влиянием других раздражителей. Этот путь определяет, к какому представительству более высокого уровня поступит возбуждение и как дальше по иерархии. Учитывая разницу в пластичности на уровнях, то образы на высоких уровнях будут более стабильны, а именно если комплексный раздражитель будет активироваться не полным или зашумленным, то все равно к активности придет тот участок, который будет более типичен для данного комплексного раздражителя. Причем иерархия в некоторых случая не является строгой, некоторые сигналы от рецепторов могут дублироваться и перескакивать уровни.
Такая структура памяти объясняет причину высокой скорости извлечения информации, возбуждение от рецепторов просто проходит по всем слоям обработки, влияя друг на друга, эти сигналы определяют путь, по которому возбуждения придет к нужным группам нейронов. Здесь нет никакого перебора информации и нет операций сравнения, с какой либо эталонной информацией и т.д. Эволюция шла по пути скорости, а не точности, хотя точность восприятия достигалась путем увеличения рецепторных полей и особыми способами организации мозга.
LTP
Рассуждения о памяти были бы не полными, если мы не коснемся темы долговременной потенциации. Этот эффект связан с тем, что при воздействии на нервную клетку сильным активирующим действием происходит повышение её чувствительности на некоторое время, причем время этого изменения в клетке может длиться от нескольких минут до нескольких недель. Преимущественно этот эффект проявляется на крупных пирамидальных клетках гиппокампа, хотя может, наблюдаться и в других областях но в меньшей концентрации.
Под действием усиленного воздействия в клетке запускается определенный каскад химических реакций, который приводит к формированию постсинаптической мембране дополнительных рецепторов, что увеличивает чувствительность клетки. Давайте представим, как это будет реализовано в модели.
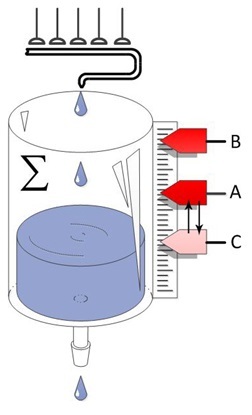
Такая особенность в работе некоторых клеток гиппокампа идет в разрез принципам привыкания в нервной системе, но такая особенность имеет больше значение. Благодаря ней организм может помнить о том, что с ним происходило несколько минут назад. Если Вы закроете глаза и некоторое время будете, побудете в тишине, то Вам не требуется даже открывать глаза, чтобы вернуть в памяти информацию о том, где Вы находитесь и какие события происходили ранее. Удивительное здесь то, что нет никаких раздражителей, которые будут активировать рефлекторные дуги, приводящие к нейронам которые ответственны за данную информацию.
В процессе деятельности в гиппокампе происходит как бы маркировка областей и нейронов посредством долговременной потенциации, маркируются те области, которые ответственны за активные образы. Благодаря чему можно легко вернуться к образам, которые были недавно активированы даже посредством более слабого воздействия.
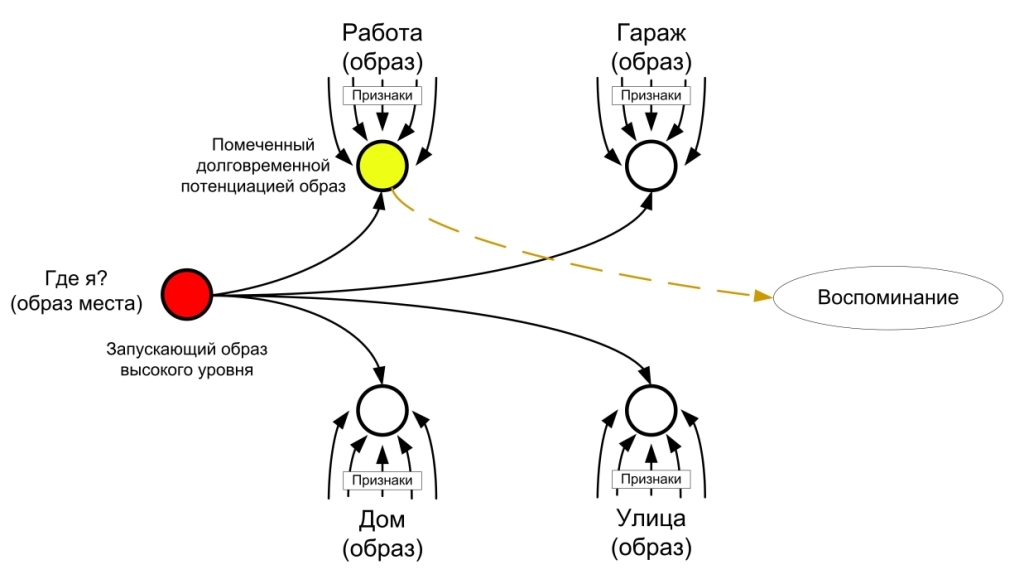
Долговременная потенциация позволяет помнить информацию которой организм оперировал недавно но при этом информация не была новой или значимой. А так же увеличивает время рабочей памяти.
Полученная теория памяти, на мой взгляд, очень проста и лаконична, причем она вытекает из основ, заложенных в работу нейроэлементов.
В следующем выпуске мы поговорим об эмоциях, и я надеюсь убедить Вас, что самые сложные в представлении человека эмоции будут доступны для электронных мозгов.
Читайте также:
