
Транспорт веществ в нервной клетке
Миллиарды нейронов формируют поверхностный слой — кору— полушарий головного мозга и полушарий мозжечка. Кроме того, в толще белого вещества нейроны образуют скопления—ядра.
Практически все нейроны ЦНС мультиполярны: сома (тело) нейронов характеризуется наличием нескольких полюсов (вершин). От каждого полюса, за исключением одного, отходят отростки — дендриты, которые образуют многочисленные разветвления. Дендритные стволы могут быть гладкими или образовывать многочисленные шипики. Дендриты образуют синапсы с другими нейронами в области шипиков или ствола дендритного дерева.
От оставшегося полюса сомы отходит отросток, проводящий нервные импульсы,— аксон. Большинство аксонов формирует коллатеральные ветви. Концевые ветви образуют синапсы с нейронами-мишенями.
Нейроны образуют два основных типа синаптических контактов: аксодендритические и аксосоматические. Аксодендритические синапсы в большинстве случаев передают возбуждающие импульсы, а аксосоматические — тормозящие.
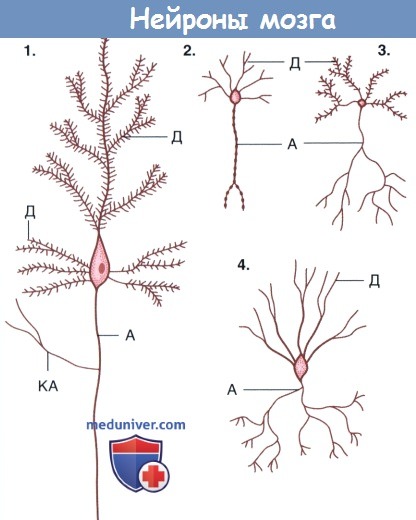
Формы нейронов мозга.
(1) Пирамидальные нейроны коры полушарий.
(2) Нейроэндокринные нейроны гипоталамуса.
(3) Шипиковые нейроны полосатого тела.
(4) Корзинчатые нейроны мозжечка. Дендриты нейронов 1 и 3 образуют шипики.
А — аксон; Д — дендрит; КА — коллатерали аксона. 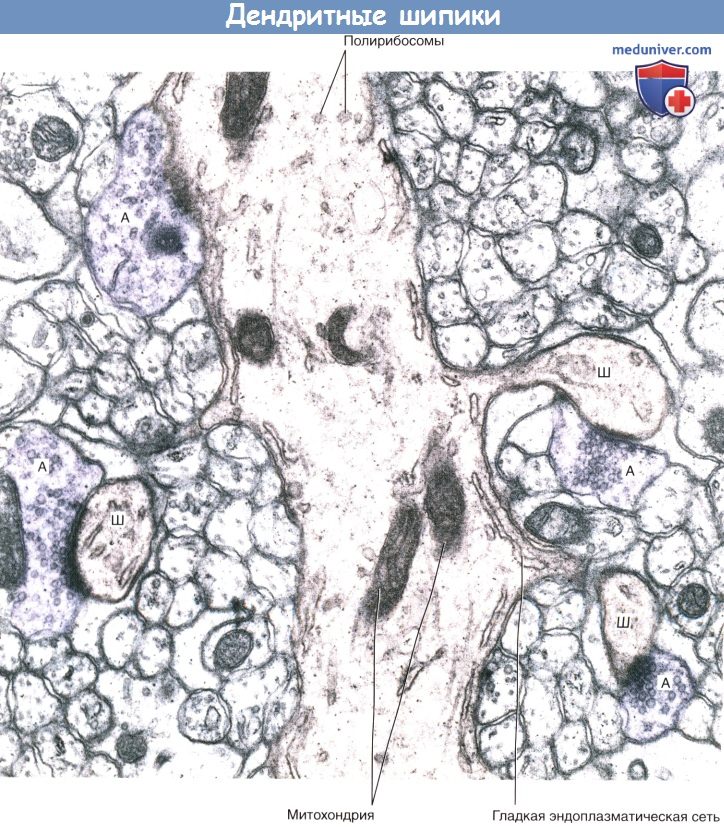
Дендритные шипики.
Срез мозжечка, на котором имеются дендриты гигантских клеток Пуркинье, образующие шипики.
В поле зрения различимы три шипика (Ш), образующие синаптические контакты с булавовидными расширениями аксонов (А).
Четвертый аксон (слева вверху) образует синапс с дендритным стволом. 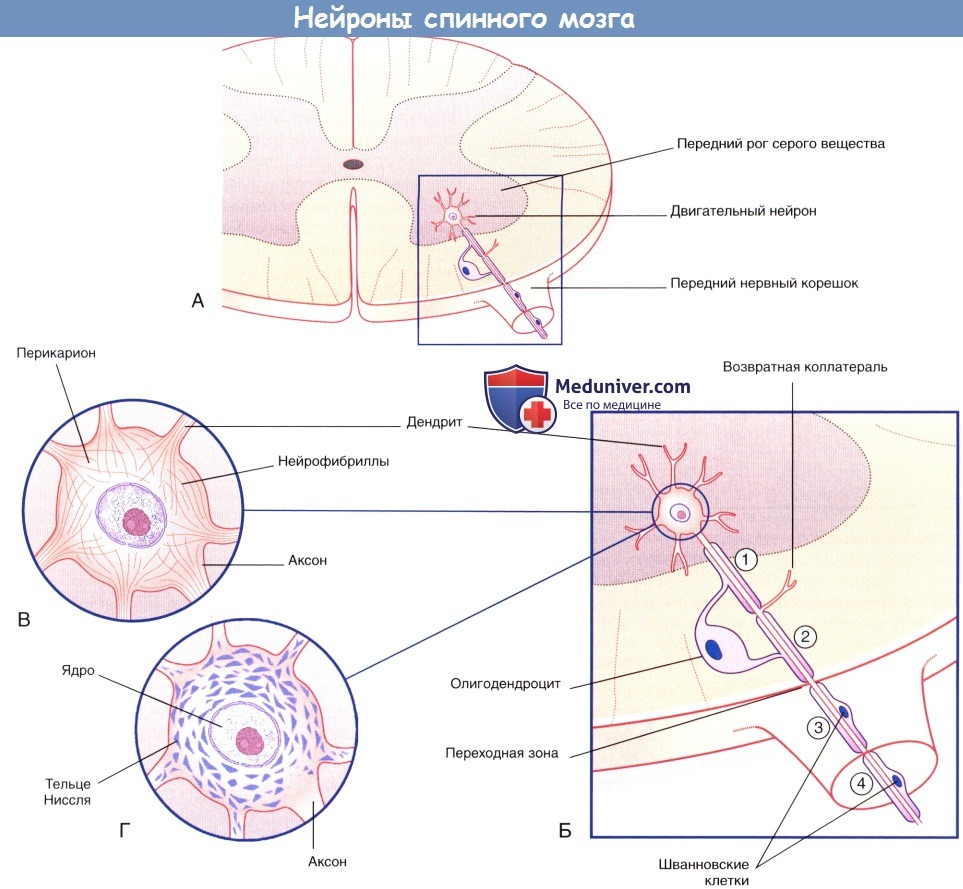
(А) Двигательный нейрон переднего рога серого вещества спинного мозга.
(Б) Увеличенное изображение (А). Миелиновые оболочки участков 1 и 2, располагающихся в белом веществе ЦНС, образованы олигодендроцитами.
Возвратная коллатеральная ветвь аксона начинается от немиелинизированного участка.
Миелиновые оболочки участков 3 и 4, относящихся к периферической части нервной системы, образованы шванновскими клетками.
Утолщение аксона в области вхождения в спинной мозг (переходного участка) соприкасается с одной стороны с олигодендроцитом, а с другой—со шванновской клеткой.
(В) Нейрофибриллы, состоящие из нейрофиламентов, видны после окрашивания солями серебра.
(Г) Тельца Ниссля (глыбки гранулярной эндоплазматической сети) видны при окрашивании катионными красителями (например, тионином).
Внутреннее строение нейронов
Цитоскелет всех структур нейрона образован микротрубочками и нейрофиламентами. Тело нейрона содержит ядро и окружающую его цитоплазму— перикарион (греч.peri— вокруг и karyon—ядро). В перикарионе расположены цистерны гранулярной (шероховатой) эндоплазматической сети — тельца Ниссля, а также комплекс Гольджи, свободные рибосомы, митохондрии и агранулярная (гладкая) эндоплазматическая сеть.
1. Внутриклеточный транспорт. В нейронах происходит обмен веществ между мембранными структурами и компонентами цитоскелета: непрерывно синтезируемые в соме новые клеточные компоненты перемещаются в аксоны и дендриты путем антероградного транспорта, а продукты метаболизма поступают путем ретроградного транспорта в сому, где происходит их лизосомальное разрушение (распознавание клеток-мишеней).
Выделяют быстрый и медленный антероградный транспорт. Быстрый транспорт (300-400 мм в сутки) осуществляют свободные клеточные элементы: синаптические пузырьки, медиаторы (или их предшественники), митохондрии, а также липидные и белковые молекулы (в том числе и белки-рецепторы), погруженные в плазматическую мембрану клетки. Медленный транспорт (5-10 мм в сутки) обеспечивают компоненты цнто-скелета и растворимые белки, в том числе и некоторые белки, задействованные в процессе высвобождения медиаторов в нервных окончаниях.
Первым среди нейротрофинов был изучен фактор роста нервов, выполняющий особенно важные функции в развитии периферической чувствительной и вегетативной нервной системы. В соме нейронов зрелого мозга синтезируется фактор роста, выделенный из головного мозга (BDNF), который транспортируется антероградно в их нервные окончания. Согласно данным, полученным в результате исследований на животных, фактор роста, выделенный из головного мозга, обеспечивает жизнедеятельность нейронов, принимая участие в обмене веществ, проведении импульсов и синаптической передаче.
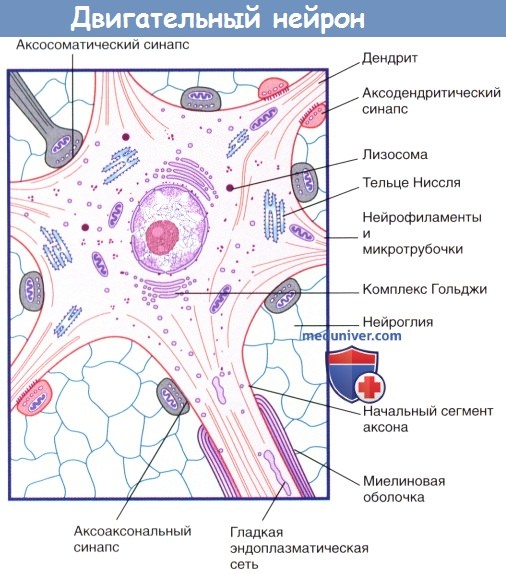
Внутреннее строение двигательного нейрона.
Изображены пять дендритных стволов, три возбуждающих синапса (выделены красным цветом) и пять тормозных синапсов.
2. Механизмы транспорта. В процессе нейронального транспорта роль поддерживающих структур выполняют микротрубочки. Связанные с микротрубочками белки перемещают органеллы и молекулы вдоль внешней поверхности миктротрубочек за счет энергии АТФ. Антероградный и ретроградный транспорт обеспечивают разные виды АТФаз. Ретроградный транспорт осуществляется за счет динеиновых АТФаз. Нарушение функционирования динеинов приводит к болезни двигательного нейрона.
Ниже описано клиническое значение нейронального транспорта.
Столбняк. При загрязнении раны почвой возможно заражение столбнячной палочкой (Clostridium tetani). Этот микроорганизм продуцирует токсин, который связывается с плазматическими мембранами нервных окончаний, проникает путем эндоцитоза в клетки и посредством ретроградного транспорта попадает в нейроны спинного мозга. Нейроны, расположенные на более высоких уровнях, также захватывают этот токсин путем эндоцитоза. Среди этих клеток необходимо особенно отметить клетки Реншоу, которые в норме оказывают тормозное действие на двигательные нейроны путем выделения тормозного медиатора—глицина.
При поглощении клетками токсина выделение глицина нарушается, вследствие чего прекращаются тормозные влияния на нейроны, осуществляющие двигательную иннервацию мышц лица, челюстей и позвоночника. Клинически это проявляется длительными и изнурительными спазмами этих мышц и в половине случаев заканчивается гибелью пациентов от истощения в течение нескольких дней. Предотвратить столбняк возможно, проведя своевременную иммунизацию в должном объеме.
Вирусы и токсичные металлы. Считают, что за счет ретроградного аксонального транспорта происходит распространение вирусов (например, вируса простого герпеса) из носоглотки в ЦНС, а также перенос токсичных металлов—алюминия и свинца. В частности, распространение вирусов по структурам мозга осуществляется за счет ретроградного межнейронального переноса.
Периферические нейропатии. Нарушение антероградного транспорта — одна из причин дистальных аксональных нейропатий, при которых развивается прогрессирующая атрофия дистальных участков длинных периферических нервов.
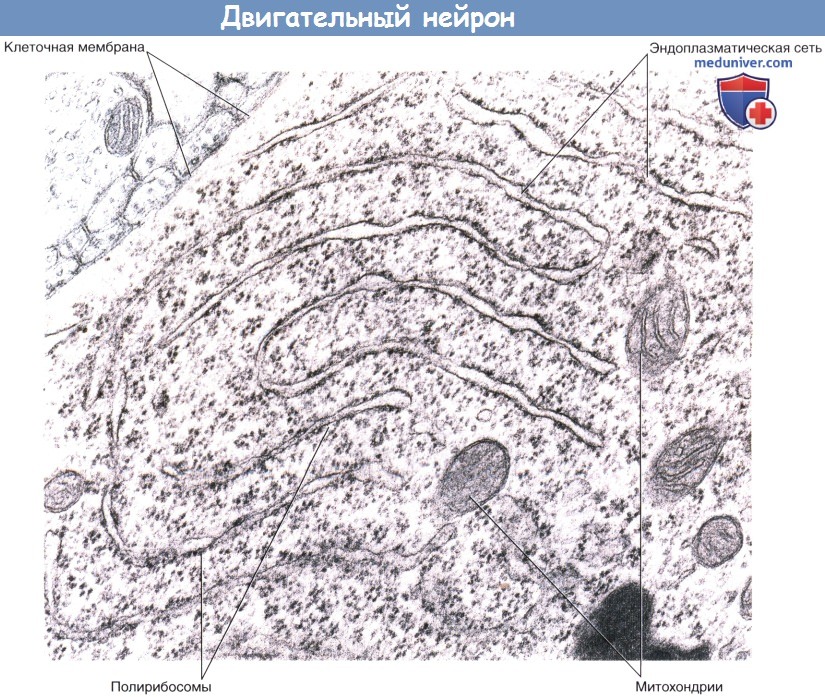
Тельце Ниссля в соме двигательного нейрона.
Эндоплазматическая сеть имеет многоуровневую структуру. Полирибосомы образуют выросты на внешних поверхностях цистерн или свободно лежат в цитоплазме.
(Примечание: для лучшей визуализации структуры слабо окрашены).
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
Для нейрона, имеющего многочисленные короткие отростки (дендриты) и длинный отросток (аксон), важным является транспорт веществ между отдельными участками нейрона и его отростками. Различают такие виды транспорта как аксонный (по направлению к его окончанию), дендритный (по направлению к окончаниям дендритов) и ретроградный транспорт, который направлен от отростков к перикариону. Аксонный транспорт подразделяется на два вида - медленный и быстрый транспорт. Рассмотрим эти все известные виды внутринейронального транспорта веществ.
Аксонный, или аксональный, транспорт (ток) представляет собой перемещение по аксону различных веществ и органелл. Он разделяется на антероградный - (прямой - из тела нейрона по аксону) и ретроградный (обратный - из аксона в тело нейрона). Вещества переносятся в цистернах агранулярной эндоплазматической сети и в пузырьках, которые перемещаются вдоль аксона благодаря взаимодействию с элементами цитоскелета (главным образом, с микротрубочкам) посредством связанных с ними сократительных белков - кинезина и динеина; процесс транспорта является Са2+-зависимым.
Быстрый аксонный транспорт совершается с относительно большой скоростью, достигающей 100-500 мм в сутки, а в отдельных случаях - даже 2000-2500 мм в сутки. С его участием переносятся различные питательные и биологически активные вещества, а также цистерны гранулярной эндоплазматической сети, митохондрии и пузырьки, содержащие медиаторы. Таким образом, быстрый аксонный транспорт, осуществляя жизнеобеспечение аксона, одновременно обеспечивает эффективность синаптической передачи. Характерно, что даже самый быстрый аксонный транспорт в миллионы раз медленнее скорости прохождения нервного импульса по нервному волокну. Следует отметить, что некоторые компоненты аксона, по-видимому, транспортируются с промежуточной скоростью. Дендритный транспорт заключается в движении веществ от тела к окончаниям дендритов. Известно, что таким способом транспортируются белки, необходимые для функционирования аксо-дендритических синапсов. В частности, транспортируется (со скоростью 3 мм/час.)ацетилхолинэстераза, которая разрушает ацетилхолин, выделяющийся в качестве медиатора в холинергических синапсах.
Ретроградные потоки веществ в нейроне осуществляют перенос веществ от аксона или дендритов к перикариону. За счет этих потоков могут транспортироваться некоторые цитоплазматические компоненты к телу клетки почти со скоростью быстрого аксонного транспорта. В частности, с участием этого вида транспорта к телу нейрона возвращаются многие цитоплазматические компоненты, что препятствует их накоплению в дендритах или аксонах. Высказано предположение, что именно благодаря ретроградному току нейрон контролирует целостность пре- и постсинаптических мембран в случае вредных воздействий на них. Ретроградный аксонный транспорт, который осуществляется со скоростью 100-200 мм в сутки, способствует удалению веществ из области терминалей, возвращению пузырьков, митохондрий.
Следует подчеркнуть, что наличие антероградных и ретроградных внутринейрональных потоков веществ может явиться причиной распространения патогенных микроорганизмов. Так, например, именно за счет аксонного транспорта проникшие в нейрон нейротропные вирусы (герпеса, бешенства, полиомиелита) могут распространяться по нейронным цепям.
В целом, феномен транспорта используется для изучения межнейронных связей путем введения маркера в область расположения терминалей или клеточных тел и выявления областей его последующего распространения описанными механизмами.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет





- Физиология
![]()
- История физиологии


Диффузия и транспорт веществ через клеточные мембраны
Обмен клетки с внешней средой различными веществами и энергией является жизненно необходимым условием ее существования.
Для поддержания постоянства химического состава и свойств цитоплазмы в условиях, когда имеют место существенные различия химического состава и свойств внешней среды и цитоплазмы клетки, должны существовать специальные транспортные механизмы, избирательно перемещающие вещества через клеточные мембраны.
В частности, клетки должны располагать механизмами доставки кислорода и питательных веществ из среды существования и удаления в нее метаболитов. Градиенты концентраций различных веществ существуют не только между клеткой и внешней средой, но и между органеллами клетки и цитоплазмой, и транспортные потоки веществ наблюдаются между различными отсеками клетки.
Особое значение для восприятия и передачи информационных сигналов имеет поддержание трансмембранной разности концентраций минеральных ионов Na + , К + , Са 2+ . Клетка затрачивает на поддержание концентрационных градиентов этих ионов существенную часть своей метаболической энергии. Запасаемая в ионных градиентах энергия электрохимических потенциалов обеспечивает постоянную готовность плазматической мембраны клетки отвечать на воздействие раздражителей. Поступление кальция в цитоплазму из межклеточной среды или из клеточных органелл обеспечивает ответ многих клеток на гормональные сигналы, контролирует выделение нейромедиаторов в синапсах, запускает сокращение мышц.

Рис. Классификация типов транспорта
Для понимания механизмов перехода веществ через клеточные мембраны необходимо учитывать как свойства этих веществ, так и свойства мембран. Транспортируемые вещества различаются молекулярной массой, переносимым зарядом, растворимостью в воде, липидах и рядом других свойств. Плазматическая и другие мембраны представлены обширными участками липидов, через которые легко диффундируют жирорастворимые неполярные вещества и не проходят вода и водорастворимые вещества полярной природы. Для трансмембранного перемещения этих веществ необходимо наличие специальных каналов в клеточных мембранах. Транспорт молекул полярных веществ затрудняется при увеличении их размеров и заряда (в этом случае требуются дополнительные механизмы переноса). Перенос веществ против концентрационных и других градиентов также требует участия специальных переносчиков и затрат энергии (рис. 1).

Рис. 1. Простая, облегченная диффузия и активный транспорт веществ через мембраны клеток
Для трансмембранного перемещения высокомолекулярных соединений, надмолекулярных частиц и компонентов клеток, не способных проникать через мембранные каналы, используются особые механизмы — фагоцитоз, пиноцитоз, экзоцитоз, перенос через межклеточные пространства. Таким образом, трансмембранное перемещение различных веществ может осуществляться с использованием разных способов, которые принято подразделять по признакам участия в них специальных переносчиков и энергозатратам. Существуют пассивный и активный транспорт через мембраны клетки.
Пассивный транспорт — перенос веществ через биомембрану по градиенту (концентрационный, осмотический, гидродинамический и т.д.) и без расхода энергии.
Активный транспорт — перенос веществ через биомембрану против градиента и с расходом энергии. У человека 30- 40 % всей энергии, образующейся в ходе метаболических реакций, расходуется на этот вид транспорта. В почках 70-80 % потребляемого кислорода идет на активный транспорт.

Под пассивным транспортом понимают перенос вещества через мембраны по различного рода градиентам (электрохимического потенциала, концентрации вещества, электрического поля, осмотического давления и др.), не требующий непосредственной затраты энергии на его осуществление. Пассивный транспорт веществ может происходить посредством простой и облегченной диффузии. Известно, что под диффузией понимают хаотические перемещения частиц вещества в различных средах, обусловленные энергией его тепловых колебаний.
Если молекула вещества электронейтральна, то направление диффузии этого вещества будет определяться лишь разностью (градиентом) концентраций вещества в средах, разделенных мембраной, например вне и внутри клетки или между ее отсеками. Если молекула, ионы вещества несут на себе электрический заряд, то на диффузию будут оказывать влияние как разность концентраций, величина заряда этого вещества, так и наличие и знак зарядов на обеих сторонах мембраны. Алгебраическая сумма сил концентрационного и электрического градиентов на мембране определяет величину электрохимического градиента.
Простая диффузия осуществляется за счет наличия градиентов концентрации определенного вещества, электрического заряда или осмотического давления между сторонами клеточной мембраны. Например, среднее содержание ионов Na+ в плазме крови составляет 140 мМ/л, а в эритроцитах — приблизительно в 12 раз меньше. Эта разность концентрации (градиент) создает движущую силу, которая обеспечивает переход натрия из плазмы в эритроциты. Однако скорость такого перехода малая, так как мембрана имеет очень низкую проницаемость для ионов Na + . Гораздо больше проницаемость этой мембраны для калия. На процессы простой диффузии не затрачивается энергия клеточного метаболизма.
Скорость простой диффузии описывается уравнением Фика:
dm/dt = -kSΔC/x,
где dm/dt - количество вещества, диффундирующее за единицу времени; к - коэффициент диффузии, характеризующий проницаемость мембраны для диффундирующего вещества; S - площадь поверхности диффузии; ΔС — разность концентраций вещества по обе стороны мембраны; х — расстояние между точками диффузии.
Из анализа уравнения диффузии ясно, что скорость простой диффузии прямо пропорциональна градиенту концентрации вещества между сторонами мембраны, проницаемости мембраны для данного вещества, площади поверхности диффузии.

Очевидно, что наиболее легко перемещаться через мембрану путем диффузии будут те вещества, диффузия которых осуществляется и по градиенту концентраций, и по градиенту электрического поля. Однако важным условием для диффузии веществ через мембраны являются физические свойства мембраны и, в частности, ее проницаемость для вещества. Например, ионы Na+, концентрация которого выше вне клетки, чем внутри ее, а внутренняя поверхность плазматической мембраны заряжена отрицательно, должны были бы легко диффундировать внутрь клетки. Однако скорость диффузии ионов Na+ через плазматическую мембрану клетки в покое ниже, чем ионов К+, который диффундирует по концентрационному градиенту из клетки, так как проницаемость мембраны в условиях покоя для ионов К+ выше, чем для ионов Na+.
Поскольку углеводородные радикалы фосфолипидов, формирующих бислой мембраны, обладают гидрофобными свойствами, то через мембрану могут легко диффундировать вещества гидрофобной природы, в частности легко растворимые в липидах (стероидные, тиреоидные гормоны, некоторые наркотические вещества и др.). Низкомолекулярные вещества гидрофильной природы, минеральные ионы диффундируют через пассивные ионные каналы мембран, формируемые каналообразующими белковыми молекулами, и, возможно, через дефекты упаковки в мембране фосфолииидных молекул, возникающие и исчезающие в мембране в результате тепловых флуктуаций.
Диффузия веществ в тканях может осуществляться не только через мембраны клеток, но и через другие морфологические структуры, например из слюны в дентинную ткань зуба через его эмаль. При этом условия для осуществления диффузии остаются теми же, что и через клеточные мембраны. Например, для диффузии кислорода, глюкозы, минеральных ионов из слюны в ткани зуба их концентрация в слюне должна превышать концентрацию в тканях зуба.
Облегченная диффузия — это вид пассивного переноса ионов через биологические мембраны, который осуществляется по градиенту концентрации с помощью переносчика.
Перенос вещества с помощью белка-переносчика (транспортера) основан на способности этой белковой молекулы встраиваться в мембрану, пронизывая ее и формируя каналы, заполненные водой. Переносчик может обратимо связываться с переносимым веществом и при этом обратимо изменять свою конформацию.
Предполагается, что белок-переносчик способен находиться в двух конформационных состояниях. Например, в состоянии а этот белок обладает сродством с переносимым веществом, его участки для связывания вещества повернуты внутрь и он формирует пору, открытую к одной из сторон мембраны.

Рис. 2. Облегченная диффузия. Описание в тексте
Связавшись с веществом, белок-переносчик изменяет свою конформацию и переходит в состояние 6. При этом конформационном превращении переносчик теряет сродство с переносимым веществом, оно высвобождается из связи с переносчиком и оказывается перемещенным в пору на другой стороне мембраны. После этого белок снова совершает возврат в состояние а. Такой перенос вещества белком-транспортером через мембрану называют унипортом.
Посредством облегченной диффузии могут транспортироваться такие низкомолекулярные вещества, как глюкоза, из интерстициальных пространств в клетки, из крови в мозг, реабсорбироваться некоторые аминокислоты и глюкоза из первичной мочи в кровь в почечных канальцах, всасываться из кишечника аминокислоты, моносахариды. Скорость транспорта веществ путем облегченной диффузии может достигать до 10 8 частиц за секунду через канал.
В отличие от скорости переноса вещества простой диффузией, которая прямо пропорциональна разности его концентраций по обе стороны мембраны, скорость переноса вещества при облегченной диффузии возрастает пропорционально увеличению разности концентраций вещества до некоторого максимального значения, выше которого она не увеличивается, несмотря на повышение разности концентраций вещества по обе стороны мембраны. Достижение максимальной скорости (насыщение) переноса в процессе облегченной диффузии объясняется тем, что при максимальной скорости в перенос оказываются вовлеченными все молекулы белков-переносчиков.
Обменная диффузия — при этом виде транспорта веществ может происходить обмен молекулами одного и того же вещества, находящимися по разные стороны мембраны. Концентрация вещества с каждой стороны мембраны остается при этом неизменной.
Разновидностью обменной диффузии является обмен молекулы одного вещества на одну или более молекул другого вещества. Например, в гладкомышечных клетках сосудов и бронхов, в сократительных миоцитах сердца одним из путей удаления ионов Са 2+ из клеток является обмен их на внеклеточные ионы Na+. На три иона входящего Na+ из клетки удаляется один ион Са 2+ . Создается взаимообусловленное (сопряженное) движение Na+ и Са 2+ через мембрану в противоположных направлениях (этот вид транспорта называют антипортом). Таким образом клетка освобождается от избыточного количества ионов Са 2+ , что является необходимым условием для расслабления гладких миоцитов или кардиомиоцитов.
Активный транспорт веществ через клеточные мембраны — это перенос веществ против их градиентов, осуществляющийся с затратой метаболической энергии. Этот вид транспорта отличается от пассивного тем, что перенос осуществляется не по градиенту, а против градиентов концентрации вещества и на него используется энергия АТФ или другие виды энергии, на создание которых АТФ затрачивалась ранее. Если непосредственным источником этой энергии является АТФ, то такой перенос называют первично-активным. Если на перенос используется энергия (концентрационных, химических, электрохимических градиентов), ранее запасенная за счет работы ионных насосов, затративших АТФ, то такой транспорт называют вторично-активным, а также сопряженным. Примером сопряженного, вторично-активного транспорта являются абсорбция глюкозы в кишечнике и ее реабсорбция в почках с участием ионов Na и переносчиков GLUT1.

Благодаря активному транспорту могут преодолеваться силы не только концентрационного, но и электрического, электрохимического и других градиентов вещества. В качестве примера работы первично-активного транспорта можно рассмотреть работу Na+ -, К+ -насоса.
Активный перенос ионов Na + и К+ обеспечивается белком- ферментом — Na+ -, К+ -АТФ-азой, способной расщеплять АТФ.
Белок Na К -АТФ-аза содержится в цитоплазматической мембране практически всех клеток организма, составляя 10% и более от общего содержания белка в клетке. На работу этого насоса тратится более 30% всей метаболической энергии клетки. Na + -, К+ -АТФ-аза может находиться в двух конформационных состояниях — S1 и S2. В состоянии S1 белок обладает сродством с ионом Na и 3 иона Na присоединяются к трем высокоаффинным местам его связывания, повернутым внутрь клетки. Присоединение иона Na" стимулирует АТФ-азную активность, и в результате гидролиза АТФ Na+ -, К+ -АТФ-аза фосфорилируется за счет переноса на нее фосфатной группы и осуществляет конформационный переход из состояния S1 в состояние S2 (рис. 3).
В результате изменения пространственной структуры белка места связывания ионов Na поворачиваются на внешнюю поверхность мембраны. Аффинность мест связывания к ионам Na+ резко уменьшается, и он, высвободившись из связи с белком, оказывается перенесенным во внеклеточное пространство. В конформационном состоянии S2 повышается аффинность центров Na+ -, К-АТФ-азы к ионам К и они присоединяют два иона К из внеклеточной среды. Присоединение ионов К вызывает дефосфорилирование белка и его обратный конформационный переход из состояния S2 в состояние S1. Вместе с поворотом центров связывания на внутреннюю поверхность мембраны два иона К высвобождаются из связи с переносчиком и оказываются перенесенными внутрь. Подобные циклы переноса повторяются со скоростью, достаточной для поддержания в покоящейся клетке неодинакового распределения ионов Na+ и К+ в клетке и межклеточной среде и, как следствие, поддержания относительно постоянной разности потенциалов на мембране возбудимых клеток.

Рис. 3. Схематическое представление работы Na+ -, К + -насоса
Вещество строфантин (оуабаин), выделяемое из растения наперстянка, обладает специфической способностью блокировать работу Na + -, К+ — насоса. После его введения в организм в результате блокады выкачивания иона Na+ из клетки наблюдаются снижение эффективности работы Na+ -, Са 2 -обменного механизма и накопление в сократительных кардиомиоцитах ионов Са 2+ . Это ведет к усилению сокращения миокарда. Препарат применяется для лечения недостаточности насосной функции сердца.
Кроме Na'-, К + -АТФ-азы имеются еще несколько типов транспортных АТФ-аз, или ионных насосов. Среди них насос, осуществляющий транспорт прогонов водорода (митохондрии клеток, эпителий почечных канальцев, париетальные клетки желудка); кальциевые насосы (пейсмекерные и сократительные клетки сердца, мышечные клетки поперечно-полосатой и гладкой мускулатуры). Например, в клетках скелетных мышц и миокарда белок Са 2+ -АТФ-аза встроен в мембраны саркоплазматического ретикулума и благодаря его работе обеспечивается поддержание высокой концентрации ионов Са 2+ в его внутриклеточных хранилищах (цистерны, продольные трубочки саркоплазматического ретикулума).
В некоторых клетках силы трансмембранной разности электрических потенциалов и градиента концентрации натрия, возникающие в результате работы Na+-, Са 2+ -насоса, используются для осуществления вторично-активных видов переноса веществ через клеточную мембрану.
Вторично-активный транспорт характеризуется тем, что перенос вещества через мембрану осуществляется за счет градиента концентрации другого вещества, который был создан механизмом активного транспорта с затратой энергии АТФ. Различают две разновидности вторично активного транспорта: симпорт и антипорт.
Симпортом называют перенос вещества, который сопряжен с одновременным переносом другого вещества в том же направлении. Симпортным механизмом переносятся йод из внеклеточного пространства в тиреоциты щитовидной железы, глюкоза и аминокислоты при их всасывании из тонкой кишки в энтероциты.
Антипортом называют перенос вещества, который сопряжен с одновременным переносом другого вещества, но в обратном направлении. Примером антипортного механизма переноса является работа упоминавшегося ранее Na + -, Са 2+ — обменника в кардиомиоцитах, К+ -, Н+ -обменного механизма в эпителии почечных канальцев.
Из приведенных примеров видно, что вторично-активный транспорт осуществляется за счет использования сил градиента ионов Na+ или ионов К+. Ион Na+ или ион К перемещается через мембрану в сторону его меньшей концентрации и тянет за собой другое вещество. При этом обычно используется встроенный в мембрану специфический белок-переносчик. Например, транспорт аминокислот и глюкозы при их всасывании из тонкого кишечника в кровь происходит благодаря тому, что белок-переносчик мембраны эпителия кишечной стенки связывается с аминокислотой (глюкозой) и ионом Na+ и только тогда изменяет свое положение в мембране таким образом, что переносит аминокислоту (глюкозу) и ион Na+ в цитоплазму. Для осуществления такого транспорта необходимо, чтобы снаружи клетки концентрация иона Na+ была гораздо больше, чем внутри, что обеспечивается постоянной работой Na+, К+ — АТФ-азы и затратой метаболической энергии.
Читайте также:
