
Виды тонуса в цнс
Тонус (греч. tonos напряжение) — постоянная (фоновая) активность нервных центров, некоторых тканей и органов, обеспечивающая их готовность к действию. Тонус является одним из проявлений гомеостаза в организме и одновременно одним из механизмов его поддержания (см. Гомеостаз). Так, постоянство Тонуса гладких мышц стенок артерий является одним из условий поддержания АД на гомеостатическом уровне.
На практике обычно выделяют Тонус нервных центров, Тонус скелетных и Тонус гладких мышц.
Тонус нервных центров
Тонус нервных центров (нервный тонус) обусловлен притоком импульсов с соответствующих рецептивных полей (см. Рецепторные зоны) и восходящими активирующими влияниями ретикулярной формации (см.). Нейронам ретикулярной формации свойственна стойкая импульсная фоновая активность, связанная с высокой хим. чувствительностью мембраны ее клеток и конвергенцией к пей коллатералей от огромного количества сенсорных путей. Поэтому, как правило, влияние ретикулярной формации на другие структуры носит тонический характер. В поддержании Т. нервных центров, помимо афферентной импульсации, поступающей от периферических рецепторов, а также от ретикулярной формации, участвуют различные гуморальные раздражители (гормоны, уровень парциального напряжения углекислоты в крови и т. д.). Постоянная фоновая активность нервных центров делает возможным существование тонических рефлексов (см. Рефлекс) и поддержание в целостном организме состояния бодрствования.
Мышечный тонус
Для определения Т. мышц применяют различные методы — прямые, основанные на определении пассивного сопротивления мышцы ее предельному растяжению, и косвенные, когда величину тонуса оценивают по сопутствующим изменениям механических характеристик мышечной ткани. К прямым методам можно отнести методы Моссо (1896) и И. Н. Филимонова (1925), применение к-рых позволяет регистрировать не только кривую сопротивления мышцы при пассивном движении, но и отдельные этапы движения во времени. В методе Бромана (1949) сочетается изучение сопротивления пассивному растяжению мышцы и электромиографическая регистрация процессов, протекающих при этом. К косвенным методам определения мышечного Т. относится измерение поперечной твердости (резистентности) мышц по глубине погружения (вдавливания) в мышцу груза.
К мышечным проприоцепторам (см.) относятся мышечные веретена и сухожильные органы. Мышечные рецепторы — это специализированные клетки, к-рые реагируют на изменения длины и напряжения мышц, происходящие при их растяжении или сокращении. Веретена соединены с мышечными волокнами параллельно, а сухожильные органы — последовательно. Поэтому сокращение мышцы сопровождается возбуждением сухожильных органов и снижением импульсации в рецепторах веретен. Каждое мышечное веретено состоит из нескольких мышечных волокон и окружающей их соединительнотканной капсулы, к-рая покрывает веретено не полностью; полюса наиболее длинных волокон выходят за ее пределы. Мышечные волокна (см. Мышечная ткань), входящие в состав веретена, получили название интрафузальных (внутриверетенных), а основные волокна скелетной мышцы — экстрафузальных (вневеретеппых). Спирали и кольца, образующие первичное нервное окончание, расположены экваториально, и каждая спираль получает ветвь от толстого афферентного волокна; каждому веретену соответствует одно афферентное волокно. Вторичные окончания расположены ближе к полюсам веретена и иннервируются более тонкими нервными волокнами (см.).
Эфферентная иннервация веретен осуществляется тонкими аксонами мотонейронов, относящихся к группе А гамма-нервных волокон (гамма-эфферентная иннервация). Экстрафузальные волокна иннервируются толстыми А альфа-нервными волокнами. Небольшая часть веретен у млекопитающих получает иннервацию от ветвей альфа-нервного волокна. Но интрафузальные волокна снабжаются не только альфа-нервными волокнами: на интрафузальных волокнах того же веретена оканчиваются и гамма-нервные волокна. Все эфферентные волокна, поступающие в веретено, называются фузимоторными.
При сокращении интрафузальных волокон растягивается экваториальная область веретена, в результате чего происходит смещение витков спирали рецептора, и как следствие — возникновение ритмической биоэлектрической активности. Изолированное раздражение гамма-волокон на фоне блокады проведения возбуждения по альфа-эфферентным волокнам не сопровождается приростом мышечного напряжения, но приводит к усилению импульсации в афферентных волокнах.
Импульсация гамма-мотонейронов усиливает активность мышечного веретена. Раздражение первичных окончаний активирует альфа-мотонейроны мышцы и ее синергистов. При этом активность альфа-мотонейронов мышц-антагонистов тормозится. Так как первичные окончания могут быть растянуты в результате активности фузимоторных нервов, то возникло предположение о возможности возбуждения альфа-мотонейронов через гамма-петлю.
Наряду с активацией α-мотонейронов через систему мышечных веретен возможно и их прямое возбуждение. В мышцах-разгибателях возбуждение вторичных окончаний вызывает торможение активности α-мотонейронов. При осуществлении миотатических рефлексов (т. е. рефлексов на растяжение) раздражению, вызывающему возбуждение первичных окончаний, противодействуют активация сухожильных органов, вторичных рецепторов и разгрузка веретен (см. Рефлекс).
Система тонических мелких альфа-мотонейронов чувствительна к импульсам от веретен, приходящим по толстым афферентным волокнам, и почти не создает активного напряжения мышц, способного ослабить эффект этих импульсов.
Совместная активация альфа- и гамма-мотонейронов, характерная для многих движений, позволяет контролировать длину мышечных волокон; незначительное увеличение нагрузки приводит к удлинению мышцы, к-рому противодействует афферентная импульсация от одновременно активированных веретен. Создаются условия для постоянной коррекции изменений активной позы.
Т. о., тонус скелетных мышц поддерживается системой тонических альфа-мотонейронов, импульсация к-рых зависит от уровня активности первичных окончаний мышечных веретен. Первичные окончания могут быть растянуты при возбуждении гамма-мотонейронов. Следовательно, Т. скелетных мышц опосредуется через гамма-петлю. Чередование сокращений различных моторных единиц в пределах одной мышцы в процессе поддержания Т. препятствует развитию утомления мышцы в целом и обеспечивает ее постоянную активность, а также сохранение позы без значительных энергозатрат.
Тонус гладких мышц обеспечивается двумя механизмами: миогенным и нейрогуморальным. Тонические сокращения гладких мышц могут продолжаться длительное время на низком энергетическом уровне, отличаясь тем самым от фазных сокращений скелетных мышц, что объясняется неодинаковой структурой миофибрилл различных видов мышечной ткани.
Особенностью нек-рых гладкомышечных клеток стенок сосудов является их способность к спонтанной активности и распространению возбуждения от клетки к клетке. Эта особенность гладких мышц обеспечивает базальный Т. сосудов, поддерживающий их в состоянии нек-рого напряжения при отсутствии внешних (нервных или гуморальных) влияний. Воздействия, уменьшающие уровень мембранного потенциала (напр., механическое растяжение или набухание клеток), увеличивают частоту спонтанных разрядов и амплитуду сокращения гладких мышц. Наоборот, гиперполяризация клеточных мембран приводит к исчезновению спонтанного возбуждения и мышечных сокращений. Метаболиты, вырабатываемые тканями, оказывают активное воздействие на гладкомышечные клетки по принципу отрицательной обратной связи. Так, при повышении Т. прекапиллярных сфинктеров уменьшается капиллярный кровоток и увеличивается концентрация метаболитов, что оказывает сосудорасширяющее действие (см. Микроциркуляция). Сосудорасширяющее действие оказывает низкое напряжение кислорода и высокое — углекислоты, повышение концентрации водородных ионов, молочной к-ты и др.
Активность гладких мышц артерий и вен, а также наружных слоев крупных прекапиллярных сосудов, как и скелетных, регулируется в основном внешними рефлекторными и гуморальными влияниями. При этом возрастает роль центральных нервных и гуморальных механизмов регуляции сосудистого Т. Возбуждение сихмпатической нервной системы способствует сокращению наружных слоев сосудистой стенки, и в ответную реакцию вовлекаются внутренние слои за счет распространения возбуждения от клетки к клетке (этим достигается централизация контроля над сосудами). При отсутствии симпатических влияний и расслаблении наружных слоев сосудистой стенки контроль над местным кровотоком осуществляется гладкими мышцами ее средних слоев, к-рые обладают собственным базальным Т. В связи с тем, что вены играют роль емкостных сосудов, чувствительность их мышечных клеток к растяжению низка. В гладких мышцах вен (за исключением воротной и брыжеечных вен) не обнаружено признаков спонтанной активности, и в обеспечении их Т. большая роль принадлежит рефлекторным механизмам регуляции. Хотя уровень базального Т. сосудистой системы определяется взаимодействием местных механических и хим. факторов, рефлекторные механизмы осуществляют контроль над всей ситуацией в целом, что создает условия для перераспределения кровотока в соответствии с потребностями организма. Напр., симпатические адренергические нервы оказывают сосудосуживающее действие под влиянием изменения состояния нервных центров (напр., гипоталамуса) или при рефлекторных реакциях, осуществляющихся на бульбарном и спинальном уровнях в результате изменения активности рецепторного аппарата сосудистых рефлексогенных зон. Гуморальное влияние на сосудистый Т. оказывает целый ряд веществ (гормоны коркового и мозгового вещества надпочечников, ангиотензин II, вазопрессин, простагландины, гистамин, серотонин и т. д.).
Наличие двух механизмов (миогенного и нейрогуморального), обусловливающих Т. сосудов, создает условия для поддержания определенного уровня системного АД, а также для быстрого местного перераспределения кровотока в связи с изменяющимися потребностями организма.
Патология мышечного тонуса может проявляться либо в виде его снижения (атония, мышечная гипотония), либо в виде повышения (мышечная гипертония). Для объективной регистрации мышечного тонуса в клинике используется пальпация мышц или оценка степени сопротивления мышцы сжатию с помощью специальных приборов. Они позволяют регистрировать также сопротивление мышцы при ее пассивном растяжении. В этом случае определяют состояние так наз. контрактильного Т., под к-рым понимают рефлекторное напряжение мышц, вызываемое при ее растяжении вследствие раздражения проприоцепторов (см.). Вместе с тем с помощью этих приборов часто бывает невозможно или трудно определять синергические изменения мышечного Т. в различных мышечных группах. Эти методы более пригодны для оценки различных терапевтических воздействий, направленных на нормализацию мышечного Т. Согласно данным Гепферта (H. Gopfert, 1960), идеальная возможность количественного определения мышечного тонуса была бы достигнута в том случае, если можно было бы установить число возбудимых в данное время мышечных волокон. Ближе всего подходит к указанной цели электромиографическая регистрация активности мышечных волокон (см. Электромиография). Регистрация электрической активности мышц при различных позотонических реакциях оказалась возможной с помощью высокочувствительных электрографических аппаратов, позволяющих улавливать характерные для тонических напряжений низковольтные колебания биопотенциала. Электромиографические исследования Т. производятся в норме и патологии при пассивных движениях и путем регистрации биоэлектрической активности в состоянии мышечного покоя, при различных рефлекторных тонических реакциях, а также при произвольных движениях.
В клип, практике чаще исследуют сопротивление мышц растяжению. При этом обследуемого просят расслабить мышцы, не оказывать активного сопротивления и производят пассивное движение, определяя при этом степень тонического сопротивления мышц.
Нередко мышечная гипотония возникает и при отсутствии непосредственного перерыва спинальной рефлекторной дуги (напр., при поражении мозжечка и его связей). Мышечная гипотония может возникать при остро возникающих церебральных параличах, напр, в раннем постинсультном периоде, и связана с временным или более стойким угнетением функции эфферентного отдела спинальной дуги (клеток переднего рога) вследствие диасхиза (см.), ослабления облегчающих супраспинальных влияний. В большинстве случаев эта диасхизальная гипотония сменяется мышечной гипертонией, являющейся следствием стойкого поражения депрессорных (тормозных) систем головного мозга и постепенным усилением облегчающих нисходящих влияний на спинальные мотонейроны.
Иногда мышечная гипотония при постинсультных параличах остается стойкой (см. Параличи, парезы). Это наблюдается, в частности, при поражении нижнестволовых и мозжечковых систем, а также при плохом общем соматическом состоянии больных со вторичным изменением функции Т. нервных центров.
Повышение мышечного тонуса связывают с поражением пирамидной и экстрапирамидной системы, причем основную роль в генезе мышечной гипертонии играет поражение экстрапирамидной системы (см.).
Нарушения мышечного Тонуса наблюдаются при многих заболеваниях нервной системы. Мышечная гипотония и атония характерна для заболеваний, связанных с поражением переднероговых мотонейронов и периферической нервной системы (см. Полиомиелит, Полиневрит, Плексит, Невриты); наблюдается она и при распространенном поражении задних корешков и задних столбов спинного мозга, напр, при спинной сухотке (см.), при заболеваниях головного мозга (напр., при поражении мозжечка и его связей, стриарной системы). Мышечная спастичность сопутствует многим церебральным и спинальным заболеваниям, протекающим с поражением нервных центров и двигательных путей, передающих депрессорные влияния на переднероговые мотонейроны. Мышечная ригидность может развиваться при многих очаговых поражениях головного мозга, но наиболее характерна для заболеваний, связанных с поражением экстрапирамидной системы (см. Паркинсонизм, Гепато-церебральная дистрофия). В раннем детском возрасте изменения мышечного Т. могут отмечаться при церебральных детских параличах (см.); при нек-рых врожденных заболеваниях, напр, при миатонии Оппенгейма (см. Миатония). Нередко элементы пластического тонуса наблюдаются вследствие возрастных изменений подкорковых образований головного мозга.
Библиогр.: Вейн А. М., Голубев В. и Берзиньш Ю. Э. Паркинсонизм, Рига, 1981; Гранит Р. Основы регуляции движений, пер. с англ., М., 1973; Давиденков С. Н. Проблема силы, Tonus’а и содружественных движений при церебральных параличах, Изв. Бакинск. Гос. ун-та, № 1, с. 49, 1921; Джонсон П. Периферическое кровообращение, пер. с англ., М., 1982; 3авалишин И. А. и Новикова В. П. Сравнительный анализ некоторых функциональных приемов электро-миографического исследования при спастичности разного происхождения, Журн. невропат, и психиат., т. 71, в. 12, с. 1812, 1971; Лунев Д. К. Нарушения мышечного тонуса при мозговом инсульте, М., 1974; Физиология движений, под ред. В. С. Гурфинкеля и др., Л., 1976; Фолков Б. и Нил Э. Кровообращение, пер. с англ., М., 1976; Юсевич Ю. С. Электромиография тонуса скелетной мускулатуры человека в норме и патологии, М., 1963, библиогр.; Brоman Т. Electromyo-mechanographic registrations of passive movements in normal and pathological subjects, Acta psychiat. scand., suppl. 53, p. 1, 1949; Handbook of clinical neurology, ed. by P. J. Vinken a. G. W. Bruyn, v. 1 — 2, Amsterdam a. o., 1975; Rushworth G. Spasticity and rigidity, on experimental study and review, J. Neurol. Neurosurg. Psychiat., v. 23, p. 99, 1960.
Г. И. Косицкий, М. И. Монгуш; Д. К. Лунев (невр.).
Скелетные мышцы всегда находятся в состоянии некоторого напряжения. Постоянное незначительное напряжение мышц, не сопровождающееся признаками утомления, называется мышечным тонусом. Односторонняя перерезка у спинальной лягушки, подвешенной на крючке штатива, чувствительных (задних) корешков спинного мозга, в которых проходят афферентные нервные волокна, иннервирующие соответствующую заднюю лапку, приводит к исчезновению мышечного тонуса этой лапки и она распрямляется. К аналогичному эффекту приводит перерезка передних (двигательных) корешков или разрушение спинного мозга. Эти опыты свидетельствуют о том, что при разрушении основных звеньев рефлекторного кольца (афферентных и эфферентных путей, нервных центров) мышечный тонус исчезает. Следовательно, мышечный тонус имеет рефлекторную природу.
Источником возбуждений, поддерживающих мышечный тонус, являются проприорецепторы. В скелетных мышцах имеются три вида проприорецепторов:
• мышечные веретена, расположенные среди мышечных волокон;
• сухожильные рецепторы Гольджи, расположенные в сухожилиях;
• пачиниевы тельца, расположенные в фасциях, сухожилиях, связках.
Особое значение в регуляции мышечного тонуса имеют мышечные веретена и сухожильные рецепторы Гольджи.
Мышечные веретена представляют собой небольшие продолговатые образования, напоминающие своим внешним видом прядильные капсулы мышечного веретена находится пучок мышечных волокон, которые называются интрафузальными, т. к. они расположенными внутри веретена в отличие от обычных мышечных волокон, которые называются зкстрафузальными.
Каждое интрафузальное волокно состоит из трех частей:
• его центральная часть называется ядерной сумкой, в которой находятся ядра мышечной клетки;
• два периферических участка, которые имеют поперечную исчерченность и обладают способностью сокращаться;
• миотрубки, расположенные между ядерной сумкой и периферическими участками.
Ядерную сумку в виде спирали окружают нервные волокна чувствительного нейрона-первичные рецепторные окончания. В области миотрубок нервные окончания афферентных нейронов гроздевидно ветвятся, образуя вторичные рецепторные окончания.
В мышце мышечное веретено одним концом прикрепляется к экстрафузальному мышечному волокну, а другим - к сухожилию этого волокна. Таким образом, мышечное веретено расположено в мышце параллельно экстрафузальным мышечным волокнам (рис.19).
При снижении тонуса экстрафузального волокна увеличивается его длина, что приводит к растяжению и раздражению первичных и вторичных рецепторных окончаний, для которых растяжение является адекватным раздражителем.
Возбуждение от рецепторных окончаний по афферентным волокнам поступает в спинной мозг к мотонейронам, расположенным в передних рогах. Мотонейроны спинного мозга принято подразделять на альфа- и гамма-мотонейроны (так как их аксоны относятся к А-альфа и А-гамма нервным волокнам). Возбуждение от альфа-мотонейронов поступает к экстрафузальным мышечным волокнам, вызывая их сокращение - тонус восстанавливается. Избыточное сокращение экстрафузальных мышечных волокон приводит к растяжению сухожильных рецепторов Гольджи, так как они прикрепляются к мышце последовательно. В них возникает возбуждение, которое поступает к тормозным вставочным нейронам спинного мозга, а от них к альфа-мотонейронам. Активность альфа-мотонейронов при этом снижается, уменьшается импульсация, идущая от них к экстрафузальным мышечным волокнам, тонус несколько снижается.
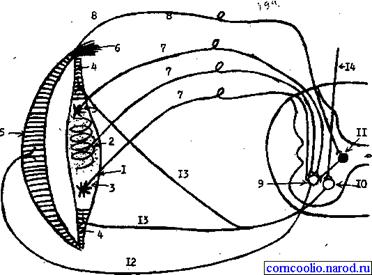
Рис. 19. Морфологические структуры, обеспечивающие регуляцию мышечного тонуса на спинальном уровне (схема). 1 - интрафузальное волокно, 2 - ядерная сумка с первичными рецепторными окончаниями; 3 - миотрубки с вторичными рецепторными окончаниями; 4 - периферические участки интрафузального волокна; 5 - экстрафузальное мышечное волокно; 6 - сухожильный рецептор Гольджи; 7 - афферентные волокна от первичных и вторичных рецепторных окончаний; 8 - афферентные волокна от сухожильного рецептора Гольджи; 9 - альфа-мотонейрон спинного мозга; 10 - гамма-мотонейрон спинного мозга; 11 - промежуточный (тормозной) нейрон спинного мозга; 12 - эфферентный путь от альфа-мотонейрона к экстрафузальному мышечному волокну, 13 - эфферентный путь от гамма-мотонейрона к периферическим участкам интрафузального волокна; 14 - возбуждение, приходящее из ЦНС к гамма-мотонейронам.
Высокая возбудимость мышечного веретена поддерживается за счет специального механизма, который образован сократительными элементами, расположенными в периферических участках интрафузальных волокон по обе стороны от ядерной сумки. Сокращение этих участков вызывает растяжение ядерной сумки и миотрубок, что приводит к возбуждению рецепторных окончаний и увеличению потока афферентных возбуждений к альфа-мотонейронам. Степень сокращения сократительных элементов интрафузальных волокон регулируется гамма-мотонейронами спинного мозга. Импульсы, приходящие по гамма-афферентным волокнам, вызывают сокращение периферических участков интрафузальных волокон.
Рассмотренные выше механизмы поддержания мышечного тонуса осуществляются на уровне спинного мозга, поэтому такой тонус называется спинальным или простейшим. Спинальный тонус характеризуется очень слабой выраженностью тонического напряжения. Такой тонус не может обеспечить поддержание позы животного и акт ходьбы, но он достаточен для осуществления простейших спинальных рефлексов.
Перерезка у животного (например, у кошки) ствола мозга между передними и задними буграми четверохолмия (операция перерезки ствола мозга называется децеребрацией) вызывает особое состояние скелетной мускулатуры, которое называется децеребрационной ригидностью или контрактильным тонусом. Это состояние характеризуется резким повышением тонуса разгибательной мускулатуры. Конечности такого животного сильно вытянуты, голова запрокинута, спина выгнута. Это состояние называется опистотонусом. Необходимо приложить большое усилие, чтобы согнуть у такого животного конечность в суставе.
Контрактильный тонус имеет рефлекторную природу. Это доказывается тем, что при перерезке передних или задних корешков спинного мозга, иннервирующих конечность, ригидность мускулатуры этой конечности исчезает.
Важную роль в возникновении контрактильного тонуса играет дорсальное вестибулярное ядро продолговатого мозга (ядро Дейтерса), которое возбуждается импульсами от рецепторов вестибулярного аппарата. При раздражении вестибулярного ядра ригидность усиливается, а при его разрушении ригидность уменьшается. Полагают, что это ядро оказывает влияние на ретикулярную формацию продолговатого мозга, которая оказывает неодинаковое влияние на нейроны спинного мозга. Раздражение медиальных отделов ретикулярной формации приводит к торможению рефлексов спинного мозга (тормозящая ретикуло-спинальная система), а раздражение латеральных отделов вызывает активацию нейронов спинного мозга (облегчающая ретикуло-спинальная система).
Ядро Дейтерса продолговатого мозга оказывает активирующее влияние на облегчающую ретикуло-спинальную систему и угнетает тормозную ретикуло-спинальную систему. Активность вестибулярного ядра тормозится красным ядром среднего мозга. При децеребрации происходит разобщение красного и вестибулярного ядер, в результате чего снижается тормозящее влияние красного ядра на вестибулярное ядро. В этих условиях оно оказывает сильное активирующее влияние на облегчающую ретикуло-спинальную систему. Поток импульсов от нее поступает в спинной мозг к гамма-мотонейронам, а от них к сократительным участкам интрафузальных волокон. Сокращение этих участков вызывает раздражение проприорецепторов. Поток возбуждений от них поступает к альфа-мотонейронам спинного мозга, а от них - к экстрафузальным мышечным волокнам, вызывая резкое увеличение их тонуса.
Разрушение красного ядра у интактного животного приводит к развитию гипертонуса мышц-разгибателей, а при его раздражении - к снижению тонуса.
Обнаружены и прямые связи красного ядра и ядра Дейтерса с мотонейронами спинного мозга. Показано, что ядро Дейтерса тормозит мотонейроны мышц-сгибателей и возбуждает мотонейроны мышц-разгибателей. Красное ядро оказывает тормозное влияние на мышцы-разгибатели и активирующее влияние на мышцы-сгибатели. Следовательно, при децеребрации, когда красное ядро отделено от ядра Дейтерса, создаются все условия для повышения тонуса мышц-разгибателей.
На ядро Дейтерса тормозное влияние оказывает и мозжечок. Удаление червячной зоны мозжечка у животного, находящегося в состоянии децеребрационной ригидности, вызывает большее растормаживание ядра Дейтерса и дальнейшее увеличение тонуса разгибателей. Электрическое раздражение червячной зоны, напротив, приводит к уменьшению тонуса этих мышц за счет активации тормозных влияний мозжечка на ядро Дейтерса.
При перерезке головного мозга выше промежуточного мозга у животного возникает особое изменение тонуса - мышцы становятся пластичными (воскоподобными), при этом конечностям можно легко придать любое положение, которое они могут сохранять длительное время. Такое состояние называется пластическим тонусом или восковой ригидностью. Пластический тонус имеет рефлекторное происхождение: после перерезки чувствительных нервов, иннервирующих конечность, все проявления пластического тонуса на этой конечности исчезают.
В возникновении пластического тонуса определенную роль играет черная субстанция среднего мозга. Черная субстанция функционально связана с базальными ганглиями - бледным шаром и полосатым телом. Нейроны черной субстанции синтезируют медиатор дофамин. Аксоны этих нейронов подходят к полосатому телу, которое также содержит дофамин. Повреждение черной субстанции, вызывающее дегенерацию дофаминергических путей к полосатому телу, сопровождаетеся заболеванием - болезнью Паркинсона. Одним из симптомов этой болезни является восковидная ригидность, которая обусловлена, по-видимому, гиперактивностью базальных ганглиев, возникающей при повреждении дофаминергического (вероятно, тормозного) пути, идущего от черной субстанции к полосатому телу.
Кроме того, черная субстанция, по-видимому, оказывает на скелетные мышцы трофическое влияние, подобно симпатической нервной системе. При перерезке выше промежуточного мозга высвобождаются структуры, которые оказывают на черную субстанцию тормозящее влияние. В таких условиях черная субстанция затормаживается и перестает оказывать достаточное трофическое влияние на скелетные мышцы и они становятся пластичными.
Пластический тонус может возникнуть не только при перерезке мозга, но и, например, при отравлении некоторыми ядами, при заболевании нервной системы, а также под влиянием гипноза. У человека пластический тонус может проявляться при особом состоянии нервной системы, которое называется каталепсией или восковой ригидностью. Человек в таком состоянии на некоторое время как бы цепенеет, причем, в неестественной позе и не меняет ее в течение долгого времени.
Большое значение в регуляции мышечного тонуса имеют базальные ядра - бледный шар и полосатое тело, которые образуют стриопаллидарную систему. Эти структуры регулируют активность всех нижележащих отделов ЦНС, участвующих в регуляции мышечного тонуса, обеспечивая адекватное перераспределение тонуса мышц при различных видах деятельности. При поражении экстрапирамидной системы, составной частью которой являются базальные ядра, возникают нарушения регуляций тонуса мускулатуры, что приводит к развитию так называемых дрожательных параличей (паркинсонизму, атетозу, хорее и др.).
Главную роль в приспособительной регуляции мышечного тонуса выполняет кора головного мозга. С ее участием и участием других структур, регулирующих мышечный тонус, формируется нормальный или корковый тонус.
Регуляция тонуса скелетной мускулатуры осуществляется экстрапирамидной системой головного мозга. Быстрые (фазные) движения обеспечиваются активностью пирамидной системы.
В условиях целого организма пирамидная и экстрапирамидная системы включаются как единое целое в функциональную архитектуру приспособительных актов, обеспечивая необходимые фазные движения и адекватные изменения мышечного тонуса. Специфическое изменение тонуса мышц возникает при осуществлении статических и стато-кинетических рефлексов, направленных на поддержание позы.
Статические рефлексы делятся на рефлексы положения и рефлексы выпрямления. Рефлексы положения обеспечивают формирование тонуса мышц, необходимого для поддержания естественного положения тела в пространстве в состоянии покоя. Рефлексы выпрямления определяют перераспределение тонуса мышц, приводящее к восстановлению естественной для данного вида животного позы в случае ее изменения.
Стато-кинетические рефлексы направлены на сохранение позы в пространстве при ускорениях прямолинейного и вращательного характера. Эти рефлексы проявляются при вращении, перемещении тела в горизонтальной и вертикальной плоскостях. Они возникают, в основном, в результате возбуждения рецепторов вестибулярного аппарата. Так, при вращательном движении наблюдается нистагм головы: вначале голова медленно поворачивается в сторону, противоположную направлению вращения, а затем быстро возвращается в. исходное положение. Такое же движения глазных яблок при вращении называется нистагмом глаз.
Перераспределение тонуса мышц шеи, туловища и конечностей происходит при быстром подъеме и спуске. Быстрый подъем сопровождается повышением тонуса сгибателей, а быстрый спуск - разгибателей конечностей. Эти рефлекторные реакции легко наблюдать при перемещении в скоростном лифте, поэтому они и называются лифтными рефлексами.
Центральная нервная система (ЦНС) – основная часть нервной системы человека. Она состоит из двух отделов: головного мозга и спинного мозга. Основные функции нервной системы –контролировать все жизненно важные процессы в организме. Головной мозг отвечает за мышление, речь, координацию. Он обеспечивает работу всех органов чувств, начиная от простой температурной чувствительности и заканчивая зрением и слухом. Спинной мозг регулирует работу внутренних органов, обеспечивает координацию их деятельности и приводит тело в движение (под контролем головного мозга). Принимая во внимание множество функций ЦНС, клинические симптомы, позволяющие заподозрить опухоль головного или спинного мозга, могут быть чрезвычайно разнообразными: от нарушения поведенческих функций до невозможности осуществлять произвольные движения частями тела, нарушений функции тазовых органов.
Клетки головного и спинного мозга
Головной и спинной мозг состоят из клеток, чьи названия и характеристики определяются их функциями. Клетки, характерные только для нервной системы, – это нейроны и нейроглия.

К опухолям головного мозга, возникающим из нейронов или их предшественников, относятся эмбриональные опухоли (ранее их называли примитивные нейроэктодермальные опухоли - ПНЭО), такие как медуллобластомы и пинеобластомы.
Опухоли, возникающие из нейроглиальных (глиальных) клеток, в общем случае называют глиомами. Однако в зависимости от конкретного типа глиальных клеток, вовлеченных в опухоль, она может иметь то или иное специфическое название. Самые распространeнные глиальные опухоли у детей – мозжечковые и полушарные астроцитомы, глиомы ствола мозга, глиомы зрительныйх путей, эпендимомы и ганглиоглиомы. Виды опухолей подробнее описаны в этой статье.
Строение головного мозга
Головной мозг имеет очень сложное строение. Различают несколько больших его отделов: большие полушария; ствол головного мозга: средний мозг, мост, продолговатый мозг; мозжечок.

Рисунок 2. Строение головного мозга
Если посмотреть на головной мозг сверху и сбоку, то мы увидим правое и левое полушария, между которыми располагается разделяющая их большая борозда — межполушарная, или продольная щель. В глубине мозга находится мозолистое тело – пучок нервных волокон, соединяющий две половины мозга и позволяющих передавать информацию от одного полушария к другому и обратно. Поверхность полушарий изрезана более или менее глубоко проникающими щелями и бороздами, между которыми расположены извилины.
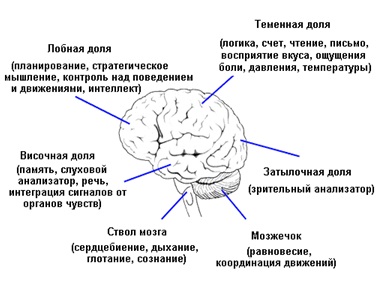
Рисунок 3. Строение полушария головного мозга
Несколько больших углублений (борозд) делят каждое полушарие на четыре доли:
- лобную (фронтальную);
- височную;
- теменную (париетальную);
- затылочную.
Теменные доли ответственны за чувство осязания, восприятие давления, боли, тепла и холода, а также за вычислительные и речевые навыки, ориентацию тела в пространстве. В передней части теменной доли располагается так называемая сенсорная (чувствительная) зона, куда сходится информация о влиянии окружающего мира на наше тело от болевых, температурных и других рецепторов.
Височные доли в значительной мере отвечает за память, слух и способность воспринимать устную или письменную информацию. В них также есть и дополнительные сложные объекты. Так, миндалевидные тела (миндалины) играют важную роль в возникновении таких состояний, как волнение, агрессия, страх или гнев. В свою очередь, миндалины связаны с гиппокампом, который содействует формированию воспоминаний из пережитых событий.
Затылочные доли – зрительный центр мозга, анализирующий информацию, которая поступает от глаз. Левая затылочная доля получает информацию из правого поля зрения, а правая – из левого. Хотя все доли больших полушарий отвечают за определенные функции, они не действуют в одиночку, и ни один процесс не связан только с одной определенной долей. Благодаря огромной сети взаимосвязей в головном мозге всегда существует коммуникация между разными полушариями и долями, а также между подкорковыми структурами. Мозг функционирует как единое целое.
Мозжечок – структура меньшего размера, которая располагается в нижней задней части мозга, под большими полушариями, и отделен от них отростком твердой мозговой оболочки – так называемым наметом мозжечка или палаткой мозжечка (тенториумом). По размеру он приблизительно в восемь раз меньше переднего мозга. Мозжечок непрерывно и автоматически осуществляет тонкое регулирование координации движений и равновесия тела.
Ствол мозга отходит вниз от центра головного мозга и проходит перед мозжечком, после чего сливается с верхней частью спинного мозга. Ствол мозга отвечает за базовые функции организма, многие из которых осуществляются автоматически, вне нашего сознательного контроля, такие как сердцебиение и дыхание. В ствол входят следующие части:
- Продолговатый мозг, который управляет дыханием, глотанием, артериальным давлением и частотой сердечных сокращений.
- Варолиев мост (или просто мост), который соединяет мозжечок с большим мозгом.
- Средний мозг, который участвует в осуществлении функций зрения и слуха.
Вдоль всего ствола мозга проходит ретикулярная формация (или ретикулярная субстанция) – структура, которая отвечает за пробуждение от сна и за реакции возбуждения, а также играет важную роль в регуляции мышечного тонуса, дыхания и сердечных сокращений.
Промежуточный мозг располагается над средним мозгом. В его состав входят, в частности, таламус и гипоталамус. Гипоталамус – это регуляторный центр, участвующий во многих важных функциях организма: в регуляции секреции гормонов (включая гормоны расположенного поблизости гипофиза), в работе автономной нервной системы, процессах пищеварения и сна, а также в контроле температуры тела, эмоций, сексуальности и т.п. Над гипоталамусом расположен таламус, который обрабатывает значительную часть информации, поступающей к головного мозгу и идущей от него.
12 пар черепно-мозговых нервов в медицинской практике нумеруются римскими цифрами от I до XII, при этом в каждой из этих пар один нерв отвечает левой стороне тела, а другой – правой. ЧМН отходит от ствола мозга. Они контролируют такие важные функции, как глотание, движения мышц лица, плеч и шеи, а также ощущения (зрение, вкус, слух). Главные нервы, передающие информацию к остальным частям тела, проходят через ствол мозга.
Мозговые оболочки питают, защищают головной и спинной мозг. Располагаются тремя слоями друг под другом: сразу под черепом находится твердая оболочка (dura mater), имеющая наибольшее количество болевых рецепторов в организме (в мозге их нет), под ней паутинная (arachnoidea), и ниже – ближайшая к мозгу сосудистая, или мягкая оболочка (pia mater).
Спинномозговая (или цереброспинальная) жидкость – это прозрачная водянистая жидкость, которая формирует еще один защитный слой вокруг головного и спинного мозга, смягчая удары и сотрясения, питая мозг и выводя ненужные продукты его жизнедеятельности. В обычной ситуации ликвор важен и полезен, но он может играть и вредную для организма роль, если опухоль головного мозга блокирует отток ликвора из желудочка или если ликвор вырабатывается в избыточном количестве. Тогда жидкость скапливается в головном мозге. Такое состояние называют гидроцефалией, или водянкой головного мозга. Поскольку внутри черепной коробки свободного места для лишней жидкости практически нет, возникает повышенное внутричерепное давление (ВЧД).
У ребёнка могут возникнуть головные боли, рвота, нарушения координации движений, сонливость. Нередко именно эти симптомы и становятся первыми наблюдаемыми признаками опухоли головного мозга.
Строение спинного мозга
Спинной мозг – это фактически продолжение головного мозга, окруженное теми же оболочками и спинномозговой жидкостью. Он составляет две трети ЦНС и является своего рода проводящей системой для нервных импульсов.

Рисунок 4. Строение позвонка и расположение спинного мозга в нем
Спинной мозг составляет две трети ЦНС и является своего рода проводящей системой для нервных импульсов. Сенсорная информация (ощущения от прикосновения, температура, давление, боль) идет через него к головному мозгу, а двигательные команды (моторная функция) и рефлексы проходят от головного мозга через спинной ко всем частям тела. Гибкий, состоящий из костей позвоночный столб защищает спинной мозг от внешних воздействий. Кости, составляющие позвоночник, называют позвонками; их выступающие части можно прощупать вдоль спины и задней части шеи. Различные части позвоночника называют отделами (уровнями), всего их пять: шейный (С), грудной (Th), поясничный (L), крестцовый (S) и копчиковый [1] .
[1] Отделы позвоночника обозначаются латинскими символами по начальным буквам соответствующих латинских названий.
Внутри каждого отдела позвонки пронумерованы.

Опухоль спинного мозга может образоваться в любом отделе –например, говорят, что опухоль обнаружена на уровне С1-С3 или на уровне L5. Вдоль всего позвоночного столба от спинного мозга отходят спинномозговые нервы в количестве 31 пары. Они связаны со спинным мозгом через нервные корешки и проходят через отверстия в позвонках к различным частям тела.
При опухолях спинного мозга возникают нарушения двух видов. Локальные (очаговые) симптомы – боль, слабость или расстройства чувствительности – связаны с ростом опухоли в конкретной области, когда этот рост затрагивает кость и/или корешки спинномозговых нервов. Более общие нарушения связаны с нарушением передачи нервных импульсов через затронутую опухолью часть спинного мозга. Может возникнуть слабость, потеря чувствительности или управления мышцами в той области тела, которая управляется спинным мозгом ниже уровня опухоли (паралич или парез). Возможны нарушения мочеиспускания и дефекации (опорожнения кишечника).
Во время операции по удалению опухоли хирургу иногда приходится удалять фрагмент внешней костной ткани (пластинку дуги позвонка, или дужку), чтобы добраться до опухоли.
Это может впоследствии спровоцировать искривление позвоночника, поэтому такой ребенок должен наблюдаться у ортопеда.
Локализация опухоли в ЦНС
Первичная опухоль головного мозга (то есть та, которая изначально родилась в данном месте и не является метастазом опухоли, возникшей в другом месте тела человека) может быть либо доброкачественной, либо злокачественной. Доброкачественная опухоль не прорастает в соседние органы и ткани, а растет, как бы отодвигая, смещая их. Злокачественное новообразование быстро растет, прорастая в соседние ткани и органы, и часто дает метастазы, распространяясь по организму. Первичные опухоли головного мозга, диагностируемые у взрослых, как правило, не распространяются за пределы ЦНС.
Дело в том, что доброкачественная опухоль, развивающаяся в другой части тела, может расти годами, не вызывая нарушения функции и не представляя угрозы для жизни и здоровья пациента. Рост же доброкачественной опухоли в полости черепа или спинномозговом канале, где мало места, быстро вызывает смещение структур мозга и появление угрожающих жизни симптомов. Удаление доброкачественной опухоли ЦНС также сопряжено с большим риском и не всегда возможно в полном объеме, учитывая количество и характер структур мозга, прилежащих к ней.
Первичные опухоли делят на низко- и высокозлокачественные. Для первых, как и для доброкачественных, характерен медленный рост и, в целом, благоприятный прогноз. Но иногда они могут перерождаться в агрессивный (высокозлокачественный) рак. Подробнее о видах опухолей мозга в статье.
Читайте также:
