
Сомит хорда и нервная трубка
Органогенезы, заключающиеся в образовании отдельных органов, составляют основное содержание эмбрионального периода. Они продолжаются в личиночном и завершаются в ювенильном периоде. Органогенезы отличаются наиболее сложными и разнообразными морфогенетическими преобразованиями. Необходимой предпосылкой перехода к органогенезам является достижение зародышем стадии гаструлы, а именно формирование зародышевых листков. Занимая определенное положение друг по отношению к Другу, зародышевые листки, контактируя и взаимодействуя, обеспечивают такие взаимоотношения между различными клеточными группами, которые стимулируют их развитие в определенном направлении. Это так называемая эмбриональная индукция —важнейшее следствие взаимодействия между зародышевыми листками.
В ходе органогенезов изменяются форма, структура и химический состав клеток, обособляются клеточные группы, представляющие собой зачатки будущих органов. Постепенно развивается определенная форма органов, устанавливаются пространственные и функциональные связи между ними. Процессы морфогенеза сопровождаются дифференциацией тканей и клеток, а также избирательным и неравномерным ростом отдельных органов и частей организма. Обязательным условием органогенезов наряду с размножением, миграцией и сортировкой клеток является их избирательная гибель (см. разд. 8.2.4).
Самое начало органогенеза называют нейруляцией. Нейруляция охватывает процессы от появления первых признаков формирования нервной пластинки до замыкания ее в нервную трубку (рис. 7.9). Параллельно формируются хорда и вторичная кишка, а лежащая по бокам от хорды мезодерма расщепляется в краниокаудальном направлении на сегментированные парные структуры — сомиты.
Нервная система позвоночных, включая человека, отличается устойчивостью основного плана строения на протяжении всей эволюционной истории подтипа. В формировании нервной трубки у всех хордовых много общего. Вначале неспециализированная спинная эктодерма, отвечая на индукционное воздействие со стороны хордомезодермы, превращается в нервную пластинку, представленную нейроэпителиальными клетками цилиндрической формы.
Рис. 7.9. Последовательные стадии формирования нервной трубки и нервного гребня на 3-й неделе развития человеческого эмбриона (поперечный срез):
1—нервная пластинка, 2—нервный гребень, 3—эктодерма, 4—хорда, 5—нервная бороздка, 6—невроцель
Нервная пластинка недолго остается уплощенной. Вскоре ее боковые края приподнимаются, образуя нервные валики, которые лежат по обе стороны неглубокой продольной нервной бороздки. Края нервных валиков далее смыкаются, образуя замкнутую нервную трубку с каналом внутри — невроцелем. Раньше всего смыкание нервных валиков происходит на уровне начала спинного мозга, а затем распространяется в головном и хвостовом направлениях. Показано, что в морфогенезе нервной трубки большую роль играют микротрубочки и микрофиламенты нейроэпителиальных клеток. Разрушение этих клеточных структур колхицином и цитохалазином В приводит к тому, что нервная пластинка остается открытой. Несмыкание нервных валиков ведет к врожденным порокам развития нервной трубки.
После смыкания нервных валиков клетки, первоначально располагавшиеся между нервной пластинкой и будущей кожной эктодермой, образуют нервный гребень. Клетки нервного гребня отличаются способностью к обширным, но строго регулируемым миграциям по всему телу (см. разд. 8.2.2, рис. 8.1) и образуют два главных потока. Клетки одного из них—поверхностного—включаются в эпидермис или дерму кожи, где дифференцируются в пигментные клетки. Другой поток мигрирует в брюшном направлении, образует чувствительные спинномозговые ганглии, симпатические нервные узлы, мозговое вещество надпочечников, парасимпатические ганглии. Клетки из черепного отдела нервного гребня дают начало как нервным клеткам, так и ряду других структур, таких, как жаберные хрящи, некоторые кроющие кости черепа.
Мезодерма, занимающая место по бокам от хорды и распространяющаяся далее между кожной эктодермой и энтодермой вторичной кишки, подразделяется на дорсальную и вентральную области. Дорсальная часть сегментирована и представлена парными сомитами. Закладка сомитов идет от головного к хвостовому концу. Вентральная часть мезодермы, имеющая вид тонкого слоя клеток, называется боковой пластинкой. Сомиты соединены с боковой пластинкой промежуточной мезодермой в виде сегментированных ножек сомитов.
Все области мезодермы постепенно дифференцируются. В начале формирования сомиты имеют конфигурацию, характерную для эпителия с полостью внутри. Под индукционным воздействием, исходящим от хорды и нервной трубки, вентромедиальные части сомитов — склеротомы —превращаются во вторичную мезенхиму, выселяются из сомита и окружают хорду и вентральную часть нервной трубки. В конце концов из них образуются позвонки, ребра и лопатки.
Дорсолатеральная часть сомитов с внутренней стороны образует миотомы, из которых разовьются поперечно-полосатые скелетные мышцы тела и конечностей. Наружная дорсолатеральная часть сомитов образует дерматомы, которые дают начало внутреннему слою кожи — дерме. Из области ножек сомитов с зачатками нефротом и гонотом образуются органы выделения и половые железы.
Правая и левая несегментированные боковые пластинки расщепляются на два листка, ограничивающих вторичную полость тела — целом. Внутренний листок, прилежащий к энтодерме, называют висцеральным. Он окружает кишку со всех сторон и образует брыжейку, покрывает легочную паренхиму и мышцу сердца. Наружный листок боковой пластинки прилежит к эктодерме и называется париетальным. В дальнейшем он образует наружные листки брюшины, плевры и перикарда.
Энтодерма у всех зародышей в конечном счете образует эпителий вторичной кишки и многие ее производные. Сама вторичная кишка всегда располагается под хордой.
Таким образом, в процессе нейруляции возникает комплекс осевых органов нервная трубка — хорда — кишка, представляющих собой характернейшую черту организации тела всех хордовых. Одинаковое происхождение, развитие и взаимное расположение осевых органов выявляют их полную гомологию и эволюционную преемственность.
При углубленном рассмотрении и сравнении процессов нейруляции у конкретных представителей типа хордовых выявляются некоторые различия, которые связаны в основном с особенностями, зависящими от строения яйцеклеток, способа дробления и гаструляции (рис. 7.10). Обращают внимание отличающаяся форма зародышей и смещение времени закладки осевых органов друг относительно друга, т.е. описанная выше гетерохрония.
Рис. 7.10. Нейрулы различных хордовых животных.
А — ланцетник; Б —лягушка; В — цыпленок:
1—нервная трубка, 2—хорда, 3—сомит, 4—ножка сомита, 5—вторичная кишка, 6—боковая пластинка, 7—энтодерма
Эктодерма, мезодерма и энтодерма в ходе дальнейшего развития, взаимодействуя друг с другом, участвуют в формировании определенных органов. Возникновение зачатка органа связано с местными изменениями определенного участка соответствующего зародышевого листка. Так, из эктодермы развиваются эпидермис кожи и его производные (перо, волосы, ногти, кожные и молочные железы), компоненты органов зрения; слуха, обоняния, эпителий ротовой полости, эмаль зубов. Важнейшими эктодермальными производными являются нервная трубка, нервный гребень и образующиеся из них все нервные клетки.
Производными энтодермы являются эпителий желудка и кишки, клетки печени, секретирующие клетки поджелудочной, кишечных и желудочных желез. Передний отдел эмбриональной кишки образует эпителий легких и воздухоносных путей, а также секретирующие клетки передней и средней долей гипофиза, щитовидной и паращитовидной желез.
Мезодерма помимо уже описанных выше скелетных структур, скелетной мускулатуры, дермы кожи, органов выделительной и половой систем образует сердечно-сосудистую систему, лимфатическую систему, плевру, брюшину и перикард. Из мезенхимы, имеющей смешанное происхождение за счет клеток трех зародышевых листков, развиваются все виды соединительной ткани, гладкая мускулатура, кровь и лимфа (подробнее в разд. 8.2.5, схема 8.1).
Зачаток конкретного органа формируется первоначально из определенного зародышевого листка, но затем орган усложняется и в итоге в его формировании принимают участие два или три зародышевых листка.
Провизорные, или временные, органы образуются в эмбриогенезе ряда представителей позвоночных для обеспечения жизненно важных функций, таких, как дыхание, питание, выделение, движение и др. Недоразвитые органы самого зародыша еще не способны функционировать по назначению, хотя обязательно играют какую-то роль в системе развивающегося целостного организма. Как только зародыш достигает необходимой степени зрелости, когда большинство органов способны выполнять жизненно важные функции, временные органы рассасываются или отбрасываются.
Время образования провизорных органов зависит от того, какие запасы питательных веществ были накоплены в яйцеклетке и в каких условиях среды происходит развитие зародыша. У бесхвостых земноводных, например, благодаря достаточному количеству желтка в яйцеклетке и тому, что развитие идет в воде, зародыш осуществляет газообмен и выделяет продукты диссимиляции непосредственно через оболочки яйца и достигает стадии головастика. На этой стадии образуются провизорные органы дыхания (жабры), пищеварения и движения, приспособленные к водному образу жизни. Перечисленные личиночные органы дают возможность головастику продолжить развитие. По достижении состояния морфофункциональной зрелости органов взрослого типа временные органы исчезают в процессе метаморфоза.
Развитие нервной трубки (нейруляция) включает образование нервной пластинки, нервного желобка и замыкания последнего в нервную трубку.
Как только возникает нотохорд, эктодерма, расположенная над ним, начинает утолщаться и формировать нервную пластинку. Первым признаком дифференцировки нейроэктодермы является удлинение клеток этой области, и клетки приподнимаются над остальной частью эктодермы. Нервная пластинка занимает около 50% всей площади эктодермы. Материал нервной пластинки первоначально возникает вблизи гензеновского узелка, затем, когда головной отросток удлиняется, формирование нервной пластинки продолжается в краниальном направлении и, в конце концов, она достигает орофарингеальной мембраны. Приблизительно, на 18-е сутки нервная пластинка прогибается вдоль своей продольной оси и образует нервный желобок с возвышениями по обе стороны желобка — нервными валиками.
В конце 3-й недели в середине эмбриона нервный желобок превращается в нервную трубку и далее этот процесс распространяется в каудальном и краниальном направлениях. Однако в краниальной области смыкание желобка в трубку происходит более быстрыми темпами. К концу 4-й недели нервная трубка полностью сформирована. В головном конце, где будет формироваться головной мозг, нервная трубка широкая и стенка ее толстая, в ней контурируются вздутия и сужения, соответствующие будущим отделам головного мозга. В каудальном направлении трубка сохраняет цилиндрическую форму и постепенно суживается. Два открытых конца нервной трубки (спереди и сзади) именуются передним и задним нейропорами. В эмбриогенезе млекопитающих, благодаря наличию нейропоров, через нервную трубку некоторое время "протекает" амниотическая жидкость. Позже нейропоры смыкаются.
Как только нервные валики сливаются и образуют нервную трубку, нейроэктодермальные клетки (клетки нервного гребня), располагающиеся между нервной трубкой и эктодермальным эпителием мигрируют в стороны от нервной трубки и далее — по всему зародышу. Позднее эти клетки в разных участках эмбриона дадут начало нескольким клеточным дифферонам, в том числе пигментным клеткам, клеткам мозгового вещества надпочечников, клеткам периферической нервной системы.
Мезодерма, расположенная латерально от хорды, образует широкие полосы вдоль каждой стороны хорды и нервной трубки зародыша, именуемые несегментированной дорсальной мезодермой. В результате индуктивного влияния хорды и нервной трубки дорсальная часть мезодермы подвергается сегментации на сомиты. Первая пара сомитов (от греч. soma — тело) развивается на 20-е сутки в шейном отделе зародыша; последующие пары формируются в кранио-каудальном направлении (приблизительно, по три пары сомитов в сутки) вплоть до конца 5-й недели эмбриогенеза. В конечном итоге возникает от 42 до 44 пар сомитов (4 пары затылочных, 7 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 8-10 копчиковых сомитов). В последующем 1-я пара затылочных и 5-7-я пары копчиковых сомитов исчезают. В этот период развития число сомитов часто используется в качестве критерия для определения возраста зародыша.
Под влиянием веществ, вырабатываемых клетками хорды и нервной трубки (эмбриональная индукция), расположенные вентрально в составе сомитов клетки начинают митотически делиться, становятся полиморфными и распределяются вокруг хорды. В совокупности эти клетки называются склеротомом. В последующем стволовые клетки в составе склеротома дифференцируются в хондробласты и участвуют в образовании осевого скелета (позвонков, ребер, черепа и т. п.). После завершения миграции клеток склеротома оставшиеся клетки сомита формируют двухслойную трубку с наружным слоем — дер-матомом и внутренним — миотомом. Из дерматома в дальнейшем будут развиваться соединительные ткани кожи, из миотома — поперечнополосатая скелетная мышечная ткань.
Еще на конечной стадии гаструляции у амфибий в слое мезодермы в центральной сагиттальной плоскости начинает выделяться тяж клеток, видимый на поперечном срезе – хорда, которая образуется в результате конвергенции мезодермальных клеток. К концу нейруляции хорда значительно вытягивается в передне-заднем направлении и истончается.
Мезодермальная закладка дифференцируется на:
· сегментированную мезодерму, которая включает в себя дорсальные сегменты – сомиты и сегментные ножки (или нефротомы), расположенные на уровне 1-28 сомита;
· несегментированную мезодерму, представленную боковыми пластинками, или спланхнотомами, расположенными наиболее вентрально, и метанефрогенной тканью, лежащей каудальнее 28 пары сомитов.
Мезодерма будущих сомитов располагается латеральнее хорды. Мезодерма скапливается на дорсальной стороне в ходе вентродорсального движения клеток. Метамеризация мезодермы начинается с середины нейруляции: происходит последовательное разделение на отдельные парные сегменты (сомиты) и сегментные ножки.Процесс распространяется от головы к хвосту, примерно каждый час формируется новая пара сомитов. Клетки сомитов эпителизуются, сильно вытягиваются в поперечном направлении.
Головные сомиты отличаются от туловищных: они распадаются сразу после образования, однако следы метамеризации головы сохраняются в виде парных закладок жаберных дуг, нервов и т.д.
Туловищные сомиты в ходе дальнейшего развития разделяются на три зоны: дерматом — стенка сомита, прилежащая к эктодерме; склеротом — часть сомита, обращенная к хорде; миотом, лежащий между первыми двумя. Потомки клеток дерматома позже образуют дермальный слой кожи, склеротома — тела позвонков, миотома — поперечно-полосатую скелетную мускулатуру.
Потомки клеток сегментных ножек (нефротомов) и метанефрогенной ткани в дальнейшем примут участие в формировании органов выделительной системы.
Боковые пластинки (спланхнотомы) подразделяются на висцеральный (внутренний) и париетальный (наружный) листки, между которыми имеется щелевидная вторичная полость (целом). Спланхнотомы правой и левой стороны тела срастаются друг с другом под кишечной трубкой; целомическая полость становится общей для обеих сторон тела.
Висцеральный листок прирастает к кишке и другим внутренним органам и образует их серозную оболочку. Париетальный листок прирастает изнутри к стенке тела, образуя пристеночную брюшину.
При малом увеличении следует рассмотреть срез ранней нейрулы. Нужно определить границы всех трех зародышевых листков, найти гастроцель, нервную пластинку, мезодерму сомитов. При большом увеличении нужно обратить внимание на строение нервной пластинки и форму клеток в различных ее слоях, найти зачаток хорды и формирующиеся сомиты.
Клетки слоя мезодермы смещаются на дорсальную сторону. Выделяется мезодерма будущих сомитов, клетки которой сильно вытягиваются.
Хорошо различим тяж хорды, состоящей из плотно расположенных клеток с четко выраженными границами, цитоплазмой, содержащей небольшое количество пигментных гранул.
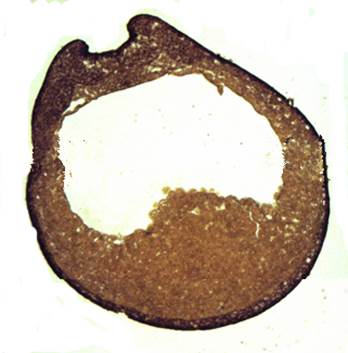
Рис. 14. Ранняя нейрула лягушки Начало формирования нервных валиков и нервного желобка. Фронтальный срез. Микропрепарат. Окр. по Ван-Гизону.
Под хордой видна замкнутая первичная кишка. Эксцентрично расположенная полость кишки имеет вид узкой щели. Ее нижняя стенка состоит из крупных клеток, цитоплазма которых богата желточными включениями. Часть клеток, прилежащих к просвету кишки, разрушена, что свидетельствует о процессе их ассимиляции. Клетки, содержащие трофический материал в виде желточных гранул, в своей совокупности составляют желточную энтодерму. Тонкая верхняя стенка кишки состоит из клеток кишечной энтодермы.
Боковые пластинки мезодермы представлены двумя слоями клеток — висцеральным и париетальным листками.
Полость бластоцеля в энтодерме на вентральной стороне зародыша редуцируется.
При малом увеличении нужно рассмотреть срез средней нейрулы. Определить границы всех трех зародышевых листков. Найти первичную кишку, нервную пластинку, дорсальную мезодерму сомитов, латеральную мезодерму боковых пластинок и закладку хорды. На большом увеличении нужно обратить внимание на строение нервной пластинки и форму клеток в различных ее слоях, высоту нервных валиков, форму клеток в формирующихся сомитах, а также различия в размерах клеток верхней и нижней стенок кишки.

Рис. 15. Средняя нейрула лягушки Сближение нервных валиков. Фронтальный срез. Микропрепарат. Окр. по Ван-Гизону.

Рис. 16. Поздняя нейрула лягушки Смыкание нервных валиков и образование нервной трубки с полостью внутри (невроцелем). Фронтальный срез. Микропрепарат. Окр. по Ван-Гизону.
При малом увеличении следует найти сформированную нервную трубку с невроцелем, первичную кишку, сомиты, листки спланхнотома и закладку хорды. На большом увеличении нужно рассмотреть клеточное строение всех перечисленных выше структур, найти скопление клеток нервного гребня между покровной эктодермой и нервной трубкой или чуть сбоку от нее.
Вопросы для самоконтроля:
1. Особенности гаструляции у рыб.
2. Последовательные этапы гаструляции у птиц. События, происходящие в гипо- и эпибласте.
3. Карты презумптивных участков зародышей рыб и птиц.
Препарат № 1. Первичная полоска зародыша курицы. Поперечный срез зародышевого щитка позади гензеновского узелка(рис. 17).
Гистологический срез, окраска гематоксилином и эозином.
Процесс гаструляции у птиц начинается еще в нижних отделах половых путей курицы и продолжается в процессе насиживания (инкубации).
В ходе первого этапа гаструляции бластодерма дискобластулы в результате миграции клеток в подзародышевую полость разделяется на два пласта: верхний многослойный (эпибласт) и нижний однослойный (первичный гипобласт). Между ними располагается щелевидная полость — бластоцель.
Зародышевый диск расположен на желтке не горизонтально — один конец всегда находится выше другого. Уже на самых ранних стадиях развития определяет положение передне-задней оси зародыша. Головной конец всегда располагается на том конце зародышевого диска, который расположен на желтке ниже. В гипобласте на заднем конце зародышевого диска в результате пролиферационной активности образуется клеточное скопление серповидной формы — так называемый серп Коллера. Клетки серпа Коллера продвигаются вперед, отодвигая клетки первичного гипобласта к краевой области зародышевого диска, — формируется вторичный гипобласт, который при дальнейшем развитии войдет в состав внезародышевой энтодермы.
Под действием ритмичного сокращения гипобласта начинаются гаструляционные движения в эпибласте куриного зародыша. В переднем конце эпибласта в результате пролиферации образуется клеточный материал, который начинает свое движение в направлении заднего конца эпибласта. При этом часть клеток перемещается по центральной линии (медленный поток), другие – по периферии (быстрый поток). Клетки периферического потока, достигая задний конец зародышевого щитка, концентрируется в светлой зоне и начинает движение назад единой полосой по центральной оси зародыша. Это скопление клеток получило название первичной полоски (рис. 17, а).
По мере продвижения первичной полоски в краниальном направлении происходит выселение части ее клеток в полость бластоцеля и расселяются между эпи- и гипобластом.
Миграция клеток из первичной полости очень активна, и клетки, продолжающие мигрировать в эпибласте в составе периферического потока к заднему концу зародыша, не могут восполнить их убыль. В результате по центральной оси первичной полоски появляется продольное углубление — первичная бороздка, которую отграничивают валики первичной полоски. Края первичной бороздки по своему значению соответствуют боковым губам бластопора.
Клетки первичной полоски дадут мезодерму сомитов, хвостовую и внезародышевую мезодерму.
На поперечном срезе в области первичной полоски (рис. 17, б) хорошо видны валики с углублением (первичной бороздкой) между ними область выселения мезодермальных клеток. Справа и слева от валиков располагаются плотный широкий клеточный слой - эпибласт. Самый нижний слой - гипобласт. Под верхним слоем размещается слой выселяющихся из первичной бороздки рыхлых мезодермальных клеток. По мере удаления от первичной полоски эпибласт истончается, приобретая вид двурядного, а затем и однослойного пласта кубических клеток. В мезодерме, ближайшей к первичной полоске, клетки расположены компактно, образуя густое скопление. В периферических участках меходермы клетки звездчатой или веретеновидной формы расположены рыхло, образуя синцитий. Гипобласт значительно тоньше эпиблиста, состоит из одного слоя уплощенных клеток; лишь на периферии слой утолщается, его клетки содержат желточные включения и приобретают неправильную звездчатую форму.
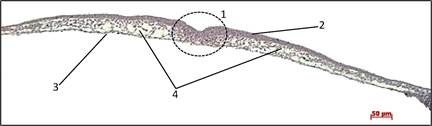
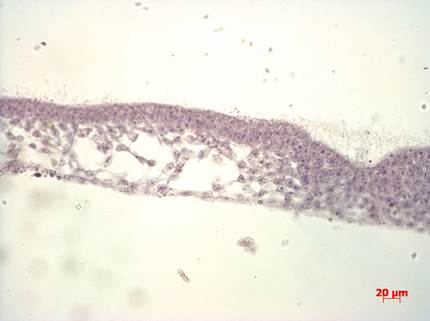
Рис. 17. Начальные стадии гаструляции у птиц. Фронтальный срез зародыша курицы в области первичной полоски. Микропрепарат. Окр. гематоксилином и эозином.
Для правильной ориентации срезов при просмотре гистологических препаратов куриных зародышей ранних стадий развития следует обращать внимание на расположение гранул желтка на краях среза: они должны быть направлены вниз. При такой ориентации спинная сторона зародыша будет наверху, а брюшная — внизу.
При малом увеличении микроскопа на стекле необходимо найти срез первичной полоски, имеющий вид тонкой изящной линии клеток, определить на срезе дорсальную и вентральную стороны (на периферии зародышевого диска с вентральной стороны видны гранулы желтка). Нужно найти на срезе область первичной полоски, и рассмотреть ее при большом увеличении. Обратить внимание на толщину эпи- и гипобласта, густое скопление мезодермальных клеток под первичной бороздкой и рыхлое их расположение между эпи- и гипобластом.
Препарат № 2. Сомиты, хорда и нервная трубка зародыша курицы. Поперечный срез зародыша впереди гензеновского узелка(рис. 18).
Гистологический срез, окраска гематоксилином и эозином.
Вторая стадия гаструляции характеризуется образованием структуры, состоящей примерно из 20 крупных клеток столбчатой формы — гензеновского узелка, являющимся результатом столкновения центрального потока клеток в эпибласте и первичной полоски.
Клетки, выселившиеся в полость бластоцеля через гензеновский узелок, мигрируют к переднему концу зародыша в виде сборной пластинки, в состав которой входит энтобласт (дает материал зародышевой энтодермы) и хордо-мезодермальный плащ (дает хордальный вырост и головную мезодерму). Клетки энтобласта присоединяются к вторичному гипобласту, формируя зародышевую (кишечную) энтодерму. В результате клетки гипобласта оказываются оттесненными на периферию зародышевого диска и участвуют только в формировании внезародышевой энтодермы (энтодермы желточного мешка). Клетки, выселяющиеся через боковые стороны первичной полоски, формируют латеральную мезодерму зародыша и внезародышевую мезодерму.
При сравнении процессов гаструляции у птиц и амфибий становится очевидным, что гензеновский узелок аналогичен дорсальной губе бластопора у амфибий, а стороны первичной полоски — боковым губам бластопора.
Одновременно с процессом гаструляции увеличивается диаметр зародышевого диска за счет разрастания края эпибласта по поверхности желтка (эпиболия).
При дальнейшем выселении клеток из первичной полоски ее материал расходуется и происходит регрессия первичной полоски. Длина ее уменьшается, и гензеновский узелок сдвигается в каудальном направлении до тех пор, пока весь клеточный материал первичной полоски не будет израсходован.
Сразу после выселения клеток сборной пластинки через гензеновский узелок начинаются конвергентные процессы, которые приводят к образованию хордального и мезодермальных тяжей, а вскоре после этого начинается дифференцировка мезодермы, распространяющаяся от передней части зародыша к его концу по мере перемещения гензеновского узелка.
Образующиеся хорда и мезодерма, инициируют начало формообразовательных процессов в нейроэктодерме. Это приводит к образованию нервной трубки по тому же механизму, что и у амфибий.
На поперечном срезе зародыша курицы впереди гензеновского узелка на поздней стадии гаструляции (рис. 17) под покровной эктодермой видна нервная трубка, имеющая форму овала с щелевидной полостью. Ниже, под нервной трубкой находится хорда и подстилающая ее кишечная энтодерма, над которой с обеих сторон хорды видны закладки будущих дуг аорты в виде двух тонкостенных полостей. По бокам от нервной трубки находятся сомиты, сегментные ножки (нефротомы) и листки спланхнотома. Иногда с дорсальной стороны сегментных ножек встречаются срезы полых трубочек – формирующихся органов выделения. Париетальный листок спланхнотома обращен к эктодерме, а висцеральный – к энтодерме. Между ними находится вторичная полость тела – целом.
В периферических частях препарата видны внезародышевая эктодерма, внезародышевые листки мезодермы (париетальный и висцеральный) и желточная (внезародышевая) энтодерма. Эти структуры в дальнейшем принимают участие в образовании туловищной и амниотической складок и стенок желточного мешка.
В толще висцерального листка мезодермы начинается ангиогенез, результатом которого являются имеющиеся здесь кровеносные сосуды, заполненные форменными элементами крови.
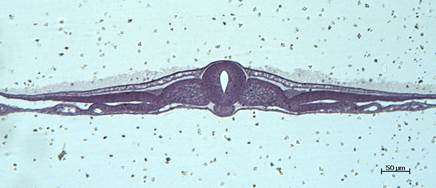
Рис. 18. Гаструляция у птиц. Фронтальный срез зародыша курицы впереди гензеновского узелка. Формирование нервной трубки и дифференцировка мезодермы. Микропрепарат. Окр. гематоксилином и эозином.
При малом увеличении микроскопа необходимо оритентировать препарат так, чтобы эктодерма и нервная трубка были обращены вверх. При большом увеличении следует рассмотреть особенности строения структур, представленных на препарате.
Содержание темы
1. а) В прошлой теме мы рассматривали начальные этапы эмбриогенеза - оплодотворение и дробление.
б) Второй из них завершается образованием бластулы .
2. В данной теме продолжим знакомство с ранними этапами эмбрионального развития.
6.1. Гаструляция
1. Как было определено (п. 5.2.1), гаструляция приводит к появлению в зародыше 3-х листков -
2. а) Далее из этих листков развиваются осевые зачатки органов.
6.1.1. Способы гаструляции
1. Гаструляция, в зависимости от вида животных, проходит различными способами.
2. Способ гаструляции во многом определяется типом бластулы, который, в свою очередь, обусловлен типом исходной яйцеклетки (п. 5.1.3.2).
6.1.1.1. Ланцетник: гаструляция путём инвагинации

6.1.1.2. Амфибии: гаструляция путём эпиболии
I. Общее описание
а) Инвагинация идёт на границе между анимальной и вегетативной частями бластулы.
б) Затем клетки анимального полюса (2) начинают энергично обрастать вегетативную часть (3) зародыша -
в) При этом серповидная бороздка всё углубляется:

II. Препарат

6.1.1.3. Птицы: гаструляция путём деламинации и иммиграции
I. Первый этап - деламинация
б) Из него-то (точнее, из его центральной части - зародышевого щитка) и образуются далее все 3 зародышевые листка -
II. Второй этап - иммиграция
в) Эти структуры - эквивалент бластопора амфибий,
т.к. представляют собой
III. Препарат
2. а) Первичная полоска (1) выглядит на поперечном срезе как утолщение в средней части зародыша;

6.1.1.4. Млекопитающие: гаструляция путём деламинации и иммиграции
I. Деламинация у млекопитающих
б) Но, кроме того, у высших животных и человека эпибласт расщепляется на два листка -
между которыми образуется полость амниона (4).
II. Иммиграция у млекопитающих
2. а) В этом месте внутрь проникают клетки (3), образующие энтодерму и мезодерму.
б) Остающиеся на поверхности клетки формируют эктодерму (4 ) .
6.1.2. Некоторые обобщения
6.1.2.1. Сводная таблица
Сведём воедино информацию о том, как тип яйцеклетки у животных определяет особенности их раннего эмбриогенеза.
однослойная, с полостью внутри
6.1.2.2. Дифференцировка - составная часть гаструляции
2. а) Дифференцировка проявляется в том, что
б) Например, клетки энтодермы далее могут превращаться только в эпителиальные клетки органов желудочно-кишечного тракта.
6.1.2.3. Факторы, вызывающие гаструляцию
6.2. Образование осевых зачатков органов
6.2.1. Производные зародышевых листков
1. Из материала трёх зародышевых листков (эктодермы, мезодермы, энтодермы) на следующем этапе эмбриогенеза формируются осевые зачатки органов.
2. Рассмотрим этот процесс на примере птиц.
6.2.1.1. Производные мезодермы
б) Формирующие её клетки мигрируют из эпибласта через первичный бугорок.
в ) А. Хорда - непарная осевая структура.
Б. Одна из её функций - установление оси тела .

б) Их ключевая особенность состоит в том, что они представляют собой сегменты - компактные скопления мезодермальных клеток.
в) А. Всего вдоль осевой линии образуется 10-11 пар сомитов (у человека - 44 пары) .
Б. Но появляются все пары сомитов не одновременно, а последовательно - в направлении от передней части зародыша к задней.
б) Расслаивается на 2 листка:
Б. Кроме того, висцеральный листок - зачаток миокарда и эпикарда.
б) Находится как внутри зародыша, так и вне его (внезародышевая мезенхима).
в) Из мезенхимы образуются
6.2.1.2. Производные эктодермы
I. Нервная трубка и ганглиозные пластинки
б) Является зачатком
6.2.1.3. Энтодерма
Б. Поэтому энтодерма называется кишечной .
а) Рассмотренная стадия эмбриогенеза часто в литературе называется нейруляцией , а сам зародыш - нейрулой .
Эти названия берут за основу один из важнейших процессов - образование нервной трубки.
б) Тем не менее, правильней использовать более общее название - " стадия образования осевых зачатков органов ".
6.2.2. Просмотр препаратов
6.2.2.1. Препараты зародыша курицы
I. Вид сверху
1. По оси зародыша - нервный желобок (1).
2. а) По краям от него зародыш значительно уплотнён -
б) А. Передняя часть мезодермы сегментирована:

II. Поперечный срез
1. Эктодерма (1 ) и её производная - нервная трубка (2) :

в) Также видны нефрогонотомы, или сегментные ножки (8) .
г) Хорошо развит спланхнотом , расщеплённый на
между которыми находится целомическая полость (6) .
6.2.2.2. Препараты зародышей амфибий
1. Гаструла амфибий (п. 6.1.1.2) заметно отличается от гаструлы птиц (п. 6.1.1.3) и млекопитающих (п. 6.1.1.4).
2. Тем не менее, и здесь образуются те же самые осевые зачатки органов.
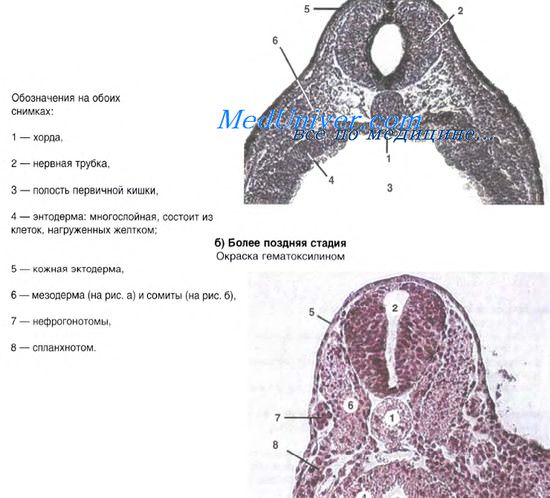
I. Относительно ранняя стадия гаструляции
б) Как видно, в месте расположения нервной трубки поверхность зародыша сильно приподнята.
2. а) В центре - обширная полость первичной кишки (3 ) .

II. Более поздняя стадия гаструляции
3. а) Остальные структуры имелись и прежде.
б) Это

6.3. Образование внезародышевых органов
и отделение от них тела зародыша
Внезародышевые органы иначе называются провизорными (временными).
Ещё одно название - зародышевые оболочки .
6.3.1. Внезародышевые органы у рыб и птиц:
схема образования
6.3.1.1. Желточный мешок
б) Затем внезародышевые части энтодермы (1) и висцерального листка мезодермы (2) начинают
б) Поэтому четыре листка :

6.3.1.2. Амнион и серозная оболочка
б) Одновременно происходит обрастание теми же листками внутренней поверхности скорлупы яйца . - Формируется серозная оболочка.
в) А. Данная оболочка отграничивает амниотическую полость,
в которой пространство между оболочкой и зародышем заполнено жидкостью.
б) В ней расположение листков обратное:
6.3.1.3. Аллантоис
б) Он образован теми же двумя листками ( внезародышевой локализации), что и желточный мешок -
6.3.2. Внезародышевые органы на препаратах
6.3.2.1. Зародыш рыб

б) Н а периферии зародыша к этим листкам присоединяются
6.3.2.2. Зародыши птиц
I. Образование складок
а) производные эктодермы -
б) производные мезодермы -

в) А. Двое из этих листков -
формируют стенку желточного мешка.
II. Смыкание складок
1. а) З десь вид на будущая презумптивная первичная кишка (1) .
б) Е ё стенка образована

3. С мыкание амниотических складок, состоящих из
6.3.3. Внезародышевые органы у млекопитающих
1. При эмбриональном развитии млекопитающих тоже образуются желточный мешок, амнион и аллантоис.
2. а) Вместо же серозной оболочки формируется хорион - ворсинчатая оболочка.
б) После внедрения хориона в слизистую оболочку матки с одной его стороны ворсинки сильно разрастаются и, глубоко проникая в слизистую, образуют вместе с ней новый орган - плаценту.
6.3.3.1. Способ образования
Амнион и желточный мешок образуются у высших млекопитающих и человека иначе, чем у птиц.
а) Он появляется в результате расщепления эпибласта на два листка -
б) Её клетки выселяются из зародышевого щитка в полость бластоцисты и постепенно обрастают все имеющиеся поверхности -
б) Он связан с хорионом с помощью амниотической ножки (8) из внезародышевой мезенхимы.
6.3.3.2. Состав стенок внезародышевых органов
Несмотрия на иной способ формирования, все перечисленные структуры имеют ту же природу, что и у птиц. -
1. а) Далее мы обратимся к строению основных типов тканей организма (общая гистология) и строению органов и система (частная гистология).
б) При этом будем касаться и эмбрионального происхождения соответствующих тканей и органов, что расширит приведённые в этом разделе сведения.
2. После чего (в последнем разделе курса) более детально рассмотрим вопросы раннего эмбриогенеза человека.
Читайте также:
