
Вегетативная нервная система вики
Содержание
В соматической нервной системе нервные волокна тянутся к скелетной мышце, коже и органам чувств, а также от них. Соматические нервные волокна обычно реагируют на стимулы из окружающей среды, как, например, в случае рефлекса отдергивания. Большая часть активности соматической нервной системы происходит сознательно и под волевым контролем. Напротив, вегетативная, или автономная, нервная система (ВНС) в основном занята регуляцией кровообращения и деятельности внутренних органов. Она отвечает на изменяющиеся условия среды ортостатической реакцией, запуском реакций с целью регуляции внутренней среды организма. Основные виды деятельности вегетативной нервной системы не подлежат волевому контролю.
Вегетативная и соматическая нервные системы, в основном на периферии, разделены анатомически и функционально (А), но тесно контактируют в центральной нервной системе (ЦНС). Периферическая вегетативная нервная система является эфферентной (двигательной), но большинство вегетативных нервов несут также афферентные (сенсорные) нейроны. Они называются также висцеральными афферентными нейронами, потому что проводят сигналы от висцеральных (внутренних) органов, таких как пищевод, желудочно-кишечный тракт, печень, легкие, сердце, артерии и мочевой пузырь. Некоторые из них называются по нерву, который они сопровождают, например афферентный нейрон блуждающего нерва.
Активность вегетативной нервной системы обычно регулируется рефлекторной дугой, которая имеет афферентный сегмент (висцеральные и/или соматические афферентные нейроны) и эфферентный сегмент (автономные и/или соматические афферентные нейроны). Афферентные волокна проводят стимулы от кожи (болевые, или ноцицептивные, стимулы) и болевых рецепторов, от механорецепторов и хеморецепторов в таких органах, как легкие, желудочно-кишечный тракт, желчный пузырь, сосудистая система и гениталии. Вегетативная система содержит автономные эфферентные волокна, которые проводят рефлекторный ответ на афферентную информацию, индуцируя сокращение гладких мышц в таких органах, как глаза, легкие, пищеварительный тракт и желчный пузырь, и влияя на работу сердца и желез. Примеры действия соматической нервной системы - афферентные стимулы от кожи и органов чувств (например, реакция на световые стимулы) и эфферентные импульсы к скелетной мускулатуре (например, кашель и рвота).
Простые рефлексы могут быть внутри органа (например, в желудке), но сложные (комплексные) рефлексы идут через контролирующие вегетативные центры в ЦНС, в основном в спинном мозге (А). Эти центры контролируются гипоталамусом, активность которого связана с вегетативной нервной системой. Кора головного мозга -самый высокоорганизованный нервный центр, связывающий вегетативную нервную систему с другими системами.
Периферическая ВНС состоит из симпатического и парасимпатического отделов (А), которые в большинстве случаев действуют независимо. Вегетативные центры симпатического отдела расположены в грудном и поясничном отделах спинного мозга, а центры парасимпатического отдела - в стволовом отделе головного мозга (глаза, железы и органы, иннервируемые блуждающим нервом) и в крестцовом отделе спинного мозга (мочевой пузырь, нижняя часть толстой кишки и половые органы) (А). Преганглионарные волокна обоих отделов вегетативной нервной системы тянутся от центров к ганглиям, где заканчиваются на постганглионарных нейронах.
Преганглионарные симпатические нейроны, начинающиеся в спинном мозге, заканчиваются или в околопозвоночной ганглионарной цепи, в шейном или брюшном ганглии, или в так называемых терминальных ганглиях. Передача стимула от преганглионарных к постганглионарным нейронам является холинергической, т. е. опосредована высвобождением нейромедиатора ацетилхолина. Стимуляция постганглионарными симпатическими волокнами всех эффекторных органов, кроме потовых желез, является адренергической, т. е. опосредована высвобождением норадреналина.
Парасимпатические ганглии расположены вблизи или внутри эффекторных органов. Синаптическая передача в парасимпатических ганглиях и эффекторных органах является холинергической (А).
Большинство органов иннервируется симпатическими и парасимпатическими нервными волокнами. Однако ответы органов на активность этих двух систем могут быть или антагонистическими (как в сердце), или комплементарными (в половых органах).
Мозговое вещество надпочечников - это и ганглий, и эндокринная железа одновременно. Преганглионарные симпатические волокна в мозговом веществе надпочечников высвобождают ацетил-холин, что ведет к выделению в кровоток адреналина (и некоторого количества норадреналина).
Ацетилхолин (АЛ) служит нейромедиатором не только двигательной концевой пластинки и в центральной нервной системе, но и в вегетативной нервной системе, ВНС , где проявляет активность
- во всех преганглионарных волокнах ВНС
- во всех парасимпатических постганглионарных нервных окончаниях
- в некоторых симпатических постганглионарных нервных окончаниях (потовые железы).
Синтез ацетилхолина. Ацетилхолин (АХ) синтезируется в цитоплазме нервных окончаний, а ацетилкофермент А (ацетил-КоА) - в митохондриях. Реакция ацетил-КоА + холин катализируется холинацетилтрансферазой, которая синтезируется в соме и достигает нервных окончаний при помощи аксоплазматического транспорта. Поскольку холин должен быть удален из внутриклеточной среды при помощи переносчиков, этот этап является лимитирующей стадией в процессе синтеза ацетилхолина.
Высвобождение ацетилхолина. Пузырьки у пресинаптического нервного окончания высвобождают содержимое в синаптическую щель, когда концентрация Са2+ в цитоплазме возрастает в ответ на потенциал действия (ПД) (А). Адреналин и норадреналин могут ингибировать высвобождение ацетилхолина путем стимуляции пресинаптических a2-адренорецепторов. В постганглионарных парасимпатических волокнах ацетилхолин блокирует свое собственное высвобождение при помощи связывания с пресинаптическими ауторецепторами (М-рецепторами; см. ниже), как показано на рис. Б.
Ацетилхолин связывается с постсинаптическими холинергическими рецепторами, или холинорецепторами, в вегетативных ганглиях и органах, иннервируемых парасимпатическими волокнами, например в сердце, в гладкой мускулатуре (глаз, бронхов, мочеточников, мочевого пузыря, гениталий, кровеносных сосудов, пищевода и желудочно-кишечного тракта), в слюнных и слезных железах, а также в симпатически иннервируемых потовых железах. Холинорецепторы бывают никотинергическими (N) или мускаринергическими (М). N-холинорецепторы (никотинергические) стимулируются алкалоидом никотином, а М-холинорецепторы (мускаринергические) - алкалоидом яда грибов мускарином.
NN-холинорецепторы вегетативных ганглиев, специфичные для нервов (А), отличаются от специфичных для мышц NM-холинорецепторов на двигательной концевой пластинке тем, что образованы разными субъединицами. Они сходны тем, что и те и другие являются ионотропными рецепторами, т. е. они действуют одновременно как холинорецепторы и как катионные каналы. Связывание ацетилхолина ведет к быстрому притоку ионов Na+ и Са2+ и к раннему (быстрому) возбуждающему постсинаптическому потенциалу действия (ВПСП), который запускает постсинаптический потенциал действия (ПД), как только превышается пороговый потенциал (А, левая часть).
М-холинорецепторы (М1-М5) воздействуют на синаптическую передачу не прямо, а посредством G-белков (метаботропные рецепторы).
М—холинорецепторы находятся в основном в вегетативных ганглиях (А), в ЦНС, в клетках желез внешней секреции. Они активируют фосфолипазу Сβ (ФЛ-Сβ) с помощью Gq-белков в постганглионарных нейронах, в результате чего высвобождаются инозитолтрифосфат (ИТФ) и диацилглицерол (ДАГ) (вторичные мессенджеры). Это стимулирует приток Са2+ и поздний возбуждающий постсинаптический потенциал (ВПСП) (А, средняя часть рисунка). Передача синаптического сигнала модулируется поздним ВПСП, а также котрансмиттерными пептидами, запускающими пептидергический ВПСП или ингибирующий постсинаптический потенциал СИПСП1 (А, правая часть).
М2-холинорецепторы расположены в сердце и функционируют в основном опосредованно, через Gj-белки. Gj-белок открывает специфические К+-каналы, локализованные в основном в синусном узле, атриовентрикулярном узле и в клетках предсердия, таким образом оказывая отрицательный хронотропный и дромотропный эффекты на сердце (Б). Gj-белки также ингибируют аденилатциклазу, снижая таким образом приток Са2+ (Б).
М3-холинорецепторы расположены в основном в гладких мышцах. Как и М1-холинорецепторы (А, средняя часть рисунка), М3-холинорецепторы запускают сокращение путем стимуляции входа Са2+ в клетку. Однако они также могут индуцировать релаксацию путем активации Са2+-зависимой NO-син-тазы, например в клетках эндотелия.
Конец действия ацетилхолина достигается путем опосредованного ацетилхолинэстеразой расщепления молекулы ацетилхолина в синаптической щели. Примерно 50% высвобождаемого холина вновь поглощается пресинаптическими нервными окончаниями (Б).
Антагонисты. Атропин блокирует все М-холинорецепторы, тогда как пирензепин, метоктрамин, гекса-гидрозиладифенидол и тропикамид селективно блокируют M1-, М2-, М3- и М4-холинорецепторы соответственно, тубокурарин- NM-холинорецепторы, а триметафан- NM-холинорецепторы.
Некоторые нейроны могут образовывать (при участии ферментов) L-допа или L-дофа (L-дигидрооксифенилаланин) из аминокислоты L-тирозина. L-допа - предшественник дофамина, норадреналина и адреналина - трех природных катехоламинов, которые синтезируются в указанной последовательности. Дофамин является финальным продуктом синтеза в нейронах, содержащих только ферменты, необходимые на первой стадии синтеза (это декарбоксилаза ароматических L-аминокислот). Дофамин используется в качестве медиатора дофаминергическими нейронами в ЦНС и вегетативными нейронами, иннервирующими почки.
Норадреналин (НА) синтезируется в том случае, если присутствует также второй фермент (дофамин-β-карбоксилаза). В большинстве симпатических постганглионарных нервных окончаниях и в норадре-нергических центральных нейронах норадреналин служит нейромедиатором наряду с комедиаторами -аденозинтрифосфатом (АТФ), соматостатином и нейропептидом Y (НП-Y).
В мозговом веществе надпочечников (см. ниже) и в адренергических нейронах продолговатого мозга N-метилтрансфераза фенилэтаноламина превращает норадреналин в адреналин (А).
Окончания немиелинизированных симпатических постганглионарных нейронов являются шишковидными (узловатыми), или варикозными (А). Эти узлы представляют собой синаптические контакты, хотя и не всегда близкие, с эффекторными органами. Они также служат участками синтеза и хранения норадреналина. L-Тирозин (А1) активно поглощается нервными окончаниями и превращается в дофамин. При адренергической стимуляции эта стадия ускоряется зависимым от протеинкиназы А (ПК-A; А2) фосфорилированием соответствующего фермента. Это приводит к увеличению запасов дофамина. Дофамин помещается в хромаффинные везикулы, где превращается в норадреналин (АЗ). Норадреналин, будучи конечным продуктом, ингибирует дальнейший синтез дофамина (отрицательная обратная связь).
Высвобождение норадреналина. Норадреналин высвобождается в синаптическую щель после поступления в нервные окончания потенциала действия и инициации притока Са2+ (А4).
Адренергические рецепторы, или адренорецепторы (В). Различают четыре основных типа адренорецепторов (а1, а2, β1 и β2) в зависимости от их аффинности к адреналину и норадреналину, а также к многочисленным агонистам и антагонистам. Все адренорецепторы реагируют на адреналин; однако норадреналин оказывает слабое действие на β2-адренорецепторы. Изопротеренол (изопреналин) активирует только β-адренорецепторы, а фентоламин блокирует только а-адренорецепторы. Активность всех адренорецепторов опосредована G-белками.
Выделяют различные подтипы (a1A, а1B. а1D) a1-адренорецепторов (Б1). Их расположение и функции; ЦНС (усиление симпатической активности), слюнные железы, печень (усиление гликогенолиза), почки (действие на порог высвобождения ренина) и гладкая мускулатура (сигнал к сокращению артериол, матки, различных протоков, бронхиол, мочевого пузыря, желудочно-кишечных сфинктеров и расширителя зрачка).
Активация а1-адренорецепторов (Б1), опосредованная Gq-белками и фосфолипазой Сβ (ФЛ-Сβ), ведет к формированию вторичных мессенджеров: инозитолтрифосфата (ИТФ), который увеличивает внутриклеточную концентрацию Са2+, и диацилглицерола (ДАГ), который активирует протеинкиназу С (ПК-С). Опосредованная Gj-белками активация a1-адренорецепторов также приводит к активации Са2+-зависимых К+-каналов. Результирующая утечка К+ гиперполяризует и расслабляет мишень - гладкую мышцу (например, в желудочно-кишечном тракте).
Различают три подтипа (a2A, а2B, а2C) a2-адренорецепторов (Б2). Их расположение и функции: ЦНС (снижение симпатической активности; например, использование а2-агониста клонидина для снижения кровяного давления), слюнные железы (уменьшение слюноотделения), островковые клетки поджелудочной железы (снижение секреции инсулина), жировые клетки (снижение липолиза), тромбоциты (увеличение агрегации) и нейроны (пресинаптические ауторецепторы, см. ниже). Активированные а2-адренорецепторы (Б2) связываются с G1 белком и ингибируют (посредством аi-субъединицы Gj-белков) аденилатциклазу (синтез цАМФ) и в то же время при помощи βу-субъединицы Gj-белков увеличивают вероятность открывания зависимых от потенциала К+-каналов (гиперполяризация). Так как аг-адренорецепторы сопряжены с Go-белками, они также ингибируют зависимые от потенциала Са2+-каналы (снижая [Са2+]внутр ).
Все β-адренорецепторы сопряжены с GS-белком и их aS-субъединицы высвобождают цАМФ как вторичный мессенджер. Затем цАМФ активирует протеинкиназу А (ПК-A), которая фосфорилирует различные белки, что зависит от типа клеток-мишеней.
Норадреналин и адреналин действуют через β1-адренорецепторы (БЗ) и открывают посредством цАМФ и протеинкиназы А Са2+-каналы L-типа в мембранах клеток сердца. Это увеличивает [Са2+]внутр. и таким образом производит положительные хронотропные, дромотропные и инотропные эффекты. Активированные Gs-белки могут также напрямую увеличивать вероятность открывания зависимых от потенциала Са2+-каналов в сердце (БЗ). В почках базальная секреция ренина увеличивается посредством β1-адренорецепторов.
Активация β2-адренорецепторов адреналином (БД) увеличивает уровень цАМФ, таким образом снижая [Са2+]внутр. (механизм этого процесса пока не ясен). Это расширяет бронхиолы и кровеносные сосуды в скелетных мышцах и расслабляет мышцы матки, различных протоков и желудочно-кишечного тракта. Дальнейшие эффекты активации β2-адренорецепторов - увеличенная секреция инсулина и гликогенолиз в печени и мышцах, а также пониженная агрегация тромбоцитов.
Адреналин также увеличивает высвобождение норадреналина в норадренергических волокнах посредством активации пресинаптических β2-адренорецепторов (Д2, А5).
Продукция тепла увеличивается при помощи β3-адренорецепторов клеток бурого жира.
Норадреналин в синаптической щели инактивируется при помощи (А6а-г):
- диффузии норадреналина из синаптической щели в кровь;
- вненейронного поглощения адреналина (в сердце, железах, гладких мышцах, нейроглии и печени) и последующей внутриклеточной деградации норадреналина посредством катехоламин-O-метилтрансферазы (КОМТ) и моноаминоксидазы (МАО);
- активного вторичного поглощения норадреналина (70%) пресинаптическим нервным окончанием. Некоторое количество поглощенного норадреналина входит во внутриклеточные везикулы (АЗ) и используется повторно, а остальная часть инактивируется при помощи МАО;
- стимуляции пресинаптических а2-адренорецепторов (ауторецепторов А6г, 7) норадреналином синаптической щели, который ингибирует дальнейшее высвобождение норадреналина.
Пресинаптические а2-адренорецепторы также можно обнаружить в холинергических нервных окончаниях, например в желудочно-кишечном тракте (снижение подвижности) и предсердии (отрицательный дромотропный эффект), а пресинаптические М-холинорецепторы присутствуют в норадренергических нервных окончаниях. Их взаимодействие позволяет в некоторой степени регулировать периферическую нервную систему. Мозговое вещество надпочечников
После стимуляции преганглионарных симпатических волокон (холинергическая передача) 95% всех клеток в мозговом веществе секретирует в кровь при помощи экзоцитоза гормон адреналин (А), а другие 5% - высвобождают норадреналин (НА). Синтез норадреналина в мозговом веществе аналогичен таковому в норадренергических нейронах (см. выше), но большая часть норадреналина покидает везикулы и превращается в цитоплазме в адреналин. Специальные везикулы, называемые хромаффинными тельцами, после этого активно запасают адреналин и готовятся к высвобождению его и комедиаторов: знкефалина, нейропептида Y путем экзоцитоза.
При реакциях организма на стресс секреция адреналина (и некоторого количества норадреналина) мозговым веществом существенно увеличивается - ответ на физический, ментальный или эмоциональный стресс. Таким образом, клетки, не иннервированные симпатически, активируются и при стрессовой реакции. Адреналин увеличивает высвобождение норадреналина из нейронов посредством пресинаптических β2-адренорецепторов (А2). Секреция адреналина мозговым веществом (опосредованная увеличением симпатической активности) стимулируется некоторыми триггерами, например физической работой, холодом, теплом, тревогой, гневом (стрессом), болью, дефицитом кислорода или снижением кровяного давления. Например, при тяжелой гипогликемии ( Нехолинергические и неадренергические нейромедиаторы Править
У человека гастринвысвобождающий пептид (ГВП) и вазоактивный интестинальный (кишечный) пептид (ВИП) служат комедиаторами в преганглионарных симпатических волокнах; а нейропептид Y (НП-Y) и соматостатин - в постганглионарных волокнах. Постганглионарные парасимпатические волокна используют в качестве комедиаторов нейропептиды энкефалин, вещество Р и/или НП-Y.
Первичной целью преганглионарной секреции пептидов, по-видимому, является регуляция постсинаптических нейронов. Существует весомое доказательство того, что АТФ (аденозинтрифосфат), НП-Y и ВИП также функционируют как независимые нейромедиаторы в вегетативной нервной системе. ВИП и ацетилхолин часто присутствуют одновременно (но в нескольких разных везикулах) в парасимпатических волокнах кровеносных сосудов, экзокринных железах и потовых железах. В желудочно-кишечном тракте ВИП (вместе с оксидом азота) индуцирует истощение слоя круговых мышц и мышц сфинктера, а также (вместе с комедиаторами динорфином и галанином) увеличивает желудочную секрецию. Oксид азота (N0) высвобождается из нитроергических нейронов.
Предназначение вегетативной нервной системы – контроль и коррекция деятельности внутренних органов. Процесс осуществляется автономно – без участия сознания людей. Это позволяет молниеносно реагировать на изменения во внешней среде, агрессиях извне. Однако, при необходимости люди могут оказывать влияние на вегетативные проявления – опосредованно, к примеру, с помощью медикаментов либо физиотерапевтических процедур.
Что собой представляет вегетативная часть нервной системы
Несмотря на огромное влияние вегетативной системы на организм каждого человека, как биологической единицы, по сути, никто не может сказать, что способен ежесекундно чувствовать ее работу. При правильном функционировании люди просто ощущают себя здоровыми.

В этом и состоит главная цель вегетативного сегмента – создание внутри организма аппарата, который бы соединял все органы и ткани в единый конгломерат для сохранения человека, как цельной природной единицы. К примеру, при повышении температуры внешней среды сразу же корректируется деятельность, дыхательной, сердечнососудистой и обменной системы. Они, взаимодействуя, создают комфортные условия для работы головного мозга и жидких тканей – профилактика обезвоживания.
К тому же вегетативный отдел контролирует пищеварительную, мочевыделительную и репродуктивную функцию. Ни одна внутренняя структура не остается без двойного присмотра – к примеру, одни импульсы замедляют частоту пульса, а иные – учащают сердцебиение. В этом заключается преимущество организма людей перед растительным или же животным миром.
По сути, на протяжении эволюции вегетативные отделы позволили людям приспосабливаться к меняющимся внешним условиям и выживать человеческому роду. В новых обстоятельствах сердечнососудистая и дыхательная система, а также пищеварение обеспечивали внутренние ткани питательными веществами. Это гарантировало сохранность особи. В последующем иннервация усложнялась и видоизменялась. В конечном итоге у современного человека без вегетативной регуляции не происходит ни одного вида деятельности, пусть и на бессознательном уровне.
Структурные особенности системы
В целом, вегетативная нервная регуляция – это сложная комбинация, как по анатомическим, так и функциональным признакам нервных элементов. В первую очередь, специалисты выделяют в ней центральный, а также периферический сегмент. Так, скопления нейронов – особых клеток, образуют своеобразные ядра в толще головного либо спинного мозга. Эти центры несут ответственность за реакцию зрачков, работу пищеварительных и дыхательных отделов.
Особое место отведено гипоталамусу и мозговой лимбической системе, как важным частям вегетативной регуляции. И если первый из них хорошо работает, то у людей железы внутренней и внешней секреции здоровы и вырабатывают биологические вещества в требуемом количестве. Поведенческие реакции также будут здоровыми – эмоции, сновидения, работоспособность.
Тогда как периферическая вегетативная нервная часть – это вегетативные нервы, а также отдельные клетки, либо сплетения. С их помощью регулирующий импульс доходит до требуемой зоны и осуществляется коррекция внутренней среды.

Помимо этого, вегетативная система обязательно рассматривается специалистами как совокупность двух крупных отделов – парасимпатического, а также симпатического. Их различают функциональные обязанности. Так, парасимпатический отдел своими нейромедиаторами – химическими молекулами, регулирует образование слюны, правильность сердечного ритма, параметры давления, моторику петель кишечника.
Тогда как, спинной мозг, где находятся центры симпатической части вегетативного отдела, несет ответственность за противоположные реакции – учащение сердцебиение, частоты дыхания, расслабление желчного пузыря, расширение зрачка. В большинстве случаев автономный отдел преганглионарными волокнами и постганглионарными сплетениями самостоятельно справляется со всеми задачами. Головной мозг далеко не всегда вмешивается в его работу.
Функции системы
Описать все многообразие функций вегетативной системы можно тем, что она регулирует физиологические процессы в тканях и обеспечивает постоянство жизнедеятельности – особь приспосабливается и выживает. Для этого нервные импульсы поступают непосредственно в иннервируемый орган, сосуд либо участок ткани. К примеру, гладкомышечные клетки кишечника.
Регулированию подлежат все метаболические процессы – приспособление к снижению/повышению концентрации гормонов, пищеварительных ферментов. Это адаптационно-трофическая вегетативная функция. В ее основе лежит транспорт питательных веществ, их перемещение внутрь клеток. Одни активизируют метаболизм, другие усиливают трофику тканей.
Функции симпатических волокон:
- изменение сокращения сердечной мышцы, возрастание ритма;
- повышение систолического давления;
- расширение диаметра бронхов, а также зрачков;
- снижение тонуса гладких мышц в кишечнике;
- повышение скорости свертывания крови и активности ферментов.
Функции парасимпатических волокон:
- снижение сердечного ритма;
- уменьшение артериального давления;
- обеспечение бронхоспазма;
- повышение тонуса мышечного слоя стенки кишечника.
При этом не следует рассматривать перечисленные функции систем в отдельности – они тесно взаимодействуют. Без одной из них не будут осуществляться и другие виды вегетативного контроля.
Формирование и развитие системы
После оплодотворения яйцеклетки в женском организме, происходит слияние двух клеток – развивается плод. Формирование непосредственно нервной системы происходит уже на 3–4 недели роста малыша.
Из особых первичных клеток нейробластов постепенно формируются симпатические узлы – для локализации в полостных органах. К примеру, в районе сердца и кишечника. Подобное формирование в период эмбриогенеза заканчивается к началу 8–9 недели.

Парасимпатический сегмент изначально размещается в районе лицевой части будущего головного мозга – из тех же нейробластов. В этот же период происходит закладка вегетативных спинномозговых центров – из симпатобластов.
Высшая вегетативная регуляция начинается с образования головного мозга. Требуемые параметры приобретает лимбическая подсистема и гиппокамп, гипоталамус и кора мозговых полушарий. Дальнейшая дифференциация вегетативных структур осуществляется по мере роста плода.
Поэтому так важно для будущей матери избегать малейших негативных воздействий – приема медикаментов, алкогольной и табачной продукции, токсических растворов. В противном случае высок риск появления различных отклонений в дальнейшем функционировании нервной системы ребенка. При тяжелых вегетативных поражениях дети становятся инвалидами и требуют специализированного наблюдения и лечения.
Отличительные признаки систем
Помимо непосредственно функциональных обязанностей, для сравнительной характеристики соматической и вегетативной нервной системы присуще иное расположение ядер – в головном, а также спином мозге. Они имеют очаговый, прерывистый характер у симпатического, а также парасимпатического отдела, но размещены равномерно в соматическом сегменте.
Иные различия вегетативной и соматической систем:
- иннервация гладкой мускулатуры осуществляется непроизвольно;
- в ряде органов наблюдается мощное сокрушение мышечных групп – к примеру, в сфинктерах;
- соматический отдел контролирует мускулатуру скелетного строения – побуждает ее к быстрым, а также сознательным сокращениям;
- вегетативное влияние обеспечивает трофику;
- очаговый выход вегетативных корешков, как от внутричерепных, так и от спинномозговых ядер – принцип сегментарности постганглионарными симпатическими, а также парасимпатическими периферическими волокнами не соблюдается;
- различие присутствует и в строении рефлекторных дуг, к тому же вся деятельность вегетативного отдела основана не только на высших центральных, но и на периферических дугах.
Специалистами было выяснено, что у вегетативных отделов присутствует ряд примитивных черт – диффузность размещения нейронов, однообразие форм, а также размеров нейронов, меньший калибр волокон из-за отсутствия миелиновой оболочки. Поэтому и скорость иннервации существенно ниже. К тому же вегетативный отдел обладает меньшей избирательностью к гормонам и механизму метаболизма.
Признаки расстройства вегетативных структур
Сложность строения и функционирования как парасимпатической, так и симпатической вегетативной системы обусловливает, что сбой в одном их сегменте, будет негативно сказываться на деятельности всего организма.
Заподозрить появление расстройства в иннервируемом органе можно по ряду признаков. К примеру, при частых симптомах сухости во рту, дрожи в кистях рук либо треморе век. Иногда на вегетативные отклонения в системе указывают проблемы со сном – трудности засыпания, прерывистость ночного отдыха, разбитость в утренние часы.
Характерными будут колебания артериального давления и температуры – без предшествующего развития гипертонической болезни либо инфекционного процесса. Человек ощущает приливы жара и зябкости, головные боли и ухудшение зрения – затем самочувствие улучшается.
В стрессовых ситуациях сбои здоровья различимы четче – резкие расстройства сердечнососудистых и пищеварительных функций, сбои в эндокринных либо дыхательных органах. Симптомы выглядят, как нарастание одышки, позывы на тошноту, рвоту, боли в районе сердца, желудка.
На подобные сигналы организма необходимо обращать пристальное внимание. В противном случае вегетативные расстройства переходят в серьезные заболевания внутренних органов, с последующими осложнениями. Вылечить сбои в парасимпатическом либо симпатическом отделе системы намного легче на начальном этапе их появления. На помощь приходят силы природы – народные рецепты отваров и настоев, современные аптечные средства, санаторно-курортное оздоровление, к примеру, гидротерапия, солнечные ванны, ароматерапия.

Из Википедии, бесплатной энциклопедии
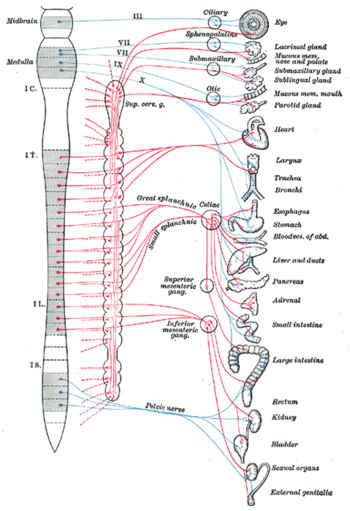
Вегетати́вная не́рвная систе́ма [1] (от лат. vegetatio — возбуждение, от лат. vegetativus — растительный), ВНС, автономная нервная система, ганглионарная нервная система (от лат. ganglion — нервный узел), висцеральная нервная система (от лат. viscera — внутренности), органная нервная система, чревная нервная система, systema nervosum autonomicum (PNA) — часть нервной системы организма, комплекс центральных и периферических клеточных структур, регулирующих функциональный уровень организма, необходимый для адекватной реакции всех его систем.
Вегетативная нервная система — отдел нервной системы, регулирующий деятельность внутренних органов, желез внутренней и внешней секреции, кровеносных и лимфатических сосудов [2] . Играет ведущую роль в поддержании постоянства внутренней среды организма и в приспособительных реакциях всех позвоночных.
Анатомически и функционально вегетативная нервная система подразделяется на симпатическую, парасимпатическую и метасимпатическую. Симпатические и парасимпатические центры находятся под контролем коры больших полушарий и гипоталамических центров [3] .
В симпатическом и парасимпатическом отделах имеются центральная и периферическая части. Центральную часть образуют тела нейронов, лежащих в спинном и головном мозге. Эти скопления нервных клеток получили название вегетативных ядер. Отходящие от ядер волокна, вегетативные ганглии, лежащие за пределами центральной нервной системы, и нервные сплетения в стенках внутренних органов образуют периферическую часть вегетативной нервной системы.
Симпатические ядра расположены в спинном мозге. Отходящие от него нервные волокна заканчиваются за пределами спинного мозга в симпатических узлах, от которых берут начало нервные волокна. Эти волокна подходят ко всем органам.
Парасимпатические ядра лежат в среднем и продолговатом мозге и в крестцовой части спинного мозга. Нервные волокна от ядер продолговатого мозга входят в состав блуждающих нервов. От ядер крестцовой части нервные волокна идут к кишечнику, органам выделения.
Метасимпатическая нервная система представлена нервными сплетениями и мелкими ганглиями в стенках пищеварительного тракта, мочевого пузыря, сердца и некоторых других органов.
Деятельность вегетативной нервной системы не зависит от воли человека. Это означает, что в обычных условиях человек не может волевым усилием заставить сердце биться реже или мышцы желудка — не сокращаться. Однако достичь сознательного влияния на многие параметры, контролируемые ВНС, можно с помощью специальных методов тренировки — например, с использованием методов биологической обратной связи.
Симпатическая нервная система усиливает обмен веществ, повышает возбуждаемость большинства тканей, мобилизует силы организма на активную деятельность. Парасимпатическая система способствует восстановлению израсходованных запасов энергии, регулирует работу организма во время сна.
Под контролем автономной системы находятся органы кровообращения, дыхания, пищеварения, выделения, размножения, а также обмен веществ и рост. Фактически эфферентный отдел ВНС осуществляет нервную регуляцию функций всех органов и тканей, кроме скелетных мышц, которыми управляет соматическая нервная система.
В отличие от соматической нервной системы, двигательный эффекторный нейрон в автономной нервной системе находится на периферии, и спинной мозг лишь косвенно управляет его импульсами.
Терминологическая путаница возникает также, когда автономной называют всю висцеральную систему (и афферентную, и эфферентную).
Классификация отделов висцеральной нервной системы позвоночных, приведённая в руководстве [5] А. Ромера и Т. Парсонса, выглядит следующим образом:
- афферентная;
- эфферентная:
- особая жаберная;
- автономная:
- симпатическая;
- парасимпатическая.
Содержание
- 1 Морфология
- 1.1 Центральный и периферический отделы
- 1.1.1 Центральный отдел
- 1.1.2 Периферический отдел
- 1.2 Симпатический, парасимпатический и метасимпатический отделы
- 1.3 Расположение ганглиев и строение проводящих путей
- 1.4 Рефлекторная дуга
- 1.1 Центральный и периферический отделы
- 2 Физиология
- 2.1 Общее значение вегетативной регуляции
- 2.2 Роль симпатического и парасимпатического отделов
- 2.3 Влияние симпатического и парасимпатического отделов на отдельные органы
- 2.4 Нейромедиаторы и клеточные рецепторы
- 3 Развитие в эмбриогенезе
- 4 Сравнительная анатомия и эволюция вегетативной нервной системы
- 5 См. также
- 6 Примечания
- 7 Литература
- 8 Ссылки
Морфология [ править | править код ]
Выделение автономной (вегетативной) нервной системы обусловлено некоторыми особенностями её строения. К этим особенностям относятся следующие:
- очаговость локализации вегетативных ядер в ЦНС;
- скопление тел эффекторных нейронов в виде узлов (ганглиев) в составе вегетативных сплетений;
- двухнейронность нервного пути от вегетативного ядра в ЦНС к иннервируемому органу.
Волокна автономной нервной системы выходят не сегментарно, как в соматической нервной системе, а из трёх отстоящих друг от друга ограниченных участков мозга: черепного, грудинопоясничного и крестцового.
Автономную нервную систему разделяют на симпатическую, парасимпатическую и метасимпатическую части. В симпатической части отростки спинномозговых нейронов короче, ганглионарные длиннее. В парасимпатической системе, наоборот, отростки спинномозговых клеток длиннее, ганглионарных короче. Симпатические волокна иннервируют все без исключения органы, в то время как область иннервации парасимпатических волокон более ограничена.
Автономная (вегетативная) нервная система подразделяется по топографическому признаку делятся на центральный и периферический отделы.
- парасимпатические ядра 3, 7, 9 и 10 пар черепных нервов, лежащие в мозговом стволе (краниобульбарный отдел).;ядра, лежащие в среднем мозге(мезоенцефалический отдел).; ядра, залегающие в сером веществе трёх крестцовых сегментов (сакральный отдел) [6]
- симпатические ядра, расположенные в боковых рогах тораколюмбального отдела спинного мозга;
- вегетативные (автономные) нервы, ветви и нервные волокна, выходящие из головного и спинного мозга;
- вегетативные (автономные, висцеральные) сплетения;
- узлы (ганглии) вегетативных (автономных, висцеральных) сплетений;
- симпатический ствол (правый и левый) с его узлами (ганглиями), межузловыми и соединительными ветвями и симпатическими нервами;
- концевые узлы (ганглии) парасимпатической части вегетативной нервной системы.
На основании топографии вегетативных ядер и узлов, различий в длине аксонов первого и второго нейронов эфферентного пути, а также особенностей функции вегетативная нервная система подразделяется на симпатическую, парасимпатическую и метасимпатическую.
Нейроны ядер центрального отдела вегетативной нервной системы — первые эфферентные нейроны на пути от ЦНС (спинной и головной мозг) к иннервируемому органу. Нервные волокна, образованные отростками этих нейронов, носят название предузловых (преганглионарных) волокон, так как они идут до узлов периферической части вегетативной нервной системы и заканчиваются синапсами на клетках этих узлов. Преганглионарные волокна имеют миелиновую оболочку, благодаря чему отличаются беловатым цветом. Они выходят из мозга в составе корешков соответствующих черепных нервов и передних корешков спинномозговых нервов.
Вегетативные узлы (ганглии): входят в состав симпатических стволов (есть у большинства позвоночных, кроме круглоротых и хрящевых рыб), крупных вегетативных сплетений брюшной полости и таза, располагаются в области головы и в толще или возле органов пищеварительной и дыхательной систем, а также мочеполового аппарата, которые иннервируются вегетативной нервной системой. Узлы периферической части вегетативной нервной системы содержат тела вторых (эффекторных) нейронов, лежащих на пути к иннервируемым органам. Отростки этих вторых нейронов эфферентного пути, несущих нервный импульс из вегетативных узлов к рабочим органам (гладкая мускулатура, железы, ткани), являются послеузелковыми (постганглионарными) нервными волокнами. Из-за отсутствия миелиновой оболочки они имеют серый цвет. Постганглионарные волокна автономной нервной системы в большинстве своем тонкие (чаще всего их диаметр не превышает 7 мкм) и не имеют миелиновой оболочки. Поэтому возбуждение по ним распространяется медленно, а нервы автономной нервной системы характеризуются бо́льшим рефрактерным периодом и большей хронаксией.
Строение рефлекторных дуг вегетативного отдела отличается от строения рефлекторных дуг соматической части нервной системы. В рефлекторной дуге вегетативной части нервной системы эфферентное звено состоит не из одного нейрона, а из двух, один из которых находится вне ЦНС. В целом простая вегетативная рефлекторная дуга представлена тремя нейронами.
Первое звено рефлекторной дуги — это чувствительный нейрон, тело которого располагается в спинномозговых узлах и в чувствительных узлах черепных нервов. Периферический отросток такого нейрона, имеющий чувствительное окончание — рецептор, берёт начало в органах и тканях. Центральный отросток в составе задних корешков спинномозговых нервов или чувствительных корешков черепных нервов направляется к соответствующим ядрам в спинной или головной мозг.
Второе звено рефлекторной дуги является эфферентным, поскольку несёт импульсы из спинного или головного мозга к рабочему органу. Этот эфферентный путь вегетативной рефлекторной дуги представлен двумя нейронами. Первый из этих нейронов, второй по счёту в простой вегетативной рефлекторной дуге, располагается в вегетативных ядрах ЦНС. Его можно называть вставочным, так как он находится между чувствительным (афферентным) звеном рефлекторной дуги и вторым (эфферентным) нейроном эфферентного пути.
Эффекторный нейрон представляет собой третий нейрон вегетативной рефлекторной дуги. Тела эффекторных (третьих) нейронов лежат в периферических узлах вегетативной нервной системы (симпатический ствол, вегетативные узлы черепных нервов, узлы внеорганных и внутриорганных вегетативных сплетений). Отростки этих нейронов направляются к органам и тканям в составе органных вегетативных или смешанных нервов. Заканчиваются постганглионарные нервные волокна на гладких мышцах, железа́х и в других тканях соответствующими концевыми нервными аппаратами.
Физиология [ править | править код ]
Вегетативная нервная система приспосабливает работу внутренних органов к изменениям окружающей среды. ВНС обеспечивает гомеостаз (постоянство внутренней среды организма). ВНС также участвует во многих поведенческих актах, осуществляемых под управлением головного мозга, влияя не только на физическую, но и на психическую деятельность человека.
Симпатическая нервная система активируется при стрессовых реакциях. Для неё характерно генерализованное влияние, при этом симпатические волокна иннервируют подавляющее большинство органов.
Известно, что парасимпатическая стимуляция одних органов оказывает тормозное действие, а других — возбуждающее действие. В большинстве случаев действие парасимпатической и симпатической систем противоположно.
Влияние симпатического отдела:
- На сердце — повышает частоту и силу сокращений сердца.
- На артерии — [7] сужает артерии большинства органов, расширяет артерии скелетных мышц.
- На кишечник — угнетает перистальтику кишечника и выработку пищеварительных ферментов.
- На слюнные железы — угнетает слюноотделение.
- На мочевой пузырь — расслабляет мочевой пузырь.
- На бронхи и дыхание — расширяет бронхи и бронхиолы, усиливает вентиляцию лёгких.
- На зрачок — расширяет зрачки.
Влияние парасимпатического отдела:
- На сердце — уменьшает частоту и силу сокращений сердца.
- На артерии — не влияет в большинстве органов, вызывает расширение артерий половых органов и мозга, сужение коронарных артерий и артерий лёгких.
- На кишечник — усиливает перистальтику кишечника и стимулирует выработку пищеварительных ферментов.
- На слюнные железы — стимулирует слюноотделение.
- На мочевой пузырь — сокращает мочевой пузырь.
- На бронхи и дыхание — сужает бронхи и бронхиолы, уменьшает вентиляцию лёгких.
- На зрачок — сужает зрачки.
Симпатический и парасимпатический отделы оказывают различное, в ряде случаев противонаправленное влияние на различные органы и ткани, а также перекрёстно влияют друг на друга. Различное воздействие этих отделов на одни и те же клетки связано со спецификой выделяемых ими нейромедиаторов и со спецификой рецепторов, имеющихся на пресинаптических и постсинаптических мембранах нейронов автономной системы и их клеток-мишеней.
Преганглионарные нейроны обоих отделов автономной системы в качестве основного нейромедиатора выделяют ацетилхолин, который действует на никотиновые рецепторы ацетилхолина на постсинаптической мембране постганглионарных (эффекторных) нейронов. Постганглионарные нейроны симпатического отдела, как правило, выделяют в качестве медиатора норадреналин, который действует на адренорецепторы клеток-мишеней. На клетках-мишенях симпатических нейронов бета-1 и альфа-1 адренорецепторы в основном сосредоточены на постсинаптических мембранах (это означает, что in vivo на них действует в основном норадреналин), а аль-2 и бета-2 рецепторы — на внесинаптических участках мембраны (на них в основном действует адреналин крови). Лишь некоторые постганглионарные нейроны симпатического отдела (например, действующие на потовые железы) выделяют ацетилхолин.
Постганглионарные нейроны парасимпатического отдела выделяют ацетилхолин, который действует на мускариновые рецепторы клеток-мишеней.
На пресинаптической мембране постганглионарных нейронов симпатического отдела преобладают два типа адренорецепторов: альфа-2 и бета-2 адренорецепторы. Кроме того, на мебране этих нейронов расположены рецепторы к пуриновым и пиримидиновым нуклеотидоам (P2X-рецепторы АТФ и др.), никотиновые и мускариновые холинорецепторы, рецепторы нейропептидов и простагландинов, опиоидные рецепторы [8] .
При действии на альфа-2 адренорецепторы норадреналина или адреналина крови падает внутриклеточная концентрация ионов Ca 2+ , и выделение норадреналина в синапсах блокируется. Возникает петля отрицательной обратной связи. Альфа-2 рецепторы более чувствительны к норадреналину, чем к адреналину.
При действии норадреналина и адреналина на бета-2 адренорецепторы выделение норадреналина обычно усиливается. Этот эффект наблюдается при обычном взаимодействии с Gs-белком, при котором растёт внутриклеточная концентрация цАМФ. Бета-два рецепторы более чувствительны к адреналину. Поскольку под действием норадреналина симпатических нервов из мозгового слоя надпочечников выделяется адреналин, возникает петля положительной обратной связи.
Однако в некоторых случаях активация бета-2 рецепторов может блокировать выделение норадреналина. Показано, что это может быть следствием взаимодействия бета-2 рецепторов с Gi/o белками и связывания (секвестирования) ими Gs-белков, которое, в свою очередь, предотвращает взаимодействие Gs-белков с другими рецепторами [1].
При действии ацетилхолина на мускариновые рецепторы симпатических нейронов выделение норадреналина в их синапсах блокируется, а при действии на никотиновые рецепторы — стимулируется. Поскольку на пресинаптических мембранах симпатических нейронов преобладают мускариновые рецепторы, обычно активация парасимпатических нервов снижает уровень выделения норадреналина из симпатических нервов.
На пресинаптических мембранах постганглионарных нейронов парасимпатического отдела преобладают альфа-2 адренорецепторы. При действии на них норадреналина выделение ацетилхолина блокируется. Таким образом, симпатические и парасимпатические нервы взаимно ингибируют друг друга.
Развитие в эмбриогенезе [ править | править код ]
- Развитие периферической (соматической) и вегетативной нервной системы. Периферическая (соматическая) и вегетативная нервная система развивается из наружного зародышевого листка — эктодермы. Черепные и спинномозговые нервы у плода закладываются очень рано (5-6 нед). Миелинизация нервных волокон происходит позже (у преддверного нерва — 4 мес; у большинства нервов — на 6-7-м месяце).
Спинномозговые и периферические вегетативные узлы закладываются одновременно с развитием спинного мозга. Исходным материалом для них служат клеточные элементы ганглиозной пластинки, её нейробласты и глиобласты, из которых образуются клеточные элементы спинномозговых узлов. Часть их смещается на периферию в места локализации вегетативных нервных узлов
Сравнительная анатомия и эволюция вегетативной нервной системы [ править | править код ]
У насекомых имеется так называемая симпатическая, или стомодеальная нервная система [9] . В её состав входит фронтальный ганглий, который находится спереди от головного мозга и соединён парными коннективами с тритоцеребрумом. От него отходит непарный фронтальный нерв, тянущийся вдоль спинной стороны глотки и пищевода. Этот нерв соединяется с несколькими нервными ганглиями; отходящие от них нервы иннервируют переднюю кишку, слюнные железы и аорту.
Читайте также:
