
Строение и регенерация периферического нерва
При разрушении участка периферического нерва в течение недели наступает восходящая дегенерация проксимальной (ближайшей к телу нейрона) части аксона с последующим некрозом как аксона, так и шванновской оболочки. На конце аксона формируется расширение (ретракционная колба). В дистальной части волокна после его перерезки отмечается нисходящая дегенерация с полным разрушением аксона, распадом миелина и последующим фагоцитозом детрита макрофагами и глией (рис. 1.5.7, 1.5.8).
Начало регенерации характеризуется сначала пролиферацией шванновских клеток, их передвижением вдоль распавшегося волокна с образованием клеточного тяжа, лежащего в эн-доневральных трубках. Таким образом, шван-новские клетки восстанавливают структурную целостность в месте разреза. Фибробласты также пролиферируют, но медленнее шванновских клеток. Указанный процесс пролиферации шванновских клеток сопровождается одновременной активацией макрофагов, которые первоначально захватывают, а затем лизируют оставшийся в результате разрушения нерва материал.
Следующий этап характеризуется прорастанием аксонов в щели, образованные шваннов-скими клетками, проталкиваясь от проксимального конца нерва к дистальному. При этом от ретракционной колбы в направлении дистальной части волокна начинают отрастать тонкие веточки (конусы роста). Регенерирующий аксон растет в дистальном направлении со скоростью 3—4 мм/сут вдоль лент из шванновских клеток (ленты Бюгнера), которые играют направляющую роль. В последующем наступает дифференциация шванновских клеток с образова-
Глава 1. КЛЕТКА И ТКАНИ
 |
| Дистальное Проксимальное > i |
| Место повреждения t t_________ tt___ ft |
J
| Ретроградная |
Уолеров- Тер- Транс-ская миналь- нейронная ная
Рис. 1.5.7. Термины, используемые при описании
различных типов дегенерации нейронов и нервных
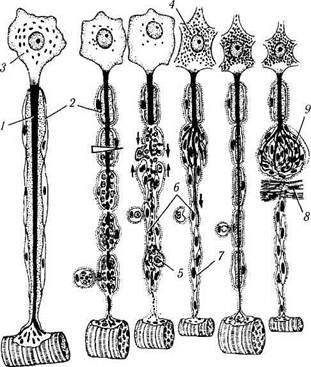
Рис. 1.5.8. Регенерация миелинового нервного волокна:
а — после перерезки нервного волокна проксимальная часть аксона (/) подвергается восходящей дегенерации, миелиновая оболочка (2) в области повреждения распадается, перикарион (3) нейрона набухает, ядро смещается к периферии, хромафильная субстанция (4) распадается; б—дистальная часть, связанная с иннервируемым органом, претерпевает нисходящую дегенерацию с полным разрушением аксона, распадом миелиновой оболочки и фагоцитозом детрита макрофагами (5) и глией; в — лем-моциты (6) сохраняются и митотически делятся, формируя тяжи — ленты Бюгнера (7), соединяющиеся с аналогичными образованиями в проксимальной части волокна (тонкие стрелки). Через 4—6 недель структура и функция нейрона восстанавливается, от проксимальной части аксона дистально отрастают тонкие веточки (жирная стрелка), растущие вдоль ленты Бюгнера; г — в результате регенерации нервного волокна восстанавливается связь с органом-мишенью и регрессирует ее атрофия; д — при возникновении преграды (8) на пути регенерирующего аксона компоненты нервного волокна формируют травматическую неврому (9), которая состоит из разрастающихся веточек аксона и леммоцитов
нием миелина и окружающей соединительной ткани. Коллатерали и терминали аксонов восстанавливаются в течение нескольких месяцев. Регенерация нервов происходит только при условии отсутствия повреждения тела нейрона, небольшом расстоянии между поврежденными концами нерва, отсутствии между ними соединительной ткани. При возникновении преграды на пути регенерирующего аксона развивается ампутационная нейрома. Регенерация нервных волокон в центральной нервной системе отсутствует.
Литература
1. Афанасьев Ю. И., Юрина Н. А. Гистология, ци
тология и эмбриология. — М.: Медицина, 1999. — 744 с.
2. Билич Г., Катинас Г. С, Назарова JJ. В. Цито
логия. — Спб., 1999.— 216 с.
3. Быков В. JI. Цитология и общая биология. —
Спб.: СОТИС, 1999.— 519 с.
4. Быков В. JI. Частная гистология человека. —
Спб.: СОТИС, 1997.— 300 с.
5. Дудел Д., Циммерман Л. Физиология чело
века: В 4 т. / Пер. с англ.; под ред. Р. Шмидта и
Г. Тевса. — Т. 2. — М.: Мир, 1985.— 240 с.
6. Луцик О. Д., 1ванова А.Й., Кабак К-С. Псто-
лопя людини.—Льв1в: Мир, 1992. — С. 399.
7. Леей А., Сикевиц И. Структура и функция клет
ки. — М.: Мир, 1971, —583 с.
8. Хэм А., Кормак Д. Гистология / Пер. с англ. —
М.: Мир, 1982,— 1350 с.
9. Елисеев В. Г. Гистология.—М.: Медицина,
1972.— С. 612.
10. Toda K-, Fitzpatrlck Т. В. The origin of melano-somes, in Kawamura T. (eds): Biology of normal and Abnormal melanocytes // Tokyo, University of Tokyo press. — 1971. — P. 265—267.
U.Szabo G., Gerald А. В., Pathak M. A. Racial differences in the fate of meanosomes in human epidermis // Nature. — 1969.— Vol. 222. — P. 1081— 1082.
12. Wolff K- Melanocyte-Keratinocyte interactions in
vivo: The fate of melanosomes // Yale J Biol Med. —
1973.—Vol. 46.— P. 384—396.
13. Ramirez F., Pereira L. The fibrillins // Int J Bio-
chem Cell Biol. — 1999. — Vol. 31. —P. 255—259.
14. Kielty С M., Shuttleworth С A. Fibrillin-contain-
ing microfibrils: structure and function in health and di
sease // Int J Biochem Cell Biol. — 1995. —Vol. 27. —
p. 747—760.
15. Sakai L. Y., Keene D. R., Engvall E. Fibrillin, a
new 350-kd glycoprotem, is a component of extracellu
lar microfibrils // J Cell Biol. — 1986. — Vol. 103.—
P. 2499.
16. Wright D. W., McDaniels С N., Swasdison S.
Immunisation with undenatured bovine zonular fibrils
results in monoclonal antibodies to fibrillin // Matrix
Biol. — 1994.— Vol. 14.— P. 41—49.
17'. Thurmond F.A., Trotter J. A. Morphology and biomechanics of the microfibrillar network of sea cucumber dermis // J Exp Biol. — 1996. — Vol. 199.— P. 1817—1828.
18. McConnell СМ., DeMont M.E., Wright G. M.
Microfibrils provide non-linear elastic behaviour in the
abdominal artery of the lobster Homarus americanus //
J. Physiol.—1997.—Vol. 499. — P. 513—526.
19. Rosenbloom J., Abrams W. R., Mecham B. Extra
cellular matrix 4: the elastic fibre // FASEB J. —
1993.— Vol. 7.— P. 1208—1218.
2\.Mayne R., Mayne P. R., Baker J. R. Fibrilllin-1 is the major protein present in bovine zonular fibrils // Invest Ophthalmol Vis Sci. — 1997. — Vol. 38.— P. 1399—1411.
22. Wheatley H.M., Traboulsi E.I., Flowers В. Е.
Immunohistochemical localization of fibrillin in human
ocular tissues // Arch Ophthalmol. — 1995. — Vol. 113.—
P. 103—109.
23. Walacce R. N., Streeten B. W., Hanna R. B. Ro
tary shadowing of elastic system microfibrils in the ocu
lar zonule, vitreous, and ligamentum nuchae // Curr
Eye Res. — 1991, — Vol. 10.— P. 99—108.
24. Wright D. W., Mayne R. Vitreous humor of
chicken contains two fibrillar systems: an analysis of
their structure // J. Ultra Mol. Struct. Res. — 1988. —
Vol. 100. — P. 224—234.
25. Bishop P., Ayad S., Reardon A. Type VI collagen
is present in human and bovine vitreous // Graefes
Arch. Clin. Exp. Ophthalmol. — 1996.— Vol. 234.—
P. 710—713.
26. Lee В., Godfrey M., Vitale E. Linkage of Mar-
fan syndrome and a phenotypically related disorder
to two different fibrillin genes // Nature.— 1991.—
Vol. 352. — P. 330—338.
27. Zhang H., Apfelroth S. D., Ни W. Structure and
expression of fibrillin-2, a novel microfibrillar compo
nent preferentially located in elastic matrices // J. Cell
Biol. — 1994.— Vol. 124.— P. 855—863.
28. Zhang H., Ни W., Ramirez F. Developmental
expression of genes suggests heterogeneity of extracel
lular microfibrils // J. Cell Biol. — 1995. — Vol. 129. —
P. 1165—1176.
29. Mir S., Wheatley H. M., Maumenee-Hussels I. E.
A comparative histologic study of the fibrillin microfibril
lar system in the lens capsule of normal subjects and
subjects with Marfan syndrome // Invest Ophthalmol
Vis Sci. — 1998. — Vol. 39. — P. 84—93.
30. Farnsworth P. N. В., Burke P. Three-dimensional architecture of the suspensory apparatus of the lens of the rhesus monkey // Exp. Eye Res. — 1977. — Vol. 25. — P. 563.
3\.Pessier A. P., Potter K-A. Ocular pathology in bovine Marfan syndrome with demonstration of altered fibrillin immunoreactivity in explanted ciliary body cells // Lab Invest. — 1996. — Vol. 75. — P. 87—95.
32. Kielty СМ., Davies 5., Phillips J. Marfan synd
rome expression and microfibrillar abnormalities in a
family with predominant ocular defects // J. Med. Ge
net. — 1994. — Vol. 31. —P. 1—5.
33. Izquierdo N. J., Traboulsi E., Enger C. Glauco
ma in the Marfan syndrome // Trans Am Ophthalmol
Soc. — 1992. — Vol. 90.— P. 111 — 122.
34. Izquierdo N. J., Traboulsi E. /., Enger C. Strabis
mus in the Marfan syndrome // Am. J. Ophthalmol. —
1994.—Vol. 117.— P. 632—635.
35. Allen R., Straatsma В., Apt L. Ocular manifesta
tions of the Marfan syndrome // Trans. Am. Acad. Oph
thalmol. Otolaryngol. — 1967. — Vol. 71.— P. 18—38.
36. Ramsay M.S., Fine B.S., Shields J.A. The Mar
fan syndrome. A histopathologic study of ocular findings
//Am. J. Ophthalmol.—1973. —Vol. 76. —P. 103—116.
37. Cross H.E., Jensen A. D. Ocular manifestations
in the Marfan syndrome and homocystinuria // Am J.
Ophthalmol. — 1973. —Vol. 75. — P. 405—419.
38. Freissler K., Kuchle M., Naumann G. О. Н. Spon
taneous dislocation of the lens in pseudoexfoliation
syndrome // Arch Ophthalmol. — 1995. — Vol. 113.—
P. 1095—11.
39. Schlutzer-Schrehardt U., Naumann G.O.H.
A histopathologic study of zonular instability in pseudo-
exfoliation syndrome // Am. J. Ophthalmol.— 1994. —
Vol. 118.— P. 730—743.
40. Schlutzer-Schrehardt U., Stumer J. P., Reme С. Е.
The fibrillin-containing microfibrillar network in the
trabecular meshwork of normal and glaucomatous eyes
// Invest Ophthalmol Vis Sci. — 1997. — Vol. 38.—
P. 2117—2126.
При повреждении нервов дегенеративные процессы аксона происходят дистальнее места поражения. Это связано с тем, что аксон является отростком нейрона, и его питание и рост зависят от сомы. Регенеративные процессы периферической нервной системы характеризуются высокой интенсивностью и в большинстве случае обеспечивают полное восстановление структур. В ЦНС, напротив, интенсивность процессов восстановления не высока, и регенерация не является полной.
а) Валлерова (антероградная) дегенерация периферических нервов. Основные этапы процесса дегенерации и их описание представлены на рисунке ниже и в подписи к нему. После разрушения или перерезки нерва в течение первых 48 ч аксоны и миелиновая оболочка под действием протеаз, высвобождающихся из шванновских клеток под влиянием ионов Са 2+ , распадаются на фрагменты элипсоидной формы дистальнее места нарушения целостности нерва. Продукты распада удаляют моноциты, которые перемещаются из кровотока в эндоневрий и превращаются в макрофаги.
Помимо осуществления фагоцитоза, макрофаги стимулируют митотическую активность шванновских клеток. Макрофаги и шванновские клетки выполняют трофическую (питательную) и направляющую функции для регенерирующих аксонов.
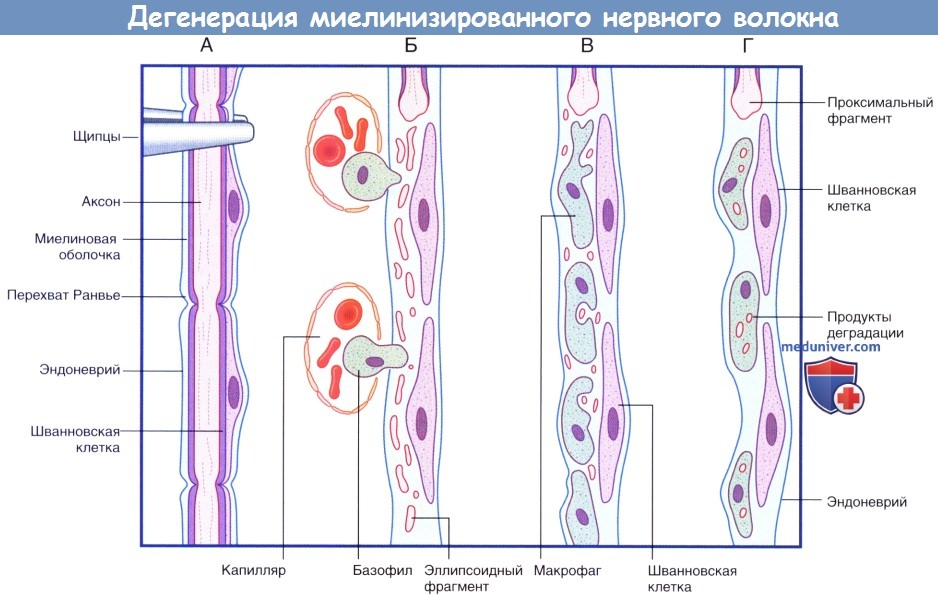
Последовательность процессов при дегенерации миелинизированного нервного волокна.
(А) Сохранное нервное волокно; продемонстрированы четыре сегмента. Выполняют пережатие нервного волокна в верхней части.
(Б) Миелиновая оболочка и аксон распадаются на мелкие частицы и фрагменты эллипсоидной формы. Моноциты проникают в эндоневрий из кровеносного русла.
(В) Мелкие продукты дегенерации поглощают моноциты.
(Г) Происходит практически полное удаление всех продуктов дегенерации. Шванновские клетки и эндоневрий сохранны. Последовательность процессов при регенерации миелинизированного нервного волокна. 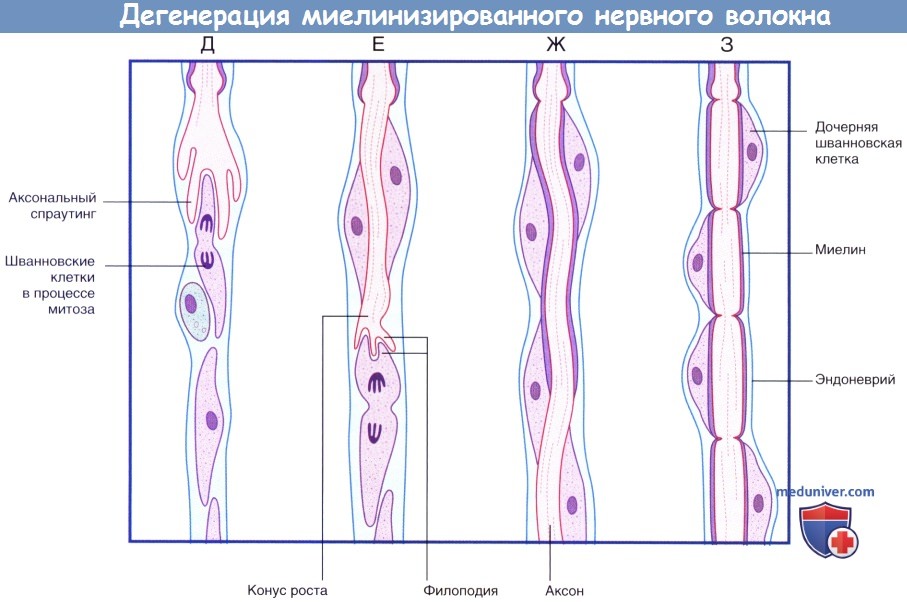
(продолжение) Последовательность процессов при дегенерации миелинизированного нервного волокна.
(Д) Аксональный спраутинг начинается с дистального участка аксона. Спраутинг оказывает митогенное дейсвтие на окружающие шванновские клетки.
(Е) Конус роста продвигается дистально вдоль поверхности шванновских клеток.
(Ж) Миелинизация начинается вдоль проксимального участка регенерирующего аксона.
(З) Общее строение вновь миелинизированного нервного волокна соответствует тем же принципам, однако миелинизированные сегменты характеризуются меньшей длиной.
б) Регенеративные процессы в периферической нервной системе. Основные этапы процесса регенерации представлены на рисунке ниже. После ровного среза нерва спраутинг (разрастание новых ветвей) на конце проксимального отрезка аксона начинается уже спустя несколько часов. Однако в клинической практике повреждения нерва часто происходят при раздавливании или разрыве. В этих случаях происходит отмирание участка нерва длиной 1 см и более, за счет чего спраутинг может продолжаться в течение недели. В случае удачной регенерации происходит тесное соприкосновение проксимального конца аксона со шванновской клеткой дистального конца пересеченного нейрона.
При нарушении формирования этой связи в месте первичного повреждения образуется псевдоневрома, представляющая собой извитые регенерирующие аксоны, погруженные в рубцовую ткань. Ампутационные псевдоневромы — источники сильных болей после ампутаций конечностей.
Регенерация нейронов при повреждении происходит двумя путями в течение нескольких часов после повреждения. На проксимальном конце пересеченного аксона появляются множественные отростки, на конце которых образуются утолщения — конусы роста. На дистальном конце шванновские клетки формируют отростки, направляющиеся навстречу конусам роста. На концах конусов роста формируются напоминающие антенны филоподии, где располагаются поверхностные рецепторы, временно связывающиеся с соответствующими поверхностными молекулами адгезии базальных мембран шванновских клеток. Актиновые филаменты филоподий прикрепляются к поверхностным рецепторам и относительно этих соединений осуществляют дальнейшее продвижение конусов роста.
Конусы роста стимулируют митотическую активность шванновских клеток. Шванновские клетки делятся и миелинизируют наиболее крупные аксоны.
При полном разрыве нервных стволов перед попыткой их восстановления, как правило, выжидают около трех недель, поскольку сразу после повреждения их соединительнотканные оболочки отечны, а в течение этого промежутка времени они становятся немного толще, что позволяет шовному материалу лучше закрепиться. Кроме того, обрезание нервов перед наложением швов приводит к развитию вторичной аксотомии проксимальной части пересеченного аксона. В ходе экспериментальных исследований на животных показано, что вторичная аксотомия стимулирует более интенсивную и длительную регенерацию.
Влияния тела нейрона на пересеченный участок нерва заключаются в следующем:
• Вследствие осмотических изменений перикариона ядро ориентируется эксцентрически.
• Клетки нейроглии заполняют все синаптические щели, изолируя исходный двигательный нейрон от синаптических контактов в сером веществе.
• В исследованиях, проведенных на обезьянах, показано, что после перерезки чувствительных нейронов 30-40 % окончаний дорсальных нервных корешков подвергается валлеровской дегенерации. Регенерация этих нейронов не происходит, поскольку их окончания располагаются в сером веществе головного мозга. Однако некоторые рецепторы восстанавливаются за счет спраутинга сохранных прилежащих нейронов. Данное наблюдение позволяет объяснить явление неполного восстановления чувствительности у таких пациентов.
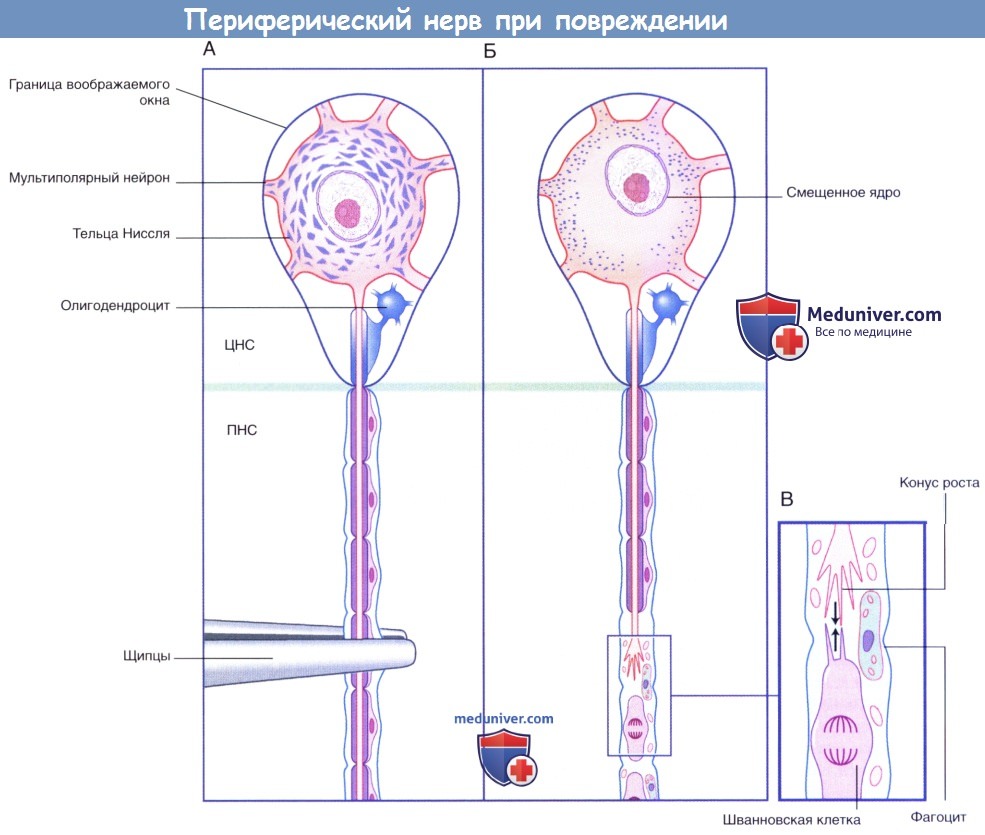
Схематическое изображение процессов, происходящих в периферическом нерве после повреждения.
(А) Двигательный нейрон ЦНС, видимый через воображаемое окно.
(Б) Хроматолизис проявляется фрагментированием и рассеиванием телец Ниссля, а также смещением ядра.
(В) В зоне повреждения в условиях удаления продуктов деградации происходит образование контакта филоподий конуса роста с проксимальными выростами шванновских клеток (указано стрелками).
ЦНС — центральная нервная система; ПНС — периферическая нервная система.
Редактор: Искандер Милевски. Дата публикации: 12.11.2018
Нервы (нервные стволы) – это структуры, которые связывают центры головного и спинного мозга с рецепторами и рабочими органами. Они состоят из миелиновых и безмиелиновых афферентных и эфферентных волокон и прослоек соединительной ткани. Могут быть отдельные нейроны и отдельные нервные ганглии. Прослойка рыхлой соединительной ткани, окружающая каждое нервное волокно, называется эндоневрий; окружающая пучок нервных волокон — периневрий, который состоит из 5-6 слоев коллагеновых волокон; между этими слоями имеются щелевидные полости, выстланные нейроэпителием, в которых циркулирует жидкость. Весь нерв окружен прослойкой соединительной ткани, которая называется эпиневрий. В периневрии и эпиневрии имеются кровеносные сосуды и нервы нервов.
Регенерация. При повреждении нерва разрываются проходящие в нем нервные волокна. После разрыва волокна в нем образуются 2 конца — конец, который связан с телом нейрона, называется центральным; конец, не связанный с нервной клеткой, называется периферическим.
В периферическом конце происходят 2 процесса: 1) дегенерация и 2) регенерация. Вначале идет процесс дегенерации, заключающийся в том, что начинается набухание нейролеммоцитов, растворяется миелиНовый слой, осевой цилиндр фрагментируется, образуются капли (овоиды), состоящие из миелина и фрагмента осевого цилиндра. К концу 2-й недели происходит рассасывание овоидов, остается только неврилемма оболочки волокна. Нейролеммоциты продолжают размножаться, из них образуются ленты (тяжи).
После рассасывания овоидов осевой цилиндр центрального конца утолщается и образуется колба роста, которая начинает расти, скользя по лентам нейролеммоцитов. К этому времени между разорванными концами нервных волокон образуется нейроглиально-соединительнотканный рубец, являющийся препятствием для продвижения колбы роста. Поэтому не все осевые цилиндры могут пройти на противоположную сторону образовавшегося рубца. Следовательно, после повреждения нервов иннервация органов или тканей полностью не восстанавливается. Между тем часть осевых цилиндров, оснащенных колбами роста, пробивается на противоположную сторону нейроглиального рубца, погружается в тяжи нейролеммоцитов. Затем мезаксон навертывается на эти осевые цилиндры, образуется миелиновый слой оболочки нервного волокна. В том месте, где находится нервное окончание, рост осевого цилиндра приостанавливается, формируются терминали окончания и все его компоненты.
95. Рефлекторная дуга вегетативного симпатического рефлекса
Преганглионарные нейроны симпатической нервной системы расположены в боковых ядрах спинного мозга, начиная с 8-го шейного сегмента и заканчиваясь 2-м поясничным сегментом включительно. В сегментах 8-го шейного, 1 и 2 грудного сегмента находятся нейроны, возбуждение которых вызывает расширение зрачка (сокращение дилататора зрачка), сокращение глазничной части круговой мышцы глаза, а также сокращение одной из мышц верхнего века.
От 1,2,3,4 и 5 грудных сегментов начинаются преганглионарные симпатические волокна, которые направляются к сердцу и бронхам.
I – Преганглионарные волокна,
II – вегетативные ганглии,
III – постганглионарные волокна и клетки-мишени,
IV– иннервируемые органы, в которых заложены клетки-мишени;
В отличие от парасимпатической нервной системы симпатическая иннервирует почти все органы: сердце, сосуды, бронхи, ГМК желудочно-кишечного тракта, ГМК мочеполовой системы, потовые железы, печень, мышцы зрачка, матку, ткани, в которых совершается липолиз, гликогенолиз, надпочечники, ряд других желез внутренней секреции.
симпатическая нервная система способствует значительному повышению работоспособности организма – под ее влиянием возрастает гликогенолиз, глюконеогенез, липолиз, усиливается деятельность сердечно-сосудистой системы, происходит перераспределение массы крови из областей, способных переносить гипоксию, в области, где наличие кислорода и энергетических источников является основой существования. Происходит улучшение вентиляции легких. Одновременно, при активации симпатической нервной системы имеет место торможение деятельности желудочно-кишечного тракта, мочевого пузыря.


Содержание:
- 1 Ганглии
- 2 Периферические нервы
- 3 Нервные окончания
- 4 Регенерация периферических нервов
В периферической нервной системе различают следующие компоненты:
Ганглии представляют собой скопление нейронов, формирующих в анатомическом смысле небольшие узелки различного размера, разбросанные в различных участках тела. Различают два типа ганглиев — цереброспинальные и вегетативные. Тела нейронов спинномозговых ганглиев, как правило, округлой формы и различного размера (от 15 до 150 мкм). Ядро располагается в центре клетки и содержит четкое круглое ядрышко (рис. 1.5.1).
Каждое тело нейрона отделено от окружающей соединительной ткани прослойкой уплощенных капсулярных клеток (амфицитов). Их можно отнести к клеткам глиальной системы. Проксимальный отросток каждой ганглиозной клетки в заднем корешке разделяется на две ветви. Одна из них вливается в спинномозговой нерв, в котором проходит к рецепторному окончанию. Вторая входит в задний корешок и достигает заднего столба серого вещества на той же стороне спинного мозга.
Ганглии вегетативной нервной системы по строению сходны с цереброспинальными ганглиями. Наиболее существенное отличие сводится к тому, что нейроны вегетативных ганглиев мультиполярны. В области глазницы обнаруживаются различные вегетативные ганглии, обеспечивающие иннервацию глазного яблока.
Периферические нервы являются четко определяемыми анатомическими образованиями и довольно прочны. Нервный ствол окутывается снаружи соединительнотканным футляром на всем протяжении. Этот наружный футляр называют эпинервием. Группы из нескольких пучков нервных волокон окружаются периневрием. От периневрия отделяются тяжи рыхлой волокнистой соединительной ткани, окружающие отдельные пучки нервных волокон. Это эндоневрий (рис. 1.5.2).
Периферические нервы обильно снабжены кровеносными сосудами.
Периферический нерв состоит из различного количества плотно упакованных нервных волокон, являющихся цитоплазматическими отростками нейронов. Каждое периферическое нервное волокно покрыто тонким слоем цитоплазмы — неврилеммой, или шванновской оболочкой. Шванновские клетки (леммоциты), участвующие в формировании этой оболочки, происходят из клеток нервного гребня.
В некоторых нервах между нервным волокном и шванновской клеткой располагается слой миелина. Первые называются миелинизированными, а вторые — немиелинизированными нервными волокнами.
Миелин (рис. 1.5.3)
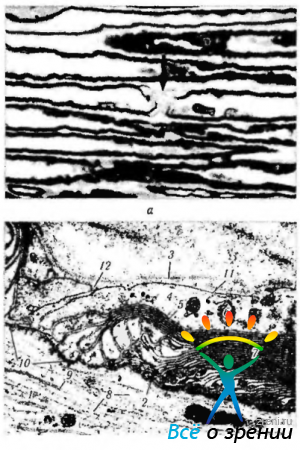
покрывает нервное волокно не сплошь, а через определенное расстояние прерывается. Участки прерывания миелина обозначаются перехватами Ранвье. Расстояние между последовательными перехватами Ранвье варьирует от 0,3 до 1,5 мм. Перехваты Ранвье имеются и в волокнах центральной нервной системы, где миелин образует олигодендроциты (см. выше). Нервные волокна разветвляются именно в перехватах Ранвье.
Каким образом формируется миелиновая оболочка периферических нервов? Первоначально шванновская клетка обхватывает аксон, так что он располагается в желобке. Затем эта клетка как бы наматывается на аксон. При этом участки цитоплазматической мембраны по краям желобка вступают в контакт друг с другом. Обе части цитоплазматической мембраны остаются соединенными, и тогда видно, что клетка продолжает обматывать аксон по спирали. Каждый виток на поперечном разрезе имеет вид кольца, состоящего из двух линий цитоплазматической мембраны. По мере наматывания цитоплазма шванновской клетки выдавливается в тело клетки.
Некоторые афферентные и вегетативные нервные волокна не имеют миелиновой оболочки. Тем не менее они защищены шванновскими клетками. Это происходит благодаря вдавливанию аксонов в тело шванновских клеток.
Механизм передачи нервного импульса в немиелинизированном волокне освещен в руководствах по физиологии. Здесь мы лишь кратко охарактеризуем основные закономерности процесса.
Известно, что цитоплазматическая мембрана нейрона поляризованна, т. е. между внутренней и наружной поверхностью мембраны существует электростатический потенциал, равный — 70 мВ. Причем внутренняя поверхность обладает отрицательным, а наружная положительным зарядом. Подобное состояние обеспечивается действием натрий-калиевого насоса и особенностями белкового состава внутрицитоплазматического содержимого (преобладание отрицательно заряженных белков). Поляризованное состояние называют потенциалом покоя.
При стимуляции клетки, т. е. нанесении раздражения цитоплазматической мембраны самыми разнообразными физическими, химическими и др. факторами, первоначально наступает деполяризация, а затем реполяризация мембраны. В физико-химическом смысле при этом наступает обратимое изменение в цитоплазме концентрации ионов К и Na. Процесс реполяризации активный с использованием энергетических запасов АТФ.
Волна деполяризации — реполяризации распространяется вдоль цитоплазматической мембраны (потенциал действия). Таким образом, передача нервного импульса есть не что иное, как распространяющаяся волна потенциала действия.
Каково же значение в передаче нервного импульса миелиновой оболочки? Выше указано, что миелин прерывается в перехватах Ранвье. Поскольку только в перехватах Ранвье цитоплазматическая мембрана нервного волокна контактирует с тканевой жидкостью, только в этих местах возможна деполяризация мембраны таким же образом, как в немиелинизированных волокнах. На остальном протяжении этот процесс невозможен в связи с изолирующими свойствами миелина. В результате этого между перехватами Ранвье (от одного участка возможной деполяризации до другого) передача нервного импульса осуществляется внутрицитоплазматическими местными токами. Поскольку электрический ток проходит гораздо быстрее, чем непрерывная волна деполяризации, передача нервного импульса в миелинизированном нервном волокне происходит значительно быстрее (в 50 раз), причем скорость увеличивается с увеличением диаметра нервного волокна, что обусловлено снижением внутреннего сопротивления. Подобный тип передачи нервного импульса называется сальтаторным. т. е. прыгающим. Исходя из изложенного, видно важное биологическое значение миелиновых оболочек.
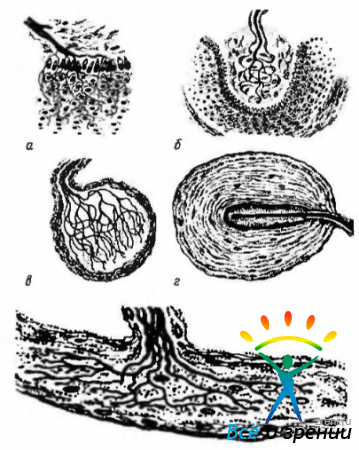
Афферентные (чувствительные) нервные окончания (рис. 1.5.5, 1.5.6).
Афферентные нервные окончания представляют собой концевые аппараты дендритов чувствительных нейронов, повсеместно располагающихся во всех органах человека и дающие информацию центральной нервной системе об их состоянии. Воспринимают они раздражения, исходящие и из внешней среды, преобразуя их в нервный импульс. Механизм возникновения нервного импульса характеризуется уже описанными явлениями поляризации и деполяризации цитоплазматической мембраны отростка нервной клетки.
Существует ряд классификаций афферентных окончаний — в зависимости от специфичности раздражения (хеморецепторы, барорецепторы, механорецепторы, терморецепторы и др.), от особенностей строения (свободные нервные окончания и несвободные).
Обонятельные, вкусовые, зрительные и слуховые рецепторы, а также рецепторы, воспринимающие движение частей тела относительно направления силы тяжести, называют специальными органами чувств. В последующих главах этой книги мы подробно остановимся только на зрительных рецепторах.
Рецепторы разнообразны по форме, строению и функциям. В данном разделе нашей задачей не является подробное описание различных рецепторов. Упомянем лишь о некоторых из них в разрезе описания основных принципов строения. При этом необходимо указать на различия свободных и несвободных нервных окончаний. Первые характеризуются тем, что они состоят только из ветвления осевых цилиндров нервного волокна и клетки глии. При этом они контактируют разветвлениями осевого цилиндра с клетками, возбуждающими их (рецепторы эпителиальных тканей). Несвободные нервные окончания отличаются тем, что в своем составе они содержат все компоненты нервного волокна. Если они покрыты соединительнотканной капсулой, они называются инкапсулированными (тельце Фатер—Пачини, осязательное тельце Мейснера, терморецепторы колбы Краузе, тельца Руффини и др.).
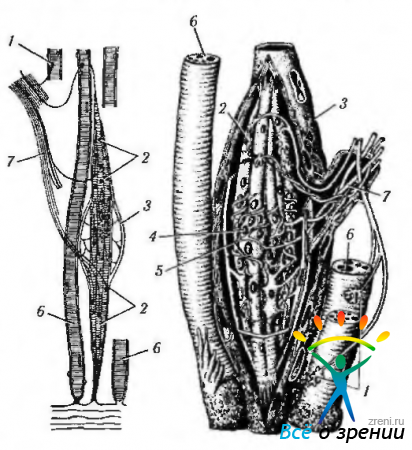
Разнообразно строение рецепторов мышечной ткани, часть которых обнаруживается в наружных мышцах глаза. В этой связи на них мы остановимся более подробно. Наиболее распространенным рецептором мышечной ткани является нервно-мышечное веретено (рис. 1.5.6). Это образование регистрирует растяжение волокон поперечно-полосатых мышц. Представляют они собой сложные инкапсулированные нервные окончания, обладающие как чувствительной, так и двигательной иннервацией. Число веретен в мышце зависит от ее функции и тем выше, чем более точными движениями она обладает. Нервно-мышечное веретено располагается вдоль мышечных волокон. Веретено покрыто тонкой соединительнотканной капсулой (продолжение периневрия), внутри которой находятся тонкие поперечнополосатые интрафузальные мышечные волокна двух видов:
-
волокна с ядерной сумкой — в расширенной центральной части которых содержатся скопления ядер (1—4— волокна/веретено);
Чувствительные нервные волокна образуют кольцеспиральные окончания на центральной части интрафузальных волокон обоих типов и гроздьевидные окончания у краев волокон с ядерной цепочкой.
Двигательные нервные волокна — тонкие, образуют мелкие нервно-мышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус.
Рецепторами растяжения мышцы являются также нервно-сухожильные веретена (сухожильные органы Гольджи). Это веретеновидные инкапсулированные структуры длиной около 0,5—1,0 мм. Располагаются они в области соединения волокон поперечнополосатых мышц с коллагеновыми волокнами сухожилий. Каждое веретено образовано капсулой из плоских фиброцитов (продолжение периневрия), которая охватывает группу сухожильных пучков, оплетенных многочисленными терминальными веточками нервных волокон, частично покрытых леммоцитами. Возбуждение рецепторов возникает при растяжении сухожилия во время мышечного сокращения.
Эфферентные нервные окончания несут информацию от центральной нервной системы к исполнительному органу. Это окончания нервных волокон на мышечных клетках, железах и др. Более подробное их описание будет приведено в соответствующих разделах. Здесь мы подробно остановимся лишь на нервно-мышечном синапсе (моторная бляшка). Моторная бляшка располагается на волокнах поперечнополосатых мышц. Состоит она из концевого ветвления аксона, образующего пресинаптическую часть, специализированного участка на мышечном волокне, соответствующего постсинаптической части, и разделяющей их синаптической щели. В крупных мышцах один аксон иннервирует большое количество мышечных волокон, а в небольших мышцах (наружные мышцы глаза) каждое мышечное волокно или их небольшая группа иннервируется одним аксоном. Один мотонейрон в совокупности с иннервируемыми им мышечными волокнами образует двигательную единицу.
Пресинаптическая часть формируется следующим образом. Вблизи мышечного волокна аксон утрачивает миелиновую оболочку и дает несколько веточек, которые сверху покрыты уплощенными леммоцитами и базальной мембраной, переходящей с мышечного волокна. В терминалах аксона имеются митохондрии и синаптические пузырьки, содержащие ацетилхолин.
Синаптическая щель имеет ширину 50 нм. Располагается она между плазмолеммой ветвлений аксона и мышечного волокна. Содержит она материал базальной мембраны и отростки глиальных клеток, разделяющих соседние активные зоны одного окончания.
Постсинаптическая часть представлена мембраной мышечного волокна (сарколеммой), образующей многочисленные складки (вторичные синаптические щели). Эти складки увеличивают общую площадь щели и заполнены материалом, являющимся продолжением базальной мембраны. В области нервно-мышечного окончания мышечное волокно не имеет исчерченности. содержит многочисленные митохондрии, цистерны шероховатого эндоплазматического ретикулума и скопление ядер.
Механизм передачи нервного импульса на мышечное волокно сходен с таковым в химическом межнейронном синапсе. При деполяризации пресинаптической мембраны происходит выделение ацетилхолина в синаптическую щель. Связывание ацетилхолина с холинорецепторами в постсинаптической мембране вызывает ее деполяризацию и последующее сокращение мышечного волокна. Медиатор отщепляется от рецептора и быстро разрушается ацетил-холинэстеразой.
При разрушении участка периферического нерва в течение недели наступает восходящая дегенерация проксимальной (ближайшей к телу нейрона) части аксона с последующим некрозом как аксона, так и шванновской оболочки. На конце аксона формируется расширение (ретракционная колба). В дистальной части волокна после его перерезки отмечается нисходящая дегенерация с полным разрушением аксона, распадом миелина и последующим фагоцитозом детрита макрофагами и глией (рис. 1.5.8).

Начало регенерации характеризуется сначала пролиферацией шванновских клеток, их передвижением вдоль распавшегося волокна с образованием клеточного тяжа, лежащего в эндоневральных трубках. Таким образом, шванновские клетки восстанавливают структурную целостность в месте разреза. Фибробласты также пролиферируют, но медленнее шванновских клеток. Указанный процесс пролиферации шванновских клеток сопровождается одновременной активацией макрофагов, которые первоначально захватывают, а затем лизируют оставшийся в результате разрушения нерва материал.
Следующий этап характеризуется прорастанием аксонов в щели, образованные шванновскими клетками, проталкиваясь от проксимального конца нерва к дистальному. При этом от ретракционной колбы в направлении дистальной части волокна начинают отрастать тонкие веточки (конусы роста). Регенерирующий аксон растет в дистальном направлении со скоростью 3—4 мм сут вдоль лент из шванновских клеток (ленты Бюгнера), которые играют направляющую роль. В последующем наступает дифференциация шванновских клеток с образованием миелина и окружающей соединительной ткани. Коллатерали и терминали аксонов восстанавливаются в течение нескольких месяцев. Регенерация нервов происходит только при условии отсутствия повреждения тела нейрона, небольшом расстоянии между поврежденными концами нерва, отсутствии между ними соединительной ткани. При возникновении преграды на пути регенерирующего аксона развивается ампутационная нейрома. Регенерация нервных волокон в центральной нервной системе отсутствует.
Читайте также:
- Центр нервно мышечной патологии человека с миастеническим центром
- Заразен ли ребенок после прививки от полиомиелита для привитых пинтаксимом
- Деятельность высокоорганизованной нервной системы животных егэ русский язык
- Как бороться с тремором рук в пожилом возрасте
- Что такое у кого впервые появились нервные клетки
